
Melatonin and Kidney Health: From Fetal Stage to Later Life


1
Department of Pediatrics, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
2
Department of Pediatrics, Kaohsiung Municipal Feng Shan Hospital—Under the Management of Chang Gung Medical Foundation, Kaohsiung 830, Taiwan
3
Department of Pharmacy, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
4
School of Pharmacy, Kaohsiung Medical University, Kaohsiung 807, Taiwan
5
Institute for Translational Research in Biomedicine, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
6
College of Medicine, Chang Gung University, Taoyuan 333, Taiwan
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2023, 24(9), 8105; https://doi.org/10.3390/ijms24098105
Submission received: 13 April 2023
/
Revised: 27 April 2023
/
Accepted: 27 April 2023
/
Published: 30 April 2023
(This article belongs to the Special Issue Pleiotropic Benefits of Melatonin: From Basic Mechanisms to Disease 2.0)
Abstract
:Melatonin, an endogenous hormone mainly released at night by the pineal gland, has multifaceted biofunctions. Emerging evidence points to melatonin having a crucial role in kidney health and disease. As the prevalence of chronic kidney disease (CKD) is still rising, a superior strategy to advance global kidney health is needed to not just treat CKD, but prevent it early on. Adult kidney disease can have its origins in early life. This review aims to evaluate the recent literature regarding melatonin’s effect on kidney development, its clinical uses in the early stage of life, animal models documenting preventive applications of melatonin on offspring’s kidney-related disease, and a thorough summary of therapeutic considerations concerning melatonin supplementation.
1. Introduction
Melatonin is a pleiotropic hormone mainly secreted at night by the pineal gland, which regulates the circadian rhythm [1]. In addition to its chronobiotic action, melatonin has antioxidant, anti-inflammatory, anti-carcinogenic, anti-apoptotic, anti-hypertensive and immunoregulatory properties [2,3,4,5]. Melatonin has a crucial role in human health and disease across the life span. Melatonin appears to be involved in normal pregnancy and fetal development [6,7]. The physiological effects of melatonin are various, and include multi-organ targets [2,3,4,5]. As a result, accumulating evidence supports the idea that melatonin holds promise in the treatment of various diseases, not only in adults but also in children and neonates [8,9,10,11].
Chronic kidney disease (CKD) is the main cause of death and disease worldwide [12]; it currently affects about 10% of the world’s population. Adult kidney disease can originate in early life, known as “Developmental Origin of Health and Disease” (DOHaD) [13]. Recent advances in human and animal studies have offered ample evidence that adverse environmental stimuli during kidney development increases the risk of CKD in adulthood via renal programming [14,15]. On the other hand, kidney disease can be averted in the early stage of life by reprogramming [16]. This vision suggests that the utmost attention is obligatory for global kidney health strategy, principally emphasizing the prevention of CKD at the earliest stage of life, not simply the treatment of established kidney disease [17].
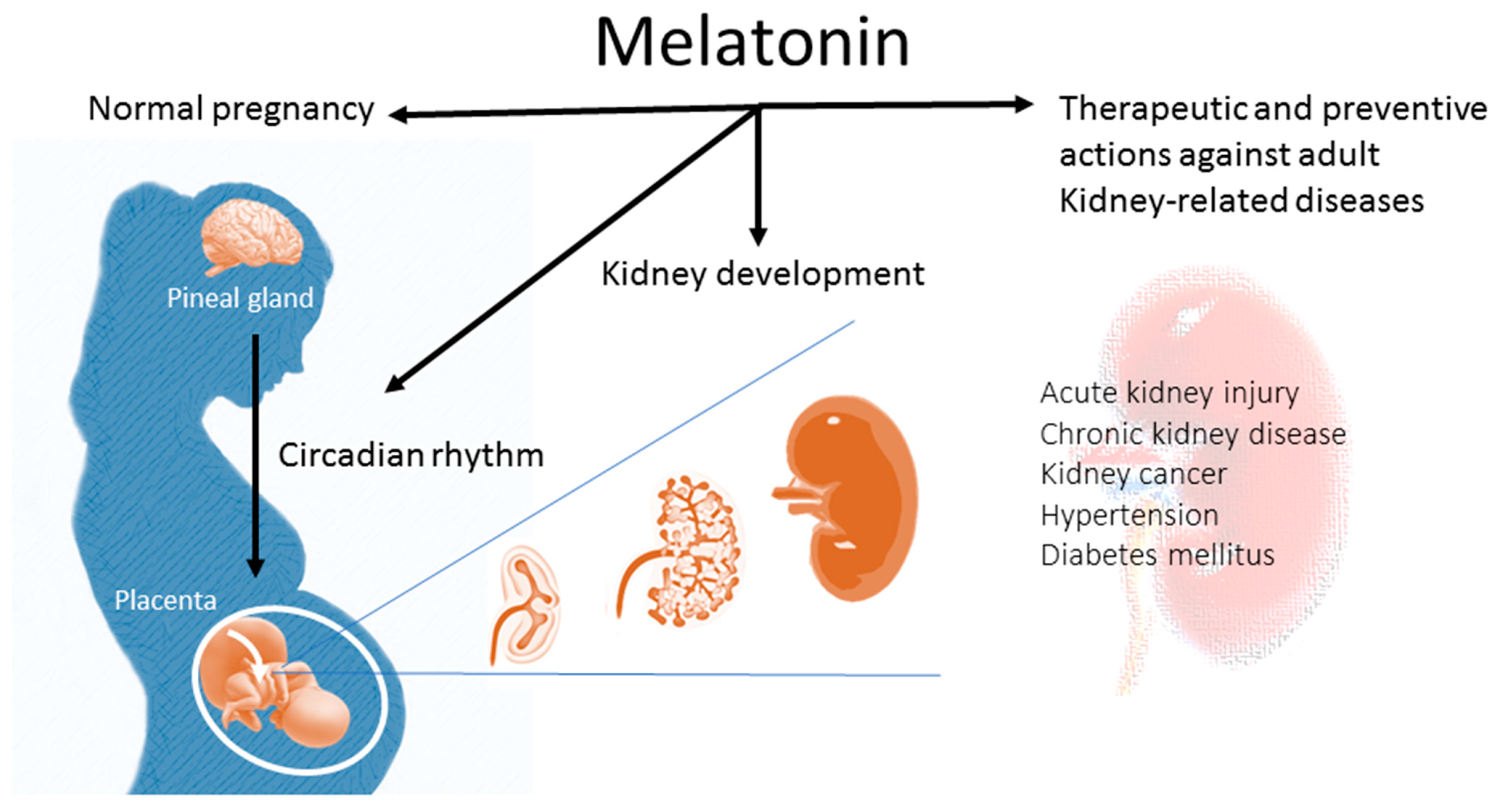
Although multiple positive actions of melatonin have been described, there is little known about the influence of melatonin on kidney health. This review, therefore, highlights the impact of melatonin on kidney development and opens important perspectives for the use of melatonin in preventive and therapeutic applications in kidney-related diseases later in life (Figure 1).
A literature review was carried out by searching the databases Embase, MEDLINE, and Cochrane Library using keywords relevant to melatonin, circadian rhythm, pregnancy, lactation, kidney disease, developmental programming, and DOHaD. A specific focus was put on the use of melatonin during pregnancy, infant, and childhood stages. The reference lists of articles were also examined to identify any additional references that would be related to this review.
2. Effects of Melatonin
2.1. Synthesis, Metabolism, and Action of Melatonin
Melatonin, or 5 methoxy-N-acetyltryptamine, was isolated from the bovine pineal gland and discovered nearly 60 years ago [18]. Melatonin is largely synthesized by the pinealocytes from tryptophan through hydroxylation, decarboxylation, acetylation, and methylation [19]. Melatonin synthesis and secretion is enhanced by darkness and inhibited by light [19]. Once secreted from the pineal gland, melatonin is quickly released into the systemic circulation to reach peripheral target tissues. Other than the pineal gland, many organs can produce melatonin, including the gastrointestinal tract, skin, retina, and bone marrow [20,21,22,23]. Take skin, for example; intracutaneous melatonin metabolites can form a potent anti-oxidative cascade through rapid local metabolism in an MT receptor-independent manner [23]. Therefore, endogenous melatonin production in skin may represent an anti-oxidative system to neutralize pathological changes such as skin aging and cancerogenesis [23].
The half-life of melatonin has been calculated to be around 30–60 min [24]. In the circulation, 70% of melatonin is bound to albumin, while the remaining 30% diffuses to neighboring tissues [4]. Melatonin is mostly metabolized in the liver and kidneys by P450 monooxygenases, and its main urinary metabolite is 6-sulfatoxymelatonin [25]. Only a small percentage (<5%) of blood melatonin is unmetabolized and excreted into the urine.
Melatonin acts through melatonin receptor-1 (MT1) and -2 (MT2), which are G protein-coupled receptors [4]. Melatonin receptors are widely distributed in the body, including the kidney [26]. In the kidneys, MT1 and MT2 receptors are predominately expressed in the kidney membrane area and basolateral membranes [26]. Both melatonin receptors have been shown to activate several signaling pathways, such as the ERK1/2 and the PI3K/AKT pathways [27,28]. Although melatonin regulates circadian rhythms via MT1 and MT2 receptors, the underlying mechanisms are not yet entirely understood and may differ between various tissues [29].
Additionally, the nuclear receptor retinoid acid receptor (ROR) has been suggested to mediate the genomic actions of melatonin [4]. However, recent work revealed that melatonin indirectly rather than directly regulates ROR activity [30]. Additionally, crystallographic evidence does not support the view that ROR is a nuclear receptor of melatonin [31]. Thus, whether ROR is a nuclear receptor of melatonin remains controversial [30]. Moreover, melatonin has receptor-independent effects. For example, melatonin-derived metabolites N1-acetyl-5-methoxykynuramine and N1-acetyl-N2-formyl-5-methoxykynuramine can act as powerful antioxidants [25]. Some melatonin actions could be secondary to its rapid metabolism to different metabolites [2,20].
While the physiologic functions of melatonin have been documented in multiple organ systems, its effects on the kidney are less well recognized. The peripheral circadian clock within the kidneys participates in various physiological functions, including glomerular filtration, tubulo-glomerular feedback mechanisms, the urine concentrating mechanism, circadian blood pressure (BP) rhythm, and the regulation of sodium transport [32]. Renal function is known to vary diurnally in healthy individuals. A previous study demonstrated that the daytime administration of melatonin to hamsters decreased urinary sodium and potassium concentrations as well as urine osmolality [33]. Since the rhythms of melatonin provide synchronization signals for peripheral clocks, dysregulated circadian rhythm and melatonin signaling may be related to kidney-related diseases [34]. These findings suggest that melatonin possibly impacts renal function by regulating proximal tubular function. Additional research is required to clarify the biochemical and physiologic details of melatonin in the kidneys so as to harness its therapeutic potential for kidney diseases.
2.2. Melatonin in Gestation and Fetal Development
Pregnant women have higher nighttime blood melatonin concentrations than non-pregnant women throughout gestation, reaching the highest level at term and falling to the basal level postpartum [35]. As shown in Figure 1, maternal melatonin is able to transfer across the placenta to the fetus, providing photoperiodic information to the fetus [36,37]. In addition to the pineal gland, the placenta can also secrete melatonin [38]. The difference is that the placenta does not produce melatonin in a circadian fashion, and it acts as a paracrine, autocrine, and endocrine hormone [38]. In the placenta, villous trophoblasts do not merely produce melatonin, but express melatonin receptors as well [38]. Melatonin in the placenta is able to work together with the MT1 and MT2 receptors to scavenge free radicals, consequently reducing oxidative damage in compromised pregnancies [39].
Melatonin receptors are widespread in the fetus from the early stages. In rodents, melatonin-binding sites in the pituitary gland are present in 15-day-old fetuses [40]. In the fetal human brain, melatonin receptors exist in many areas [41]. These data suggest that maternal melatonin has a role in the early stages of fetal development. Prior work revealed that a disrupted maternal and embryonic molecular clock impaired organogenesis in the fetus [42]. Another study showed that maternal melatonin deficiency caused circadian rhythm disruption and intrauterine growth retardation (IUGR) in adult rat offspring, which was prevented by maternal melatonin treatment [43]. The findings above support the fact that maternal melatonin is closely linked to fetal development as well as organogenesis.
3. Impact of Melatonin on Kidney Development
3.1. Kidney Development
Human kidney development begins as early as week three, followed by metanephros at five weeks of gestation [44]. Nephrogenesis is initiated when reciprocal interactions between the ureteric bud (UB) and the metanephric mesenchyme (MM) form the UB-derived collecting system and MM-derived nephron. The nephron is the basic structural and functional unit of the kidney [45]. It is composed of the glomerulus and the renal tubule. In humans, the first nephron is formed at nine weeks of gestation. A rapid increase in nephrons occurs from 18 to 32 weeks, and nephrogenesis is complete by 36 weeks of gestation. As nephrogenesis is not completed by preterm birth, nephron under-endowment is prone to be present in premature infants.
Apart from reduced nephron number, impaired nephrogenesis might cause a broad spectrum of malformed kidneys, namely congenital anomalies of the kidney and urinary tract (CAKUT) [46]. It is known that CAKUT is the major cause of pediatric CKD [46]. A low nephron number induces glomerular hyperfiltration and compensatory glomerular hypertrophy, thereby resulting in a vicious cycle of further nephron loss [45].
3.2. Effects of Maternal Melatonin on Offspring Kidney
Maternal melatonin deficiency induces offspring hypertension, a common comorbidity of CKD [49]. There are several ways to induce maternal chronodisruption, including constant light exposure, diurnal light deficiency, continuous darkness, and photoperiod shifts [50]. One study showed that photoperiod shifts during pregnancy adversely program not only BP but also kidney function in adult rat offspring [51]. Another report demonstrated that the exposure of the mother rats to continuous light during pregnancy and lactation caused offspring hypertension [50]. Continuous light exposure alters several kidney genes responsible for high BP [52].
Using whole-genome RNA next-generation sequencing (NGS), we earlier analyzed the renal transcriptome from male rat offspring born to dams that received melatonin supplementation [53]. Melatonin (0.01% in drinking water) was administered during pregnancy and lactation to cover the entire period of nephrogenesis. At 1, 12, and 16 weeks of age, 455, 230, and 132 differentially expressed genes were identified in offspring’s kidneys, respectively. It looks like alterations of transcriptome are induced by melatonin declining over time.
In support of melatonin’s epigenetic actions [54], maternal melatonin therapy can up-regulate several epigenetic regulators during kidney development [53]. Additionally, numerous biological pathways are regulated by melatonin administration during nephrogenesis [53]. Noteworthily, the tryptophan metabolism pathway is regulated by maternal melatonin supplementation in 1-week-old offspring’s kidneys as melatonin is derived from tryptophan. Several genes involved in the melatonin biosynthesis (i.e., Tph1, Ddc, and Asmt) and MT receptors (i.e., Mtnr1b, Rora, and Rorb) were up-regulated, suggesting that melatonin administration to mother rats can program metabolic and signaling pathways of melatonin in their offspring’s kidneys [53]. The findings above indicate that maternal melatonin administration is able to program the offspring’s kidney via the regulation of specific genes and pathways.
4. Preventive and Therapeutic Benefits of Melatonin
Through its pleiotropic effects, melatonin may be efficacious in managing various kidney-related diseases, such as hypertension [55], diabetes mellitus [56], acute kidney injury [57], CKD [58], and kidney cancer [59]. Taking CKD as an example, the renoprotective mechanisms of melatonin cover antioxidant, anti-apoptotic, anti-fibrotic, and anti-inflammatory effects [58]. Although plenty of preclinical studies have been conducted in this regard, evidence is scarce in the clinical setting. In view of the recent advances in DOHaD research, it has become evident that adulthood kidney-related diseases can be averted in the earliest stage by reprogramming [16]. Here, as the scope of the current review, we primarily focused on the use of melatonin from fetal to childhood stages as a therapeutic strategy for kidney disease. For more in-depth information regarding adulthood kidney disease, please refer to reviews published elsewhere.
4.1. Melatonin Use in Humans
Melatonin is a commonly used dietary supplement in the United States [60], while it is a prescription-only drug in Canada, Japan, the European Union, and Australia. Oral melatonin supplementation in humans has a favorable safety profile, with the usual daily doses of melatonin being 2 to 10 mg in diverse populations [61].
However, little information is available regarding melatonin’s use and safety in pregnant or breastfeeding women on the basis of clinical trials. At present, pregnant and lactating women are not recommended to use melatonin considering a lack of human studies [61].
As reviewed elsewhere [10,11], the use of melatonin as a treatment option has been evaluated in neonatal diseases, including hypoxic–ischemic injury, periventricular leukomalacia, respiratory distress syndrome, bronchopulmonary dysplasia, and sepsis. In infants and children, melatonin has also been widely used for purposes including sleep disorders and seizure disorders, and also as an alternative sedative drug [10,11]. Nevertheless, no information exists with regard to kidney disease within the pediatric population. Considering melatonin treatment gives useful results in adult kidney-related diseases, whether melatonin is an effective therapy in pediatric kidney disease is worthy of further evaluation.
4.2. Melatonin Use for the Early Origins of Kidney Diseases in Animal Models
So far, there have been several animal studies reporting that melatonin exogenous supplementation during gestation could be beneficial for both mother and fetus [62], while only a small proportion of them have focused on offspring outcomes [55]. Here, we list in Table 1 a summary of reports relevant to reprogramming effects of melatonin on kidney-related diseases in offspring [63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78]. The therapeutic duration is restricted solely to fetal to early childhood stages prior to disease onset.
Although melatonin has been examined for developmental programming in several species [79,80], the most studied species in this regard are rats. As revealed in Table 1, reprogramming effects of melatonin treatment on rat offspring have been evaluated from aged 8 to 27 weeks. As one rat month is the same as three human years [81], reported renal effects can be translated to human stages from adolescents to young adults. As a result, there remains a lack of information regarding the long-term effects of melatonin on older adults.
Table 1 illustrates various maternal insults that can induce the developmental programming of kidney-related disease in adult offspring, which can be averted by the early use of melatonin. These early life insults include gestational chronodisruption [63], maternal continuous light exposure [65], maternal caloric restriction [66], maternal NG-nitro-L-arginine-methyl ester (L-NAME) exposure [67], maternal high methyl-donor diet [68], maternal high-fructose diet [69], maternal high-fructose diet plus post-weaning high-salt diet [70], antenatal glucocorticoid exposure [71], and antenatal glucocorticoid administration plus post-weaning high-fat diet [72].
Maternal melatonin supplementation could be administered during gestation [73,74], lactation [74,75], or both periods [65,66,67,68,69,70,71,72,73]. Additionally, melatonin treatment during early childhood has been tested in the adenine-primed pediatric chronic kidney disease (CKD) model [76], and the young spontaneously hypertensive rat (SHR) model [77,78]. Table 1 shows that the renal effects of the early use of melatonin included improvement in hypertension, reduced nephron number, kidney function, and altered renal transcriptome.
Notably, certain mechanisms participate in the developmental programming of kidney-related diseases, such as deficient NO, oxidative stress, aberrant RAS, epigenetic regulation, disrupted autophagy–nutrient sensing pathway, glucocorticoid effect, and dysbiotic gut microbiota. Figure 2 is a graphic illustration of the therapeutic and protective mechanisms of melatonin interrelated to renal programming. Below, we will discuss each mechanism in turn.
4.3. Deficient NO
NO has an essential role in renal physiology and BP control [82]. Deficient NO has a crucial role in kidney disease and hypertension of developmental origins [83]. Conversely, adverse programing processes and renal outcomes can be averted by NO-targeting interventions during gestation and lactation. Melatonin can increase NO via reducing asymmetric dimethylarginine (ADMA), an inhibitor of NO synthase [84]. In young SHRs, the antihypertensive effect of melatonin coincided with the decrease of ADMA in the plasma and kidneys [77,78]. In a maternal NO deficiency model, deficient NO in gestation caused by L-NAME administration led to offspring hypertension in adulthood [67], whilst elevated BP and reduced renal NO could be restored concurrently by maternal melatonin therapy [67].
The perinatal use of melatonin is also beneficial to offspring hypertension relevant to the rebalancing of the ADMA-NO pathway in animal models of maternal caloric restriction [66], maternal high-fructose diet [69], and combined maternal high-fructose and post-weaning high-salt diets [70]. In an adenine-induced pediatric CKD model [76], melatonin therapy from 3 to 6 weeks of age prevented CKD-primed hypertension and kidney damage, which was associated with a reduction in ADMA. These data above revealed that melatonin could interact with NO, by which it provided beneficial effects against offspring kidney-related diseases.
4.4. Oxidative Stress
As a well-known antioxidant, melatonin displays a protective role against oxidative stress by scavenging free-radicals and activating antioxidant enzymes [2]. A previous review revealed that several environmental stimuli in pregnancy linked oxidative stress to the developmental programming of kidney disease [85], covering maternal nutrition imbalance, maternal disorders, environmental chemical and toxin exposure, and medication use.
Due to the low-antioxidant capacity of the fetus, a surplus of reactive oxygen species (ROS) under adverse intrauterine conditions overwhelms antioxidants, leading to oxidative damage and, thus, compromising fetal development [86].
Prior work provides evidence of how maternal melatonin therapy protected adult progeny against oxidative stress-related renal programming in models of maternal caloric restriction [66], maternal L-NAME exposure [67], maternal high methyl-donor diet [68], maternal high-fructose diet [69], and antenatal glucocorticoid exposure [71]. When targeting oxidative stress, the antioxidant actions of melatonin included decreased ROS-producing enzyme expression, reduced ROS production, increased antioxidant capacity, and decreased oxidative DNA damage.
Although melatonin has a significant impact on improving oxidative stress, one study revealed that maternal melatonin supplementation which averted the rise in BP in young SHR offspring might not be attributed to its antioxidant effects in kidneys [64]. Even though recent advances have been made in the understanding of how oxidative stress impacts renal programming, further work is needed to discover other protective mechanisms of melatonin, not just its antioxidant functions.
4.5. Disrupted Autophagy–Nutrient Sensing Pathways
Emerging evidence points to the dysregulation of nutrient sensing signaling and autophagy linking to a range of kidney diseases [87,88]. Several nutrient-sensing signals are involved in fetal programming [89], including silent information regulator T1 (SIRT1), peroxisome proliferator-activated receptors (PPARs), AMP-activated protein kinase (AMPK) and PPARγ co-activator 1α (PGC-1α). In pregnancy, maternal nutritional status influences fetal development via nutrient-sensing signals [89]. Melatonin can modulate autophagy by changing nutrient sensing pathways [90].
Melatonin supplementation protecting against kidney disease is connected to AMPK activation [91]. In a maternal methyl-donor diet model, offspring hypertension coincided with the reduced renal expression of SIRT1, AMPKα2, PPARβ, and PPARγ [68]. Another study explored perinatal melatonin therapy and found that its beneficial actions against high-fructose plus high-salt diet-induced offspring hypertension were related to regulating renal AMPKα2, AMPKβ2, SIRT1, SIRT4, PPARγ, and PGC-1α expression [70].
In an antenatal dexamethasone administration plus post-weaning high-fat diet model, the activation of genes related to nutrient sensing and autophagy prevented offspring’s hypertension [92]. These findings were consistent with a previous study showing that melatonin could mediate the renoprotective effect by upregulating the AMPK/SIRT1 axis and enhancing the autophagy in a rat model of diabetic nephropathy [93]. Although a link between melatonin and autophagy behind renal programming has been established, whether its reprogramming effect is attributed to the enhancement of autophagy needs to be evaluated further.
4.6. Aberrant RAS
Melatonin has a role in suppressing RAS [94], which is known as a hormonal cascade controlling BP and kidney development [95,96]. The angiotensin II (Ang II)/angiotensin-converting enzyme (ACE) cascade is known to be the classic RAS, which can be counter-regulated by the Ang (1–7)/ACE2 non-classic axis. In the developing kidney, RAS genes are vastly expressed and have a transient biphasic response with downregulation of the classic RAS in neonates that becomes normalized over time [97]. However, maternal insults enable the interruption of this normalization and improperly initiate the classic RAS, leading to kidney disease and hypertension in adult offspring [96,97].
The classic RAS axis is activated in melatonin-deficient hypertension [49]. BP-lowering effects have been reported in studies using melatonin in pregnancy and lactation, together with blocking the RAS, in animal models of gestational chronodisruption [63], maternal continuous light exposure [65], maternal caloric restriction [66], and maternal high-fructose diet [69]. Another study revealed that maternal melatonin therapy prevented offspring hypertension programmed by antenatal dexamethasone administration plus post-weaning high-fat diet, which coincided with an enhanced non-classic RAS axis by increasing renal Agtr1b and Mas1 expression [72]. These observations above reveal a crosstalk between the RAS and melatonin behind renal programming, while additional research is necessary to confirm that the renoprotective actions of melatonin are directly RAS-dependent.
4.7. Gut Microbiota Dysbiosis
Another renoprotective mechanism of melatonin against renal programming might be due to its capacity to shape gut microbiota. The gut is a rich source of extrapineal melatonin with a ~400 times higher melatonin concentration in the gut than in the pineal gland [98]. Melatonin is a tryptophan-derived metabolite. Of note is that many tryptophan metabolites derived from gut microbiota participate in the developmental programming of kidney disease [99].
Emerging evidence supports gut microbiota dysbiosis in early life having adverse effects resulting in diseases in adulthood [100], such as kidney disease [101]. As reviewed elsewhere [101], a pathogenic interconnection, namely the gut–kidney axis, between the gut microbiota and kidney diseases is implicated in CKD and its comorbidities. Disturbed microbiota compositions and microbial metabolites are involved in pathogenesis. These metabolites include short chain fatty acids (SCFAs), trimethylamine-N-oxide (TMAO), tryptophan-derived uremic toxins, etc. [102,103,104,105].
A study investigating the effect of melatonin therapy for weeks in young CKD rats found that CKD-primed hypertension and kidney damage was prevented by melatonin [76]. The beneficial action of melatonin was accompanied by alterations in gut microbiota, including increased α-diversity, enhancement of the abundance of the phylum Proteobacteria and the genus Roseburia, and an improved TMAO metabolic pathway. Considering that several gut microbiota-targeted therapies have been applied for early prevention of CKD [101], a better understanding of how melatonin mediates gut microbiota underlying renal programming needs to be evaluated further.
4.8. Others
In view of the multifaceted actions of melatonin, there might be other mechanisms by which it provides an advantage: (1) by counteracting glucocorticoid programming, (2) by activating nuclear factor erythroid 2-related factor 2 (Nrf2), and (3) by regulating mitochondrial function. Similar to melatonin, glucocorticoid participates in the circadian rhythm [106]. Glucocorticoid and melatonin can downregulate each other’s receptors [49,107]. Considering that melatonin therapy prevented glucocorticoid programming-induced hypertension, the interplay between melatonin and glucocorticoid on renal programming deserves further elucidation. Additionally, melatonin can act like an Nrf2 activator [108]. In this regard, the coupling of melatonin and its metabolites to the activation of Nrf2 has been demonstrated in various organ systems [109,110,111,112]. Prior work indicated that Nrf2 activation has benefits for the developmental programming of hypertension [92,113]. Although this remains speculative, Nrf2 and other potential mechanisms are awaiting further clarification.
Melatonin can be specifically targeted to the mitochondria, where it acts as an antioxidant [114]. In mitochondria, cytochrome c is a natural scavenger of H2O2, preventing its accumulation. When electron transport is disrupted, the cytochrome c-dependent pseudo-peroxidase reaction with melatonin could become dominant to exhibit a protective mechanism [115]. Considering substantial evidence that has accumulated on the protective role of mitochondrial targeting against kidney disease [116], it is interesting to know how melatonin would coordinate mitochondrial interactions with the developing kidney to later determine kidney health and disease.
5. Pending Issues and Future Directions
If melatonin is be used as a potential intervention in pregnant women to prevent their offspring’s kidney disease, one major concern is its safety. A recent review revealed that a total of seven of the pregnancy studies and three of the lactation studies involved exogenous melatonin [117]. The dose of melatonin utilized in these studies ranged from 8 to 30 mg daily. No major adverse events or safety concerns were stated in most studies [117], except for one case of reported bleeding [118]. Nevertheless, currently there is a lack of clinical trials of melatonin use during pregnancy and lactation, especially trials related to offspring outcomes. According to the available evidence, no conclusions can be made about the safety of exogenously administered melatonin during gestation and lactation on the long-term outcome for the babies perinatally exposed.
Another pending question that needs to be addressed is the ideal dose and administration route. Most human studies using 10 mg of daily melatonin are not adequate to provide an adequate comparison with data on the therapeutic dose derived from animal studies [119]. The doses of melatonin used should be evaluated in the 40–100 mg/day range in view of the equivalent human doses of melatonin based on preclinical data. Notably, melatonin acts as a pro-oxidant at very high concentration (1–10 mM) [120]. In this situation, melatonin cannot be considered as a hormone but a context-dependent regulator. Considering that melatonin might display pro-oxidant activity when used at high concentrations [120], there is an utmost need for further research to recommend a maximum dose of melatonin in humans.
Melatonin is traditionally administered orally, while its drawback is low bioavailability due to fast release. Consequently, melatonin-sustained release formulations to humans via oral, intranasal, transdermal, and transmucosal administrations have been developed [121]. Nevertheless, whether these routes are suitable for pregnancy and whether melatonin’s pharmacokinetics are different in pregnant women are still not known [121].
Currently, melatonin might be the best peripheral biomarker for the circadian clock [122]. The onset of melatonin secretion under dim light conditions is the single most accurate marker for assessing the circadian pacemaker [122]. Accordingly, there have been several assays developed to analyze melatonin in blood and saliva for this aspect [123]. As its daytime physiological level is very low, there is a need for a specific assay for melatonin that is sensitive enough to detect low concentrations (<2 pg/mL). To date, several analytical methods for the quantitative measurement of melatonin concentrations in the plasma and saliva have been developed, including liquid chromatography with mass spectrometric detection (LC)–MS, gas chromatography (GC)–MS, radioimmunoassay (RIA), and enzyme-linked immunosorbent assay (ELISA) [123]. In clinical studies, ELISA and RIA are the most commonly used methods. However, many studies have been considered either meaningless or flawed due to extremely high levels caused by poor assay specificity [123]. Given that people who have no pineal gland display extremely low (<1 pg/mL) circulating levels of melatonin [124] and that melatonin level varies by age, with the lowest levels in young infants [125], future work in developing a simple high-precision method for determining melatonin in clinical practice is of utmost necessity.
Melatonin can be identified in both animal foods and edible plants [126]. A related question is whether dietary melatonin in pregnancy is appropriate for an efficient protection mechanism for offspring’s kidney disease The Mediterranean diet has been recommended for patients with CKD [127]. Part of this beneficial action might be attributed to the components of Mediterranean diet, which cover several melatonin-rich foods such as fish, red wine, olives, nuts, and fruits. Hence, some melatonin-rich foods might be of great value for development into functional foods, which would contribute to the prevention and treatment of kidney diseases.
As melatonin has pleiotropic biofunctions, its protective actions are difficult to predict or evaluate with a holistic approach in an experiment. Is there a dose-dependent mechanism behind the reprogramming effect of melatonin? Which protective mechanism might be most important? If so, what is the efficient dose and when should melatonin be implemented, and in which way, to mediate a specific protection mechanism? All these questions are still open-ended. Therefore, additional work in developing an ideal methodology is required to obtain a full-scope view of its protective mechanisms to ensure that melatonin therapy would only apply in the right direction.
6. Concluding Remarks and Perspectives
There is substantial evidence that melatonin participates in the pathophysiology of kidney health and disease. Although the use of melatonin as a potential preventive strategy is promising in preclinical studies, more work needs to be done to deliver it clinically.
So far, safety and efficacy data are largely lacking regarding the use of melatonin during gestation and breastfeeding and its reprogramming effects on offspring’s kidney disease. This review emphasizes the need for clinical studies in this aspect, including on exogenous melatonin, during pregnancy and lactation. Additionally, it is crucial to establish guidelines for the clinical use of melatonin for pregnant and lactating women.
In conclusion, melatonin contributes significantly to kidney health. Melatonin therapy in pregnancy and lactation can serve as a reprogramming strategy to prevent kidney disease while clinical translation is pending. Our review highlights a new path for the use of melatonin in working towards reducing the global burden of kidney disease.
Author Contributions
Funding acquisition, Y.-L.T.; conceptualization, Y.-L.T., K.-S.T., C.-Y.H. and C.-N.H.; data curation, Y.-L.T., K.-S.T., C.-Y.H. and C.-N.H.; writing—original draft, Y.-L.T., K.-S.T., C.-Y.H. and C.-N.H.; writing—review and editing, Y.-L.T., K.-S.T., C.-Y.H. and C.-N.H. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Kaohsiung Chang Gung Memorial Hospital, Kaohsiung, Taiwan, under grants CMRPG8N0171, CORPG8N0091 and CORPG8N0121.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Armstrong, S.M. Melatonin and circadian control in mammals. Experientia 1989, 45, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and inflammation—Story of a double-edged blade. J. Pineal Res. 2018, 65, e12525. [Google Scholar] [CrossRef]
- Ekmekcioglu, C. Melatonin receptors in humans: Biological role and clinical relevance. Biomed. Pharmacother. 2006, 60, 97–108. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Patel, K.K.; Dehari, D.; Agrawal, A.K.; Singh, S. Melatonin and its ubiquitous anticancer effects. Mol. Cell Biochem. 2019, 462, 133–155. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Nakamura, Y.; Terron, M.P.; Flores, L.J.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and pregnancy in the human. Reprod. Toxicol. 2008, 25, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Voiculescu, S.E.; Zygouropoulos, N.; Zahiu, C.D.; Zagrean, A.M. Role of melatonin in embryo fetal development. J. Med. Life 2014, 7, 488–492. [Google Scholar]
- Minich, D.M.; Henning, M.; Darley, C.; Fahoum, M.; Schuler, C.B.; Frame, J. Is Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients 2022, 14, 3934. [Google Scholar] [CrossRef]
- Pohanka, M. New Uses of Melatonin as a Drug; A Review. Curr. Med. Chem. 2022, 29, 3622–3637. [Google Scholar] [CrossRef]
- Hartz, I.; Handal, M.; Tverdal, A.; Skurtveit, S. Paediatric Off-Label Use of Melatonin--A Register Linkage Study between the Norwegian Prescription Database and Patient Register. Basic Clin. Pharmacol. Toxicol. 2015, 117, 267–273. [Google Scholar] [CrossRef]
- Gitto, E.; Aversa, S.; Reiter, R.J.; Barberi, I.; Pellegrino, S. Update on the use of melatonin in pediatrics. J. Pineal Res. 2011, 50, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Tonelli, M.; Stanifer, J.W. The global burden of kidney disease and the sustainable development goals. Bull. World Health Organ. 2018, 96, 414D–422D. [Google Scholar] [CrossRef]
- Haugen, A.C.; Schug, T.T.; Collman, G.; Heindel, J.J. Evolution of DOHaD: The impact of environmental health sciences. J. Dev. Orig. Health Dis. 2014, 6, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, R.L. Evolution, kidney development, and chronic kidney disease. Semin. Cell Dev. Biol. 2019, 91, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Luyckx, V.A.; Bertram, J.F.; Brenner, B.M.; Fall, C.; Hoy, W.E.; Ozanne, S.E.; Vikse, B.E. Effect of fetal and child health on kidney development and long-term risk of hypertension and kidney disease. Lancet 2013, 382, 273–283. [Google Scholar] [CrossRef]
- Paauw, N.D.; van Rijn, B.B.; Lely, A.T.; Joles, J.A. Pregnancy as a critical window for blood pressure regulation in mother and child: Programming and reprogramming. Acta Physiol. 2017, 219, 241–259. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. The First Thousand Days: Kidney Health and Beyond. Healthcare 2021, 9, 1332. [Google Scholar] [CrossRef]
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef]
- Sato, K.; Meng, F.; Francis, H.; Wu, N.; Chen, L.; Kennedy, L.; Zhou, T.; Franchitto, A.; Onori, P.; Gaudio, E.; et al. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J. Pineal Res. 2020, 68, e12639. [Google Scholar] [CrossRef] [PubMed]
- Odinokov, D.; Hamblin, M.R. Aging of lymphoid organs: Can photobiomodulation reverse age-associated thymic involution via stimulation of extrapineal melatonin synthesis and bone marrow stem cells? J. Biophotonics 2018, 11, e201700282. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Slominski, R.M.; Raman, C.; Chen, J.Y.; Athar, M.; Elmets, C. Neuroendocrine signaling in the skin with a special focus on the epidermal neuropeptides. Am. J. Physiol. Cell Physiol. 2022, 323, C1757–C1776. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Tan, D.X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef]
- Faria, J.A.; Kinote, A.; Ignacio-Souza, L.M.; de Araujo, T.M.; Razolli, D.S.; Doneda, D.L.; Paschoal, L.B.; Lellis-Santos, C.; Bertolini, G.L.; Velloso, L.A.; et al. Melatonin acts through MT1/MT2 receptors to activate hypothalamic Akt and suppress hepatic gluconeogenesis in rats. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E230–E242. [Google Scholar] [CrossRef]
- Ha, E.; Yim, S.V.; Chung, J.H.; Yoon, K.S.; Kang, I.; Cho, Y.H.; Baik, H.H. Melatonin stimulates glucose transport via insulin receptor substrate-1/phosphatidylinositol 3-kinase pathway in C2C12 murine skeletal muscle cells. J. Pineal Res. 2006, 41, 67–72. [Google Scholar] [CrossRef]
- Tosini, G.; Owino, S.; Guillaume, J.L.; Jockers, R. Understanding melatonin receptor pharmacology: Latest insights from mouse models, and their relevance to human disease. Bioessays 2014, 36, 778–787. [Google Scholar] [CrossRef]
- Ma, H.; Kang, J.; Fan, W.; He, H.; Huang, F. ROR: Nuclear Receptor for Melatonin or Not? Molecules 2021, 26, 2693. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.K.; Takeda, Y.; Janjetovic, Z.; Brozyna, A.A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.C.; et al. RORα and ROR γ are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. FASEB J. 2014, 28, 2775–2789. [Google Scholar] [CrossRef] [PubMed]
- Costello, H.M.; Johnston, J.G.; Juffre, A.; Crislip, G.R.; Gumz, M.L. Circadian clocks of the kidney: Function, mechanism, and regulation. Physiol. Rev. 2022, 102, 1669–1701. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.A.; Studier, E.H.; Stallone, J.N.; Kennedy, C.M. Effects of melatonin (in water metabolism and renal function in male Syrian hamsters (Mesocricetus auratus). J. Pineal Res. 1992, 13, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, R.; Douma, L.G.; Scindia, Y.; Gumz, M.L. Circadian rhythms and renal pathophysiology. J. Clin. Investig. 2022, 132, e148277. [Google Scholar] [CrossRef] [PubMed]
- Ejaz, H.; Figaro, J.K.; Woolner, A.M.F.; Thottakam, B.M.V.; Galley, H.F. Maternal Serum Melatonin Increases During Pregnancy and Falls Immediately After Delivery Implicating the Placenta as a Major Source of Melatonin. Front. Endocrinol. 2021, 11, 623038. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Okamoto, K.; Hayashi, K.; Wakatsuki, A.; Tamura, S.; Sagara, Y. Maternal-fetal transfer of melatonin in pregnant women near term. J. Pineal Res. 1998, 25, 129–134. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Rosales-Corral, S.A. Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology. Hum. Reprod. Update 2014, 20, 293–307. [Google Scholar] [CrossRef]
- Lanoix, D.; Beghdadi, H.; Lafond, J.; Vaillancourt, C. Human placental trophoblasts synthesize melatonin and express its receptors. J. Pineal Res. 2008, 45, 50–60. [Google Scholar] [CrossRef]
- Lanoix, D.; Guérin, P.; Vaillancourt, C. Placental melatonin production and melatonin receptor expression are altered in preeclampsia: New insights into the role of this hormone in pregnancy. J. Pineal Res. 2012, 53, 417–425. [Google Scholar] [CrossRef]
- Vaněček, J. The Melatonin Receptors in Rat Ontogenesis. Neuroendocrinology 1988, 48, 201–203. [Google Scholar] [CrossRef]
- Wu, Y.H.; Zhou, J.N.; Balesar, R.; Unmehopa, U.; Bao, A.; Jockers, R.; Van Heerikhuize, J.; Swaab, D.F. Distribution of MT1 melatonin receptor immunoreactivity in the human hypothalamus and pituitary gland: Colocalization of MT1 with vasopressin, oxytocin, and corticotropin-releasing hormone. J. Comp. Neurol. 2006, 499, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, D.; Achten, C.; Dallmann, F.; Oster, H. Embryonic development and maternal regulation of murine circadian clock function. Chronobiol. Int. 2015, 32, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Mendez, N.; Abarzua-Catalan, L.; Vilches, N.; Galdames, H.A.; Spichiger, C.; Richter, H.G.; Valenzuela, G.J.; Seron-Ferre, M.; Torres-Farfan, C. Timed maternal melatonin treatment reverses circadian disruption of the fetal adrenal clock imposed by exposure to constant light. PLoS ONE 2012, 7, e42713. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.; Sampogna, R.V.; Sakurai, H.; Bush, K.T.; Nigam, S.K. Branching morphogenesis and kidney disease. Development 2004, 131, 1449–1462. [Google Scholar] [CrossRef] [PubMed]
- Bertram, J.F.; Douglas-Denton, R.N.; Diouf, B.; Hughson, M.; Hoy, W. Human nephron number: Implications for health and disease. Pediatr. Nephrol. 2011, 26, 1529–1533. [Google Scholar] [CrossRef]
- Murugapoopathy, V.; Gupta, I.R. A primer on congenital anomalies of the kidneys and urinary tracts (CAKUT). Clin. J. Am. Soc. Nephrol. 2020, 15, 723–731. [Google Scholar] [CrossRef]
- Dan, H.; Ruan, T.; Sampogna, R.V. Circadian Clock Regulation of Developmental Time in the Kidney. Cell Rep. 2020, 31, 107661. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Chan, J.Y.; Lee, C.T. Transcriptome analysis in rat kidneys: Importance of genes involved in programmed hypertension. Int. J. Mol. Sci. 2015, 16, 4744–4758. [Google Scholar] [CrossRef]
- Simko, F.; Reiter, R.J.; Pechanova, O.; Paulis, L. Experimental models of melatonin-deficient hypertension. Front. Biosci. 2013, 18, 616–625. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Light and Circadian Signaling Pathway in Pregnancy: Programming of Adult Health and Disease. Int. J. Mol. Sci. 2020, 21, 2232. [Google Scholar] [CrossRef]
- Mendez, N.; Torres-Farfan, C.; Salazar, E.; Bascur, P.; Bastidas, C.; Vergara, K.; Spichiger, C.; Halabi, D.; Vio, C.; Richter, H.G. Fetal programming of renal dysfunction and high blood pressure by chronodisruption. Front. Endocrinol. 2019, 10, 362. [Google Scholar] [CrossRef] [PubMed]
- Baydaş, G.; Erçel, E.; Canatan, H.; Dönder, E.; Akyol, A. Effect of melatonin on oxidative status of rat brain, liver and kidney tissues under constant light exposure. Cell Biochem. Funct. 2001, 19, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Chan, J.Y. Transcriptional regulation of programmed hypertension by melatonin: An epigenetic perspective. Int. J. Mol. Sci. 2014, 15, 18484–18495. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Reiter, R.J. Epigenetic regulation: A new research area for melatonin? J. Pineal Res. 2008, 44, 41–44. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Developmental and Early Life Origins of Hypertension: Preventive Aspects of Melatonin. Antioxidants 2022, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Karamitri, A.; Jockers, R. Melatonin in type 2 diabetes mellitus and obesity. Nat. Rev. Endocrinol. 2019, 15, 105–125. [Google Scholar] [CrossRef]
- Dun, R.L.; Lan, T.Y.; Tsai, J.; Mao, J.M.; Shao, Y.Q.; Hu, X.H.; Zhu, W.J.; Qi, G.C.; Peng, Y. Protective Effect of Melatonin for Renal Ischemia-Reperfusion Injury: A Systematic Review and Meta-Analysis. Front. Physiol. 2022, 12, 791036. [Google Scholar] [CrossRef]
- Theofilis, P.; Vordoni, A.; Kalaitzidis, R.G. The Role of Melatonin in Chronic Kidney Disease and Its Associated Risk Factors: A New Tool in Our Arsenal? Am. J. Nephrol. 2022, 53, 565–574. [Google Scholar] [CrossRef]
- Maleki Dana, P.; Reiter, R.J.; Hallajzadeh, J.; Asemi, Z.; Mansournia, M.A.; Yousefi, B. Melatonin as a potential inhibitor of kidney cancer: A survey of the molecular processes. IUBMB Life 2020, 72, 2355–2365. [Google Scholar] [CrossRef]
- Clarke, T.C.; Black, L.I.; Stussman, B.J.; Barnes, P.M.; Nahin, R.L. Trends in the use of complementary health approaches among adults: United States, 2002–2012. Natl. Health Stat. Rep. 2015, 79, 1–16. [Google Scholar]
- Andersen, L.P.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2016, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Genario, R.; Morello, E.; Bueno, A.A.; Santos, H.O. The usefulness of melatonin in the field of obstetrics and gynecology. Pharmacol. Res. 2019, 147, 104337. [Google Scholar] [CrossRef] [PubMed]
- Mendez, N.; Halabi, D.; Salazar-Petres, E.R.; Vergara, K.; Corvalan, F.; Richter, H.G.; Bastidas, C.; Bascur, P.; Ehrenfeld, P.; Seron-Ferre, M.; et al. Maternal melatonin treatment rescues endocrine, inflammatory, and transcriptional deregulation in the adult rat female offspring from gestational chronodisruption. Front. Neurosci. 2022, 16, 1039977. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Sirajudeen, K.N.; Sundaram, A.; Zakaria, R.; Singh, H.J. Effects of antenatal, postpartum and post-weaning melatonin supplementation on blood pressure and renal antioxidant enzyme activities in spontaneously hypertensive rats. J. Physiol. Biochem. 2011, 67, 249–257. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lin, Y.J.; Chan, J.Y.H.; Lee, C.T.; Hsu, C.N. Maternal melatonin or agomelatine therapy prevents programmed hypertension in male offspring of mother exposed to continuous light. Biol. Reprod. 2017, 97, 636–643. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Hsu, C.N.; Lee, C.T. Melatonin therapy prevents programmed hypertension and nitric oxide deficiency in offspring exposed to maternal caloric restriction. Oxid. Med. Cell Longev. 2014, 2014, 283180. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal NG-Nitro-L-arginine-methyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636. [Google Scholar] [CrossRef]
- Tain, Y.L.; Chan, J.Y.H.; Lee, C.T.; Hsu, C.N. Maternal Melatonin Therapy Attenuates Methyl-Donor Diet-Induced Programmed Hypertension in Male Adult Rat Offspring. Nutrients 2018, 10, E1407. [Google Scholar] [CrossRef]
- Tain, Y.L.; Leu, S.; Wu, K.L.; Lee, W.C.; Chan, J.Y. Melatonin prevents maternal fructose intake-induced programmed hypertension in the offspring: Roles of nitric oxide and arachidonic acid metabolites. J. Pineal Res. 2014, 57, 80–89. [Google Scholar] [CrossRef]
- Tain, Y.L.; Leu, S.; Lee, W.C.; Wu, K.L.H.; Chan, J.Y.H. Maternal Melatonin Therapy Attenuated Maternal High-Fructose Combined with Post-Weaning High-Salt Diets-Induced Hypertension in Adult Male Rat Offspring. Molecules 2018, 23, E886. [Google Scholar] [CrossRef]
- Tain, Y.L.; Chen, C.C.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Kuo, H.C.; Huang, L.T. Melatonin attenuates prenatal dexamethasone-induced blood pressure increase in a rat model. J. Am. Soc. Hypertens. 2014, 8, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Sheen, J.M.; Yu, H.R.; Chen, C.C.; Tiao, M.M.; Hsu, C.N.; Lin, Y.J.; Kuo, K.C.; Huang, L.T. Maternal Melatonin Therapy Rescues Prenatal Dexamethasone and Postnatal High-Fat Diet Induced Programmed Hypertension in Male Rat Offspring. Front. Physiol. 2015, 6, 377. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Lee, B.N.; Kim, J.S. Effects of maternal-melatonin treatment on open-field behaviors and hypertensive phenotype in spontaneously hypertensive rats’ (SHR) offspring. Exp. Anim. 2002, 51, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Tain, Y.L. Postnatal dexamethasone-induced programmed hypertension is related to the regulation of melatonin and its receptors. Steroids 2016, 108, 1–6. [Google Scholar] [CrossRef]
- Wu, T.H.; Kuo, H.C.; Lin, I.C.; Chien, S.J.; Huang, L.T.; Tain, Y.L. Melatonin prevents neonatal dexamethasone induced programmed hypertension: Histone deacetylase inhibition. J. Steroid Biochem. Mol. Biol. 2014, 144, 253–259. [Google Scholar] [CrossRef]
- Hsu, C.N.; Yang, H.W.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Melatonin Prevents Chronic Kidney Disease-Induced Hypertension in Young Rat Treated with Adenine: Implications of Gut Microbiota-Derived Metabolites. Antioxidants 2021, 10, 1211. [Google Scholar] [CrossRef]
- Cheng, M.C.; Wu, T.H.; Huang, L.T.; Tain, Y.L. Renoprotective effects of melatonin in young spontaneously hypertensive rats with L-NAME. Pediatr. Neonatol. 2014, 55, 189–195. [Google Scholar] [CrossRef]
- Tain, Y.L.; Huang, L.T.; Lin, I.C.; Lau, Y.T.; Lin, C.Y. Melatonin prevents hypertension and increased asymmetric dimethylarginine in young spontaneous hypertensive rats. J. Pineal Res. 2010, 49, 390–398. [Google Scholar] [CrossRef]
- Lemley, C.O.; Vonnahme, K.A. Physiology and endocrinology symposium: Alterations in uteroplacental hemodynamics during melatonin supplementation in sheep and cattle. J. Anim. Sci. 2017, 95, 2211–2221. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Wang, F.; Tian, X.; Ji, P.; Liu, G. Effects of melatonin administration on embryo implantation and offspring growth in mice under different schedules of photoperiodic exposure. Reprod. Biol. Endocrinol. 2017, 15, 78. [Google Scholar] [CrossRef]
- Sengupta, P. The Laboratory Rat: Relating Its Age with Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Baylis, C. Nitric oxide synthase derangements and hypertension in kidney disease. Curr. Opin. Nephrol. Hypertens. 2012, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carlström, M. Nitric oxide signalling in kidney regulation and cardiometabolic health. Nat. Rev. Nephrol. 2021, 17, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Kao, Y.H.; Hsieh, C.S.; Chen, C.C.; Sheen, J.M.; Lin, I.C.; Huang, L.T. Melatonin blocks oxidative stress-induced increased asymmetric dimethylarginine. Free Radic. Biol. Med. 2010, 49, 1088–1098. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Perinatal Oxidative Stress and Kidney Health: Bridging the Gap between Animal Models and Clinical Reality. Antioxidants 2022, 12, 13. [Google Scholar] [CrossRef]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef]
- Huynh, C.; Ryu, J.; Lee, J.; Inoki, A.; Inoki, K. Nutrient-sensing mTORC1 and AMPK pathways in chronic kidney diseases. Nat. Rev. Nephrol. 2023, 19, 102–122. [Google Scholar] [CrossRef]
- Zha, D.; Wu, X. Nutrient sensing, signaling transduction, and autophagy in podocyte injury: Implications for kidney disease. J. Nephrol. 2023, 36, 17–29. [Google Scholar] [CrossRef]
- Jansson, T.; Powell, T.L. Role of placental nutrient sensing in developmental programming. Clin. Obstet. Gynecol. 2013, 56, 591–601. [Google Scholar] [CrossRef]
- Fernández, A.; Ordóñez, R.; Reiter, R.J.; González-Gallego, J.; Mauriz, J.L. Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 2015, 59, 292–307. [Google Scholar] [CrossRef]
- Promsan, S.; Thongnak, L.; Pengrattanachot, N.; Phengpol, N.; Sutthasupha, P.; Lungkaphin, A. Agomelatine, a structural analog of melatonin, improves kidney dysfunction through regulating the AMPK/mTOR signaling pathway to promote autophagy in obese rats. Food Chem. Toxicol. 2022, 165, 113190. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Lin, I.C.; Yu, H.R.; Sheen, J.M.; Huang, L.T.; Tain, Y.L. Early Postweaning Treatment with Dimethyl Fumarate Prevents Prenatal Dexamethasone- and Postnatal High-Fat Diet-Induced Programmed Hypertension in Male Rat Offspring. Oxid. Med. Cell Longev. 2018, 2018, 5343462. [Google Scholar] [CrossRef] [PubMed]
- Siddhi, J.; Sherkhane, B.; Kalavala, A.K.; Arruri, V.; Velayutham, R.; Kumar, A. Melatonin prevents diabetes-induced nephropathy by modulating the AMPK/SIRT1 axis: Focus on autophagy and mitochondrial dysfunction. Cell Biol. Int. 2022, 46, 2142–2157. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Ishigaki, S.; Isobe, S. The pivotal role of melatonin in ameliorating chronic kidney disease by suppression of the renin-angiotensin system in the kidney. Hypertens. Res. 2019, 42, 761–768. [Google Scholar] [CrossRef]
- Yosypiv, I.V. Renin-angiotensin system in mammalian kidney development. Pediatr. Nephrol. 2020, 36, 479–489. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Targeting the renin–angiotensin–aldosterone system to prevent hypertension and kidney disease of developmental origins. Int. J. Mol. Sci. 2021, 22, 2298. [Google Scholar] [CrossRef]
- Kett, M.M.; Denton, K. Renal programming: Cause for concern? Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R791–R803. [Google Scholar] [CrossRef]
- Bubenik, G.A. Gastrointestinal melatonin: Localization, function, and clinical relevance. Dig. Dis. Sci. 2002, 47, 2336–2348. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Developmental programming and reprogramming of hypertension and kidney disease: Impact of tryptophan metabolism. Int. J. Mol. Sci. 2020, 21, 8705. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, M.; Ueno, M.; Itoh, Y.; Suda, W.; Hattori, M. Uremic toxin-producing gut microbiota in rats with chronic kidney disease. Nephron 2017, 135, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Pluznick, J.L. Microbial short-chain fatty acids and blood pressure regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Al Khodor, S.; Reichert, B.; Shatat, I.F. The microbiome and blood pressure: Can microbes regulate our blood pressure? Front. Pediatr. 2017, 5, 138. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, M.T.; Ramezani, A.; Manal, A.; Raj, D.S. Trimethylamine N-Oxide: The good, the bad and the unknown. Toxins 2016, 8, 326. [Google Scholar] [CrossRef]
- Astiz, M.; Oster, H. Perinatal Programming of Circadian Clock-Stress Crosstalk. Neural. Plast. 2018, 2018, 5689165. [Google Scholar] [CrossRef]
- Cottrell, E.C.; Seckl, J.R. Prenatal stress, glucocorticoids and the programming of adult disease. Front. Behav. Neurosci. 2009, 3, 19. [Google Scholar] [CrossRef]
- Ahmadi, Z.; Ashrafizadeh, M. Melatonin as a potential modulator of Nrf2. Fundam. Clin. Pharmacol. 2020, 34, 11–19. [Google Scholar] [CrossRef]
- Janjetovic, Z.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites protect human melanocytes against UVB-induced damage: Involvement of NRF2-mediated pathways. Sci. Rep. 2017, 7, 1274. [Google Scholar] [CrossRef]
- Stefan, J.; Kim, T.K.; Schedel, F.; Janjetovic, Z.; Crossman, D.K.; Steinbrink, K.; Slominski, R.M.; Zmijewski, J.; Tulic, M.K.; Reiter, R.J.; et al. Differential and Overlapping Effects of Melatonin and Its Metabolites on Keratinocyte Function: Bioinformatics and Metabolic Analyses. Antioxidants 2021, 10, 618. [Google Scholar] [CrossRef]
- Joshi, A.; Upadhyay, K.K.; Vohra, A.; Shirsath, K.; Devkar, R. Melatonin induces Nrf2-HO-1 reprogramming and corrections in hepatic core clock oscillations in Non-alcoholic fatty liver disease. FASEB J. 2021, 35, e21803. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.H.; Ridlo, M.R.; Lee, B.C.; Kim, G.A. Melatonin-Nrf2 Signaling Activates Peroxisomal Activities in Porcine Cumulus Cell-Oocyte Complexes. Antioxidant 2020, 9, 1080. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Lin, Y.J.; Yu, H.R.; Lin, I.C.; Sheen, J.M.; Huang, L.T.; Tain, Y.L. Protection of Male Rat Offspring against Hypertension Programmed by Prenatal Dexamethasone Administration and Postnatal High-Fat Diet with the Nrf2 Activator Dimethyl Fumarate during Pregnancy. Int. J. Mol. Sci. 2019, 20, 3957. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.; Tan, D.X.; Jou, M.J.; Galano, A.; Xu, B. Melatonin as a mitochondria-targeted antioxidant: One of evolution’s best ideas. Cell Mol. Life Sci. 2017, 74, 3863–3881. [Google Scholar] [CrossRef] [PubMed]
- Semak, I.; Naumova, M.; Korik, E.; Terekhovich, V.; Wortsman, J.; Slominski, A. A novel metabolic pathway of melatonin: Oxidation by cytochrome c. Biochemistry 2005, 44, 9300–9307. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Lerman, A.; Lerman, L.O. The Emerging Role of Mitochondrial Targeting in Kidney Disease. Handb. Exp. Pharmacol. 2017, 240, 229–250. [Google Scholar]
- Vine, T.; Brown, G.M.; Frey, B.N. Melatonin use during pregnancy and lactation: A scoping review of human studies. Braz. J. Psychiatry. 2022, 44, 342–348. [Google Scholar] [CrossRef]
- Luciani, M.; Massoud, M.; Foligno, S.; Di Felice, G.; Pulcinelli, F.; Rapini, N. Melatonin antiplatelets effect through breastfeeding: A sobering case. Haemophilia 2019, 25, 138–139. [Google Scholar]
- Cardinali, D. Are melatonin doses employed clinically adequate for melatonin-induced cytoprotection? Melatonin Res. 2019, 2, 106–132. [Google Scholar] [CrossRef]
- Ossenia, R.A.; Rata, P.; Bogdan, A.; Warneta, J.M.; Touitou, Y. Evidence of prooxidant and antioxidant action of melatonin on human liver cell line HepG2. Life Sci. 2000, 68, 387–399. [Google Scholar] [CrossRef]
- Zetner, D.; Andersen, L.P.H.; Rosenberg, J. Pharmacokinetics of Alternative Administration Routes of Melatonin: A Systematic Review. Drug Res. 2015, 66, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Smits, M.; Spence, W.; Srinivasan, V.; Cardinali, D.P.; Lowe, A.D.; Kayumov, L. Dim light melatonin onset (DLMO): A tool for the analysis of circadian phase in human sleep and chronobiological disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2007, 31, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, D.J. A critical review of melatonin assays: Past and present. J. Pineal Res. 2019, 67, e12572. [Google Scholar] [CrossRef]
- Slawik, H.; Stoffel, M.; Riedl, L.; Veselý, Z.; Behr, M.; Lehmberg, J.; Pohl, C.; Meyer, B.; Wiegand, M.; Krieg, S.M. Prospective study on salivary evening melatonin and sleep before and after pinealectomy in humans. J. Biol. Rhythm. 2016, 31, 82–93. [Google Scholar] [CrossRef]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, Functions and Therapeutic Benefits. Curr. Neuropharmacol. 2017, 15, 434–443. [Google Scholar] [CrossRef]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.Y.; Xu, D.P.; Li, H.B. Dietary Sources and Bioactivities of Melatonin. Nutrients 2017, 9, 367. [Google Scholar] [CrossRef]
- Pérez-Torres, A.; Caverni-Muñoz, A.; González García, E. Mediterranean Diet and Chronic Kidney Disease (CKD): A Practical Approach. Nutrients 2022, 15, 97. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram highlighting the impact of melatonin in normal pregnancy and kidney development. Additionally, the use of melatonin in early life can protect against renal programming-induced adulthood kidney diseases later in life.
Figure 1.
Schematic diagram highlighting the impact of melatonin in normal pregnancy and kidney development. Additionally, the use of melatonin in early life can protect against renal programming-induced adulthood kidney diseases later in life.

Figure 2.
Therapeutic and preventive mechanisms of melatonin against renal programming. NO = nitric oxide; RAS = renin-angiotensin system.
Figure 2.
Therapeutic and preventive mechanisms of melatonin against renal programming. NO = nitric oxide; RAS = renin-angiotensin system.


{kind=link}
{kind=link}
Table 1.
Renoprotective effects of melatonin use in early life protect against early origins of kidney-related diseases in animal models.
Table 1.
Renoprotective effects of melatonin use in early life protect against early origins of kidney-related diseases in animal models.
| Dose and Treatment Period | Animal Model | Species/ Gender | Age at Evaluation | Renoprotective Effects and Mechanisms | Ref. |
|---|---|---|---|---|---|
| Gestation | |||||
| 2 μg/mL in drinking water from gestational day 0 to 18 | Gestational chronodisruption | SD rat/F | 90 days | Restored specific kidney genes related to BP control | [63] |
| 10 mg/kg/day in drinking water | Genetic hypertension | SHR/M | 8 weeks | Decreased the rise in BP | [64] |
| Gestation and lactation | |||||
| 0.01% in drinking water | Maternal continuous light exposure | SD rat/M | 12 weeks | Prevented hypertension and restored the aberrant RAS | [65] |
| 0.01% in drinking water | Maternal caloric restriction | SD rat/M | 12 weeks | Prevented hypertension, reduced oxidative stress, restored the aberrant RAS, and increased renal NO | [66] |
| 0.01% in drinking water | Maternal L-NAME exposure | SD rat/M | 12 weeks | Prevented hypertension, reduced oxidative stress, and increased renal NO | [67] |
| 0.01% in drinking water | Maternal high methyl-donor diet | SD rat/M | 12 weeks | Attenuated hypertension, reduced oxidative stress, and altered renal transcriptome | [68] |
| 0.01% in drinking water | Maternal high-fructose diet | SD rat/M | 12 weeks | Prevented hypertension, reduced oxidative stress, restored the aberrant RAS, and increased renal NO | [69] |
| 0.01% in drinking water | Maternal high-fructose diet plus post-weaning high-salt diet | SD rat/M | 12 weeks | Attenuated hypertension, restored NO system, and improved nutrient sensing signals | [70] |
| 0.01% in drinking water | Prenatal dexamethasone exposure | SD rat/M | 16 weeks | Prevented hypertension, increased nephron number, and reduced oxidative stress | [71] |
| 0.01% in drinking water | Prenatal dexamethasone exposure plus post-weaning high-fat diet | SD rat/M | 16 weeks | Prevented hypertension and restored the aberrant RAS | [72] |
| 20 μg/mL in drinking water | Genetic hypertension | SHR/M | 27 weeks | Prevented the rise in BP | [73] |
| Lactation | |||||
| 0.01% in drinking water | Neonatal dexamethasone exposure | SD rat/M | 16 weeks | Prevented hypertension and preserved histone deacetylase gene expression | [74] |
| 0.01% in drinking water | Neonatal dexamethasone exposure | SD rat/M | 16 weeks | Prevented hypertension, increased renal melatonin level, and upregulated MT2 protein expression | [75] |
| Early childhood | |||||
| 10 mg/kg/day in drinking water from 3 to 6 weeks of age | Adenine-induced CKD | SD rat/M and F | 9 weeks | Prevented hypertension, attenuated kidney injury, increased NO, and altered gut microbiota | [76] |
| 0.01% in drinking water from 4 to 10 weeks of age | Genetic hypertension plus L-NAME exposure | SHR/M | 10 weeks | Prevented hypertension, reduced renal oxidative stress and ADMA concentration | [77] |
| 0.01% in drinking water from 4 to 12 weeks of age | Genetic hypertension | SHR/M | 12 weeks | Prevented hypertension, reduced oxidative stress and plasma ADMA concentration | [78] |
L-NAME = NG-nitro-l-arginine methyl ester; CKD = chronic kidney disease; SD = Sprague Dawley rat; SHR = spontaneously hypertensive rat; M = male; F = female; NO = nitric oxide; RAS = renin-angiotensin system; BP = blood pressure; ADMA = asymmetric dimethylarginine; MT2 = melatonin receptor 2.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tang, K.-S.; Ho, C.-Y.; Hsu, C.-N.; Tain, Y.-L. Melatonin and Kidney Health: From Fetal Stage to Later Life. Int. J. Mol. Sci. 2023, 24, 8105. https://doi.org/10.3390/ijms24098105
AMA Style
Tang K-S, Ho C-Y, Hsu C-N, Tain Y-L. Melatonin and Kidney Health: From Fetal Stage to Later Life. International Journal of Molecular Sciences. 2023; 24(9):8105. https://doi.org/10.3390/ijms24098105
Chicago/Turabian StyleTang, Kuo-Shu, Chun-Yi Ho, Chien-Ning Hsu, and You-Lin Tain. 2023. "Melatonin and Kidney Health: From Fetal Stage to Later Life" International Journal of Molecular Sciences 24, no. 9: 8105. https://doi.org/10.3390/ijms24098105
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.