
Ca2+ Dependent Formation/Collapse of Cylindrical Ca2+-ATPase Crystals in Scallop Sarcoplasmic Reticulum (SR) Vesicles: A Possible Dynamic Role of SR in Regulation of Muscle Contraction
 ,
, 
Abstract
:1. Introduction
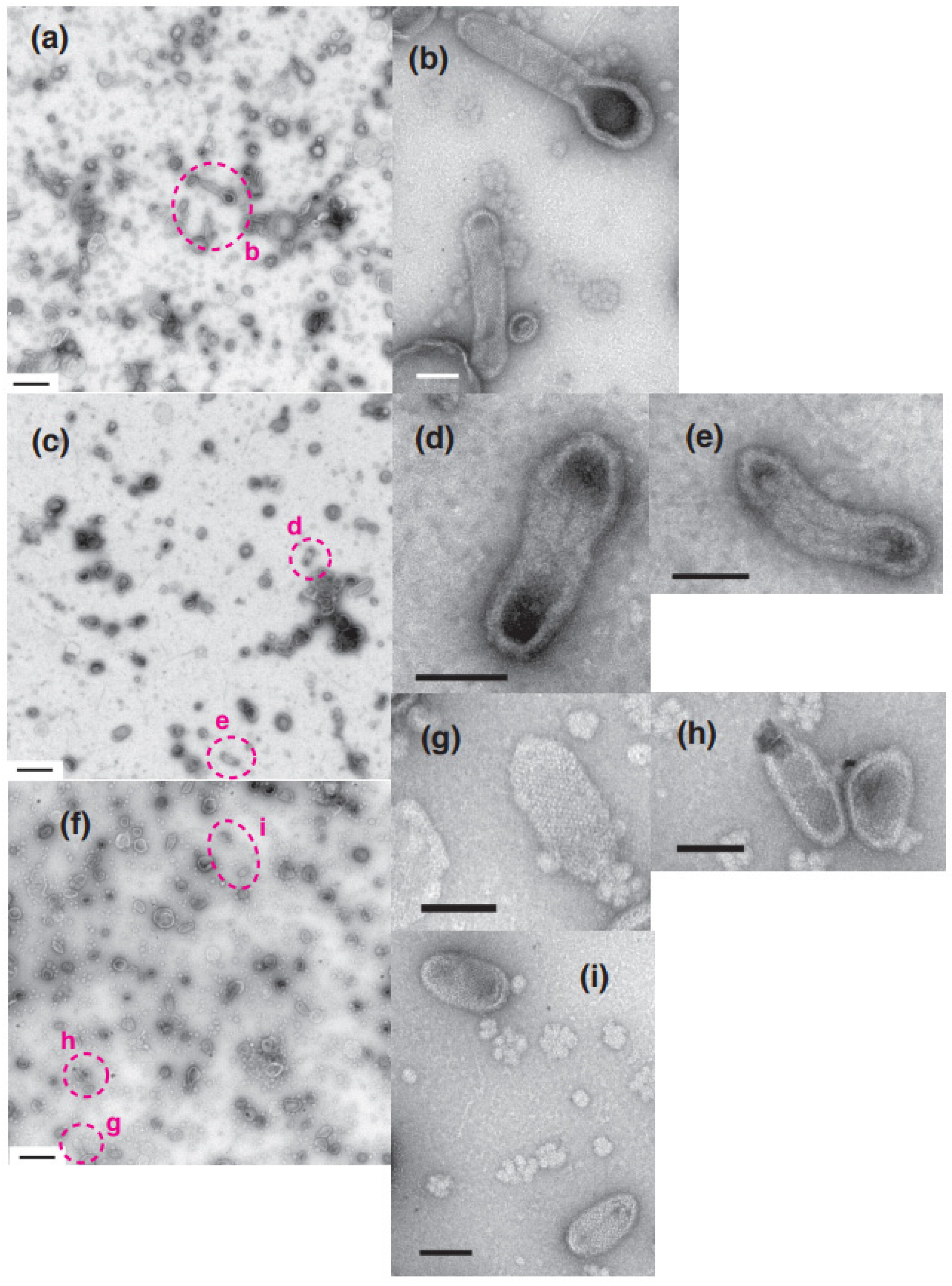
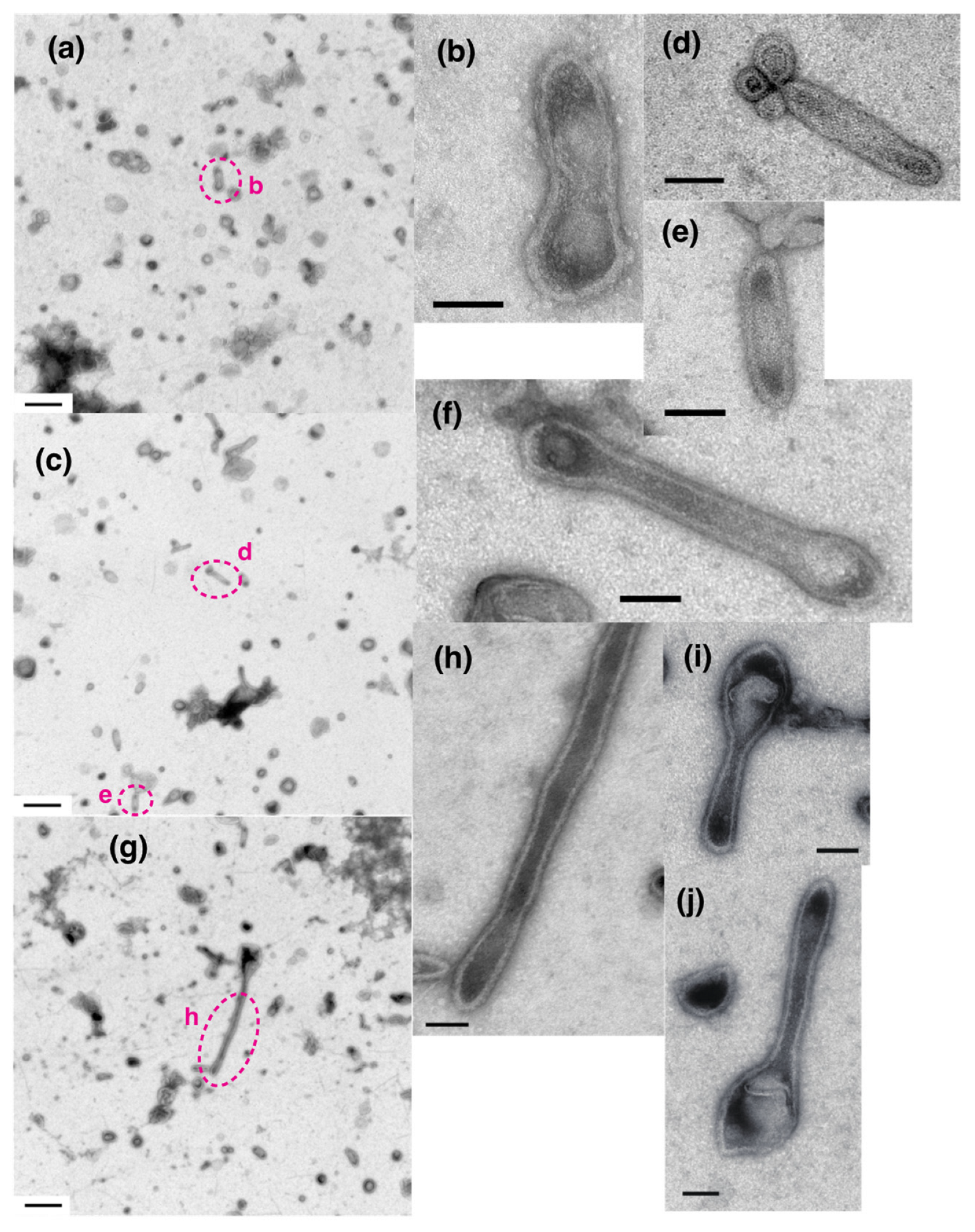
2. Results
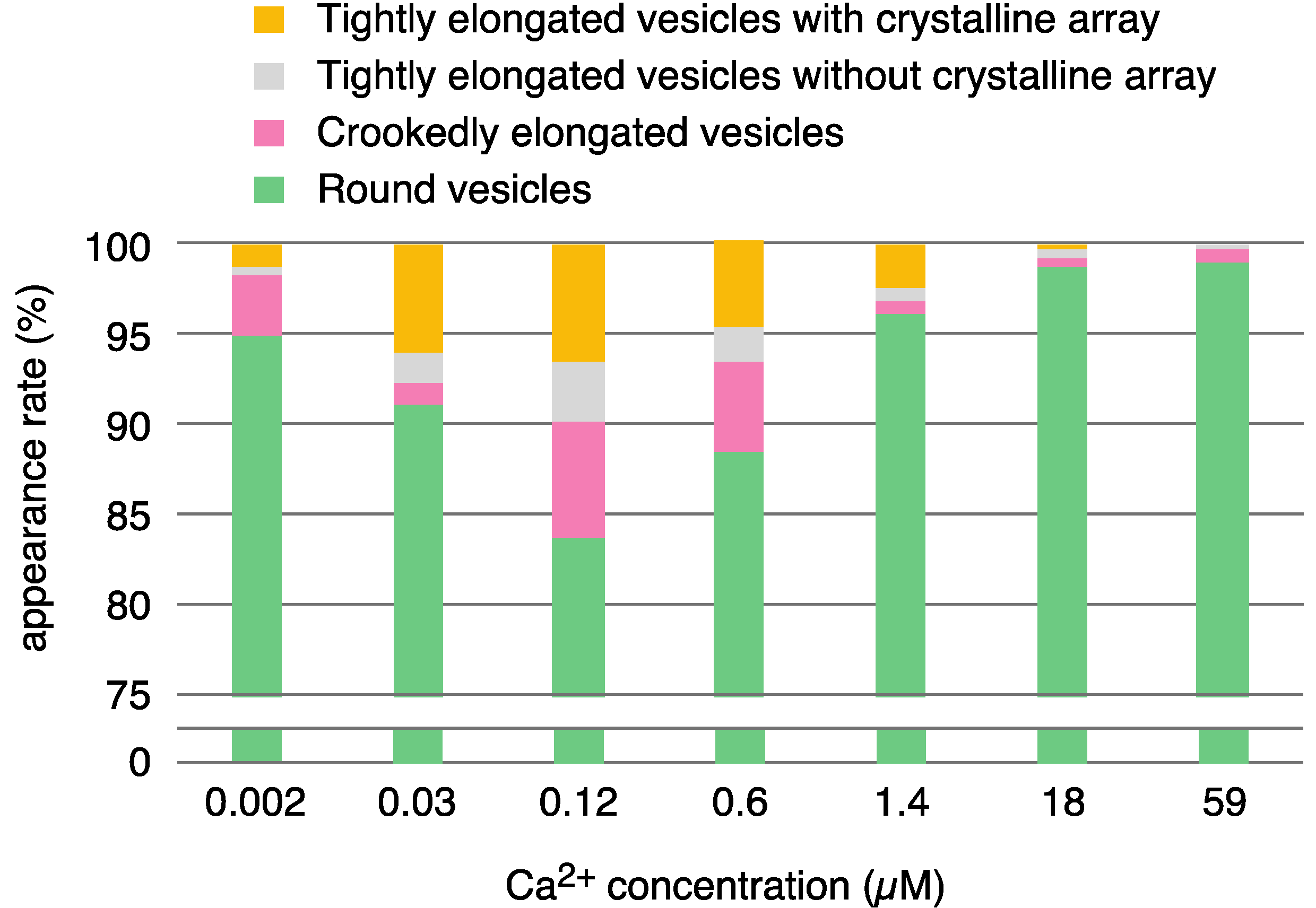
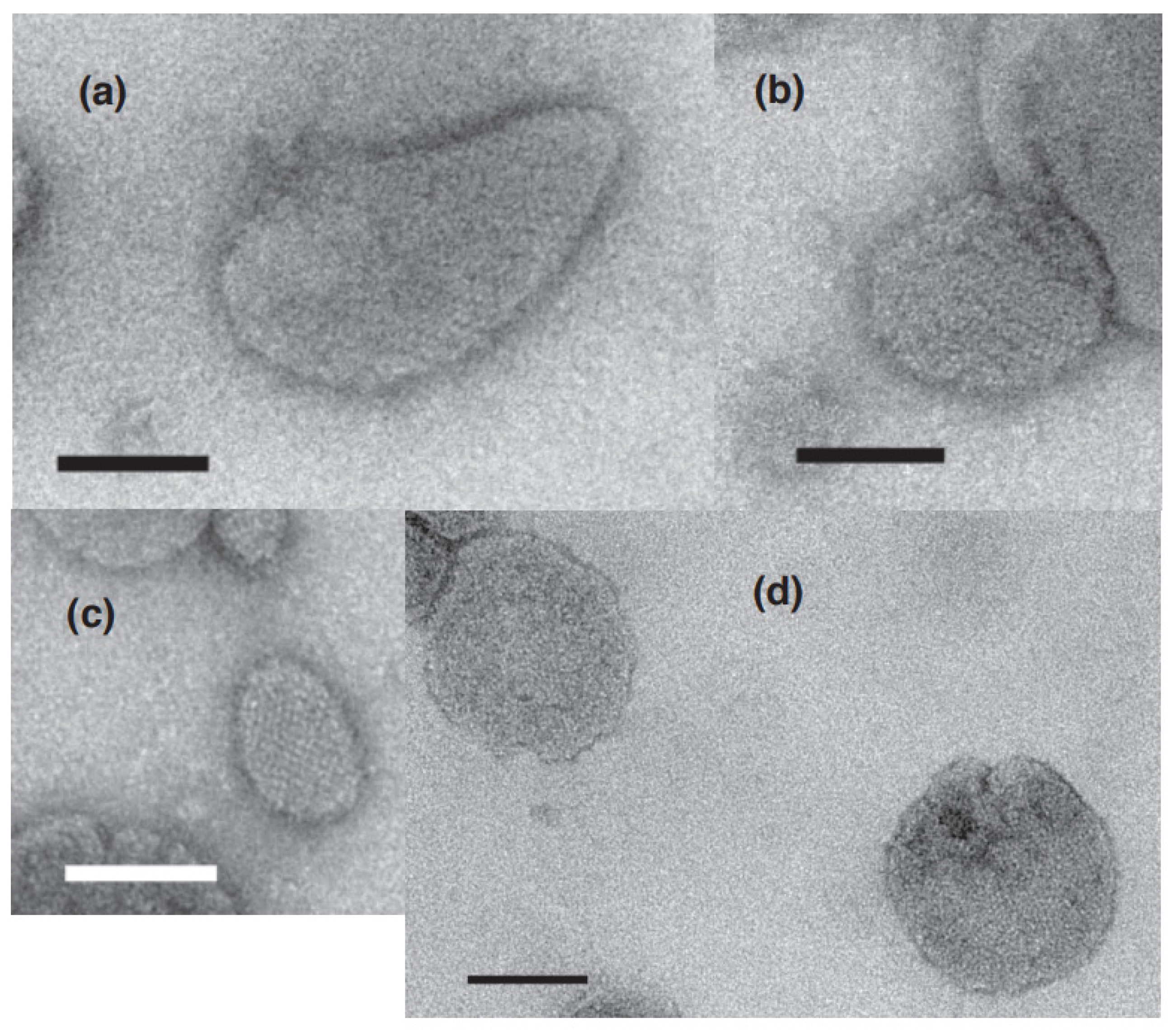
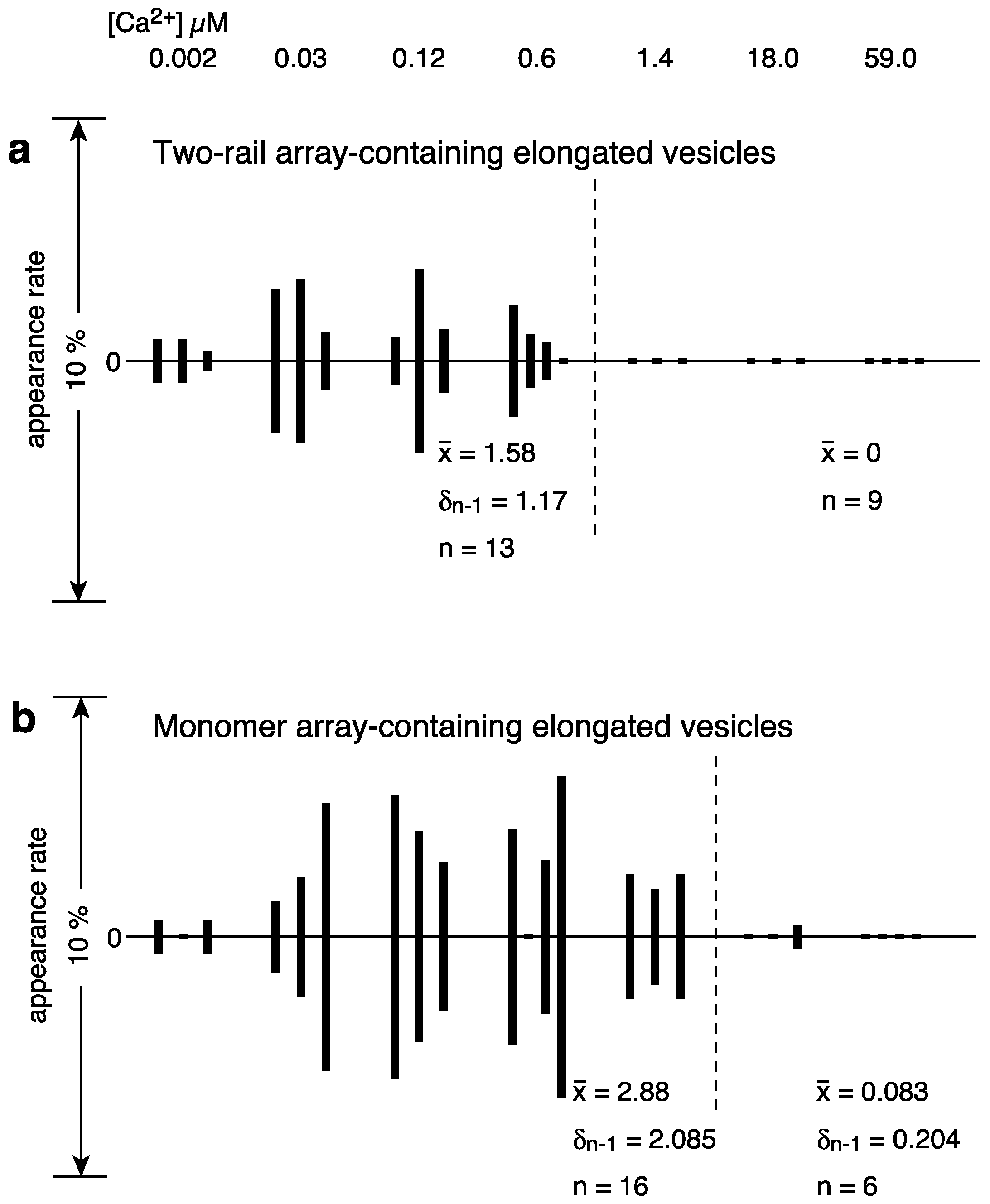
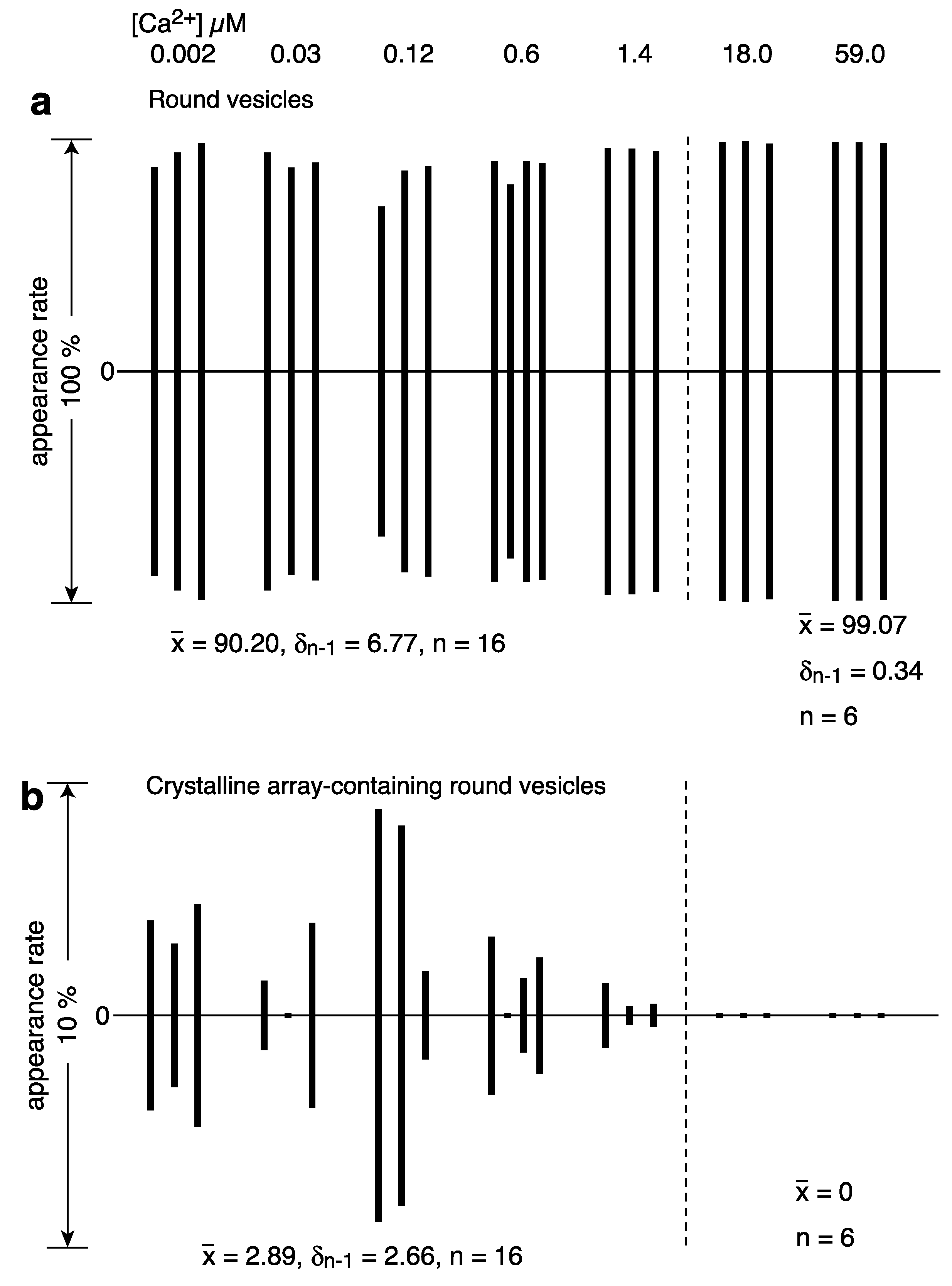
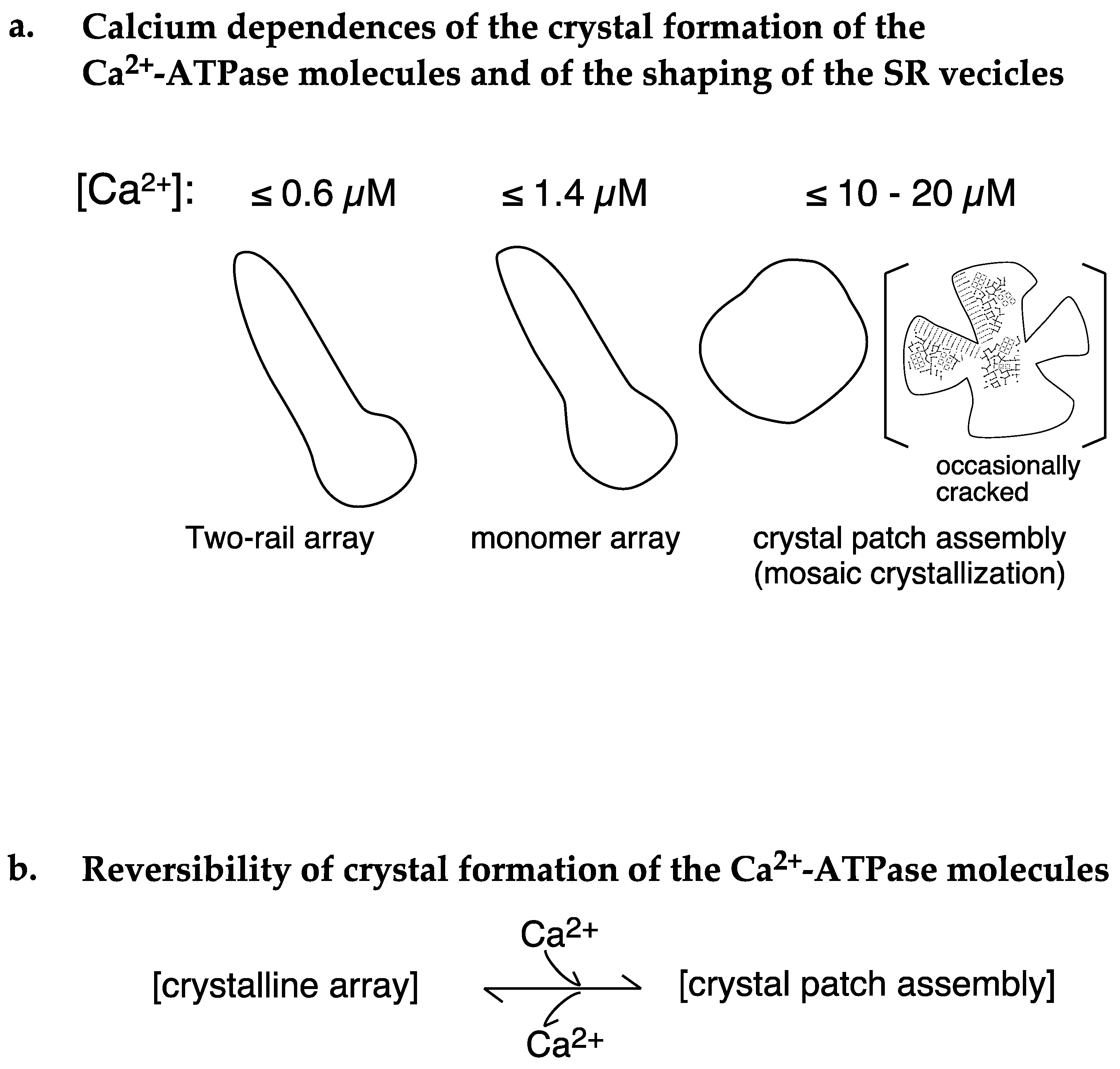
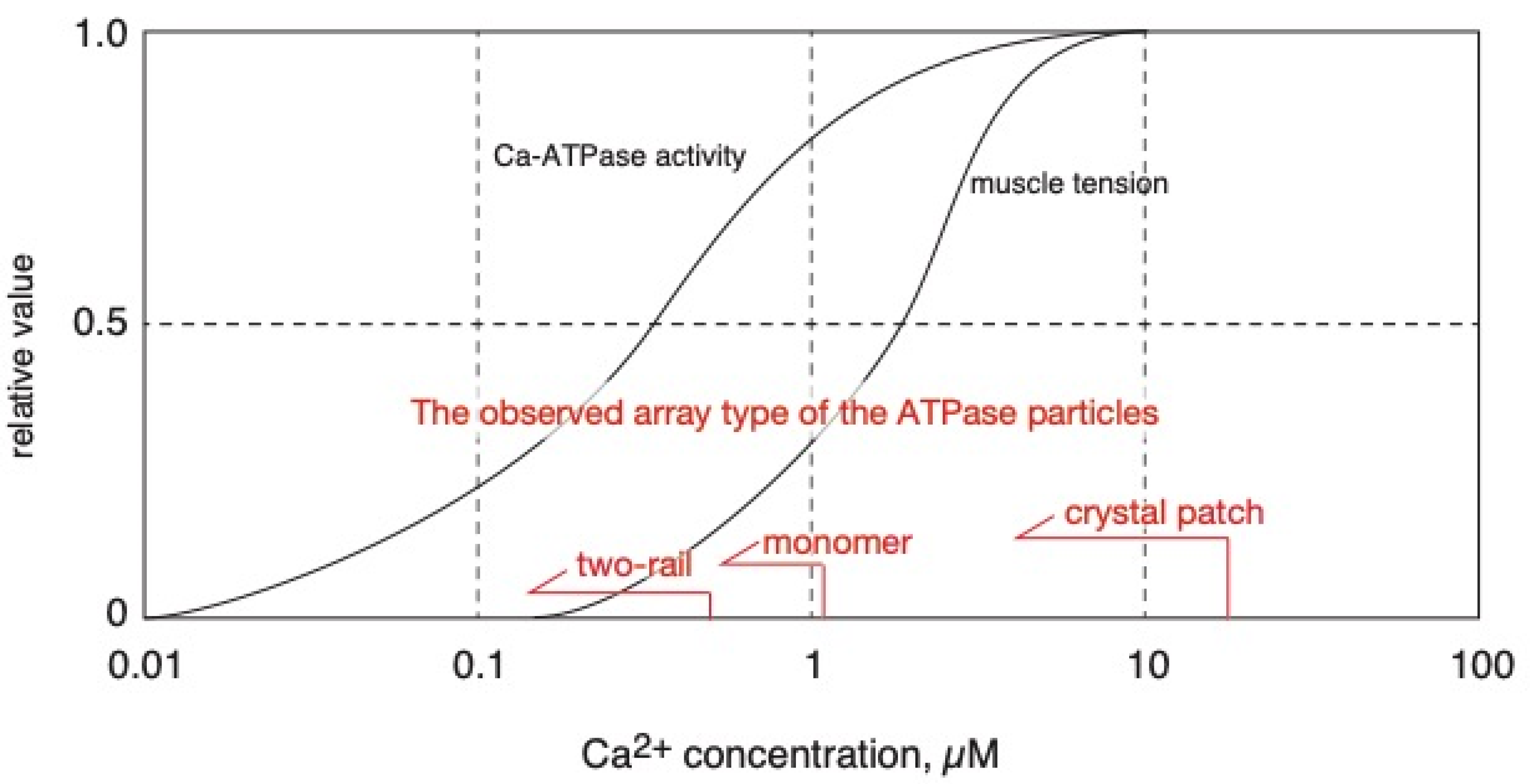
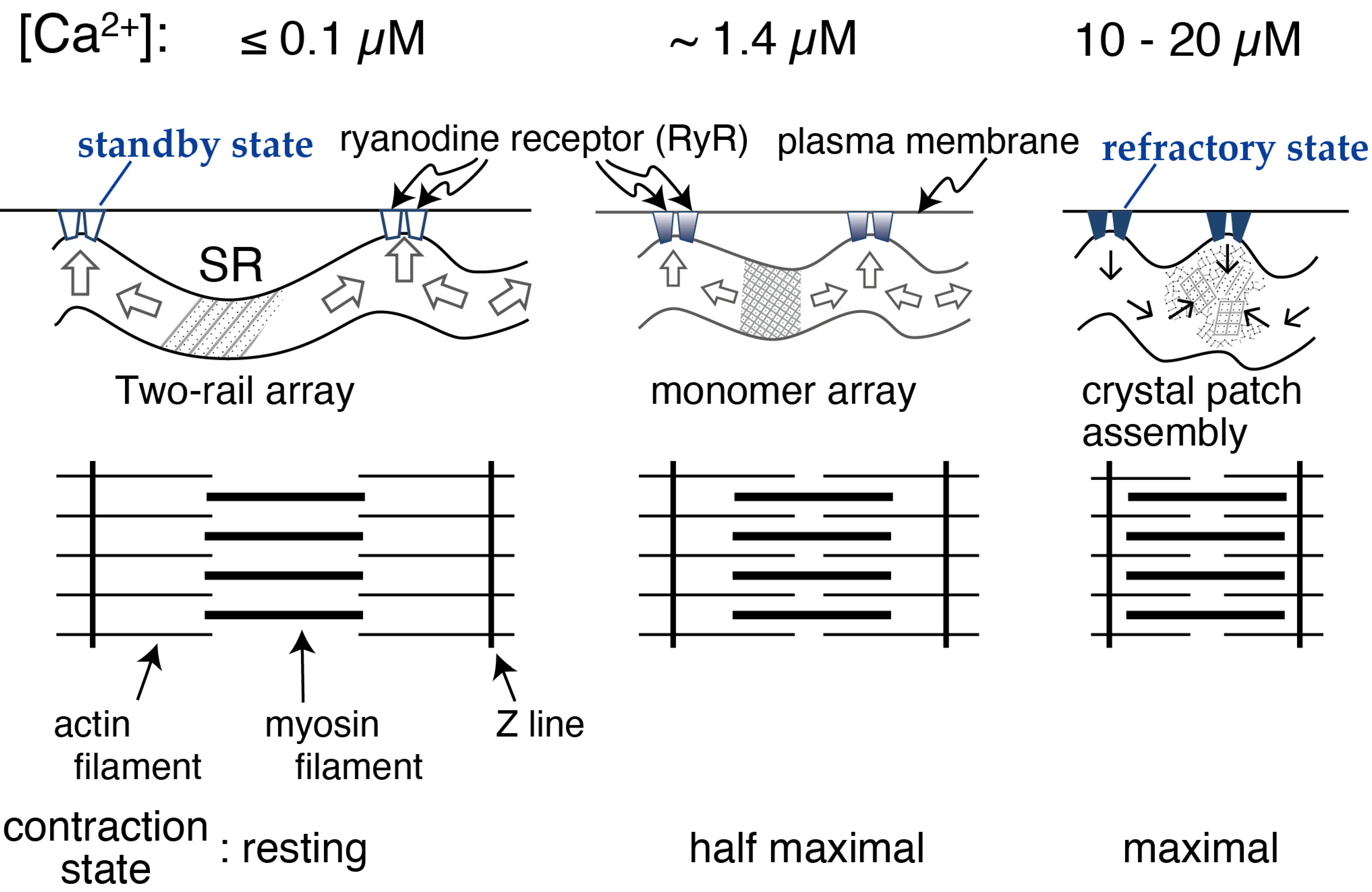
2.1. Calcium Dependence of the ATPase Crystallization and Elongation of SR Vesicles in the Presence of ATP
2.2. [Ca2+] Jump-Up and -Down
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SR | Sarcoplasmic reticulum |
| EGTA | ethylenebis (oxyethylenenitrilo) tetraacetic acid |
| TG | Thapsigargin |
| DMSO | dimethylsulfoxide |
| TEM | Transmission electron microscopy/microscope |
| SD | Standard deviation |
References
- Nakamura, J.; Maruyama, Y.; Tajima, G.; Komeiji, Y.; Suwa, M.; Sato, C. Ca2+-ATPase molecules as a calcium-sensitive membrane-endoskeleton of sarcoplasmic reticulum. Int. J. Mol. Sci. 2021, 22, 2624. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Shirakata, Y.; Sato, D.; Konishi, K.; Watanabe, T.; Nakamura, J. Isolation and characteristics of scallop sarcoplasmic reticulum with calcium transport activity. J. Biochem. 1992, 112, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, J.; Maruyama, Y.; Tajima, G.; Suwa, M.; Sato, C. Elongation and contraction of scallop sarcoplasmic reticulum (SR): ATP stabilizes Ca2+-ATPase crystalline array elongation of SR vesicles. Int. J. Mol. Sci. 2022, 23, 3311. [Google Scholar] [CrossRef] [PubMed]
- Sagara, Y.; Wade, J.B.; Inesi, G. A conformational mechanism for formation of a dead-end complex by the sarcoplasmic reticulum ATPase with thapsigargin. J. Biol. Chem. 1992, 267, 1286–1292. [Google Scholar] [CrossRef]
- Sanger, J.W.; Sanger, J.M. Sarcoplasmic reticulum in the adductor muscle of a Bermuda scallop: Comparison of smooth versus cross-striated portions. Biol. Bull. 1985, 168, 447–460. [Google Scholar] [CrossRef]
- Loesser, K.E.; Castellani, L.; Franzini-Armstrong, C. Dispositions of junctional feet in muscles of invertebrates. J. Muscle Res. Cell Motil. 1992, 13, 161–173. [Google Scholar] [CrossRef]
- Quinn, K.E.; Castellani, L.; Ondrias, K.; Ehrlich, B.E. Characterization of the ryanodine receptor/channel of invertebrate muscle. Am. J. Physiol. 1998, 274, R494–R502. [Google Scholar] [CrossRef]
- Franzini-Armstrong, C. Studies of the triad: I. Structure of the junction in frog twitch fibers. J. Cell Biol. 1970, 47, 488–499. [Google Scholar] [CrossRef]
- Castellani, L.; Hardwicke, P.M.D.; Franzini-Armstrong, C. Effect of Ca2+ on the dimeric structure of scallop sarcoplasmic reticulum. J. Cell Biol. 1989, 108, 511–520. [Google Scholar] [CrossRef]
- Castellani, L.; Hardwicke, P.M.D. Crystalline structure of sarcoplasmic reticulum from scallop. J. Cell Biol. 1983, 97, 557–561. [Google Scholar] [CrossRef] [Green Version]
- Kalabokis, V.N.; Bozzola, J.J.; Castellani, L.; Hardwicke, P.M.D. A possible role for the dimer ribbon state of scallop sarcoplasmic reticulum. J. Biol. Chem. 1991, 266, 22044–22050. [Google Scholar] [CrossRef]
- Jensen, A.-M.L.; Sørensen, T.L.-M.; Olesen, C.; Møller, J.V.; Nissen, P. Modulatory and catalytic modes of ATP binding by the calcium pump. EMBO J. 2006, 25, 2305–2314. [Google Scholar] [CrossRef]
- Patel, A.; Malinovska, L.; Saha, S.; Wang, J.; Alberti, S.; Krishnan, Y.; Hyman, A.A. ATP as a biological hydrotrope. Science 2017, 356, 753–756. [Google Scholar] [CrossRef]
- Komenami, T.; Yoshimura, A.; Matsuno, Y.; Sato, M.; Sato, C. Network of Palladium-Based Nanorings Synthesized by Liquid-Phase Reduction Using DMSO-H2O: In Situ Monitoring of Structure Formation and Drying Deformation by ASEM. Int. J. Mol. Sci. 2020, 21, 3271. [Google Scholar] [CrossRef]
- Nishiyama, H.; Suga, M.; Ogura, T.; Maruyama, Y.; Koizumi, M.; Mio, K.; Kitamura, S.; Sato, C. Atmospheric Scanning Electron Microscope Observes Cells and Tissues in Open Medium Through Silicon Nitride Film. J. Struct. Biol. 2010, 169, 438–449. [Google Scholar] [CrossRef]
- Sato, K.; Naya, M.; Hatano, Y.; Kondo, Y.; Sato, M.; Narita, Y.; Nagano, K.; Naito, M.; Nakayama, K.; Sato, C. Colony spreading of the gliding bacterium Flavobacterium johnsoniae in the absence of the motility adhesin SprB. Sci. Rep. 2021, 11, 697. [Google Scholar] [CrossRef]
- Sato, K.; Naya, M.; Hatano, Y.; Kondo, Y.; Sato, M.; Narita, Y.; Nagano, K.; Naito, M.; Sato, C. Biofilm spreading by the adhesin-dependent gliding motility of Flavobacterium johnsoniae. 1. Internal structure of the biofilm. Int. J. Mol. Sci. 2021, 22, 1894. [Google Scholar] [CrossRef]
- Naya, M.; Sato, C. Pyrene Excimer-Based Fluorescent Labeling of Neighboring Cysteines by Protein Dynamics: ASEM-Induced Thiol-Ene Click Reaction for High Spatial Resolution CLEM. Int. J. Mol. Sci. 2020, 21, 7550. [Google Scholar] [CrossRef]
- Lea, T.J.; Fenton, M.J.; Potter, J.D.; Ashley, C.C. Rapid activation by photolysis of nitr-5 in skinned fibers of the striated adductor muscle from the scallop. Biochim. Biophys. Acta 1990, 1034, 186–194. [Google Scholar] [CrossRef]
- Dux, L.; Taylor, K.A.; Ting-Beall, H.P.; Martonosi, A. Crystallization of the Ca2+-ATPase of sarcoplasmic reticulum by calcium and lanthanide ions. J. Biol. Chem. 1985, 260, 11730–11743. [Google Scholar] [CrossRef]
- Nakamura, J.; Tajima, G. Independence of two conformations of sarcoplasmic reticulum Ca2+-ATPase molecules in hydrolyzing acetyl phosphate. A two-pair model of the ATPase structural unit. J. Biol. Chem. 1997, 272, 19290–19294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, J.; Tajima, G.; FuruKohri, T. Oligomer of Two Types of Conformational Variants of Sarcoplasmic Reticulum Ca2+-ATPase Molecules. In Na/K-ATPase and Related ATPases; Taniguchi, K., Kaya, S., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2000; pp. 373–379. [Google Scholar]
- Toyoshima, C.; Inesi, G. Structural basis of ion pumping by Ca2+-ATPase of the sarcoplasmic reticulum. Annu. Rev. Biochem. 2004, 73, 269–292. [Google Scholar] [CrossRef] [PubMed]
- Olesen, C.; Picard, M.; Winther, A.-M.L.; Gyrup, C.; Morth, J.P.; Oxvig, C.; Møller, J.V.; Nissen, P. The structural basis of calcium transport by the calcium pump. Nature 2007, 450, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Rios, E.; Ma, J.; Gonzales, A. The mechanical hypothesis of excitation-contraction (EC) coupling in skeletal muscle. J. Musc. Res. Cell Motil. 1991, 12, 127–135. [Google Scholar] [CrossRef]
- Ferguson, D.G.; Schwartz, H.W.; Franzini-Armstrong, C. Subunit structure of junctional feet in triads of skeletal muscle: A freeze-drying, rotary-shadowing study. J. Cell Biol. 1984, 99, 1735–1742. [Google Scholar] [CrossRef] [Green Version]
- Wagenknecht, T.; Hsieh, C.-E.; Rath, B.; Fleischer, S.; Marko, M. Electron Tomography of Frozen-Hydrated Isolated Triad Junctions. Biophys. J. 2002, 83, 2491–2501. [Google Scholar] [CrossRef] [Green Version]
- Wagenknecht, T.; Hsieh, C.; Marko, M. Skeletal muscle triad junction ultrastructure by focused-ion-beam milling of muscle and cryo-electron tomography. Eur. J. Transl. Myol. Basic Appl. Myol. 2015, 25, 49–56. [Google Scholar] [CrossRef]
- Asghari, P.; Scriven, D.R.L.; Ng, M.; Panwar, P.; Chou, K.C.C.; van Petegem, F.; Moore, E.D.W. Cardiac ryanodine receptor distribution is dynamic and changed by auxiliary proteins and post-translational modification. eLife 2020, 9, e51602. [Google Scholar] [CrossRef]
- Merton, P.A. Problems of muscle fatigue. Brit. Med. Bull. 1956, 12, 219–221. [Google Scholar] [CrossRef]
- Peachey, L.D. The sarcoplasmic reticulum and transverse tubules of the frog’s sartorius. J. Cell Biol. 1965, 25, 209–231. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.R.; Palade, G.E. Studies on the endoplasmic reticulum. III. Its form and distribution in striated muscle cells. J. Biophys. Biochem. Cytol. 1957, 3, 269–300. [Google Scholar] [CrossRef] [Green Version]
- Ogata, T.; Yamasaki, Y. Scanning electron-microscopic studied on the three-dimensional structure of sarcoplasmic reticulum in the mammalian red, white and intermediate muscle fibers. Cell Tissue Res. 1985, 242, 461–467. [Google Scholar] [CrossRef]
- Song, C.; Satoh, T.; Sekiguchi, T.; Kato, K.; Murata, K. Structural fluctuations of the human proteasome α7 homo-tetradecamer double ring imply the proteasomal α-ring assembly mechanism. Int. J. Mol. Sci. 2021, 22, 4519. [Google Scholar] [CrossRef]
- Sato, C.; Ueno, Y.; Asai, K.; Takahashi, K.; Sato, M.; Engel, A.; Fujiyoshi, Y. The voltage-sensitive sodium channel is a bell-shaped molecule with several cavities. Nature 2001, 409, 1047–1051. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calcium Concentration (µM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0.002 | 0.03 | 0.12 | 0.6 | 1.4 | 18 | 59 | |||
| elongated vesicles | 32 (5.0%) | 45 (9.0%) | 68 (16.2%) | 48 (11.4%) | 30 (3.8%) | 8 (1.3%) | 5 (1.0%) | ||
| tightly elongated | 11 (1.7) | 38 (7.6) | 41 (9.8) | 28 (6.7) | 26 (3.3) | 3 (0.5) | 2 (0.4) | ||
| with crystal-array | 8 (1.2) | 30 (6.0) | 27 (6.4) | 20 (4.8) | 19 (2.4) | 1 (0.2) | 0 | ||
| without crystal-array | 3 (0.5) | 8 (1.6) | 14 (3.3) | 8 (1.9) | 7 (0.9) | 2 (0.3) | 2 (0.4) | ||
| crookedly elongated | 21 (3.3) | 7 (1.4) | 27 (6.4) | 20 (4.8) | 4 (0.5) | 5 (0.8) | 3 (0.6) | ||
| round vesicles | 612 (95.0) | 456 (91.0) | 352 (83.8) | 373 (88.6) | 767 (96.2) | 618 (98.7) | 515 (99.0) | ||
| total | 644 | 501 | 420 | 421 | 797 | 626 | 520 | ||
| A: [Ca2+] Jump-Up | Control 0.003 → 0.003 | 0.003 → 1.1 µM | 0.003 → 9.8 µM | ||
|---|---|---|---|---|---|
| elongated vesicles | 20 (9.8%) | 13 (9.0%) | 11 (1.9%) | ||
| tightly elongated | 20 (9.8) | 13 (9.0) | 7 (1.2) | ||
| with crystal-array | 6 (2.9) | 5 (3.4) | 0 | ||
| without crystal-array | 14 (6.9) | 8 (5.5) | 7 (1.2) | ||
| crookedly elongated | 0 | 0 | 4 (0.7) | ||
| with crystal-array | 0 | 0 | 0 | ||
| without crystal-array | 0 | 0 | 4 (0.7) | ||
| round vesicles | 184 (90.2) | 132 (91.0) | 550 (98.0) | ||
| with crystal-array | 2 (1.0) | 2 (1.4) | 0 | ||
| without crystal-array | 182 (89.2) | 130 (89.7) | 550 (98.0) | ||
| total | 204 (100) | 145 (100) | 561 (100) | ||
| B: [Ca2+] Jump-Down | Control 16 → 15 µM | 16 → 0.003 µM | |||
| elongated vesicles | 5 (2.1%) | 20 (14.2%) | |||
| with crystal-array | 0 | 3 (2.1) | |||
| without crystal-array | 5 (2.1) | 17 (12.1) | |||
| round vesicles | 233 (97.9) | 121 (85.8) | |||
| with crystal-array | 0 | 1 (0.7) | |||
| without crystal-array | 233 (97.9) | 120 (85.1) | |||
| total | 238 (100) | 141 (100) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, J.; Maruyama, Y.; Tajima, G.; Hayakawa, S.; Suwa, M.; Sato, C. Ca2+ Dependent Formation/Collapse of Cylindrical Ca2+-ATPase Crystals in Scallop Sarcoplasmic Reticulum (SR) Vesicles: A Possible Dynamic Role of SR in Regulation of Muscle Contraction. Int. J. Mol. Sci. 2023, 24, 7080. https://doi.org/10.3390/ijms24087080
Nakamura J, Maruyama Y, Tajima G, Hayakawa S, Suwa M, Sato C. Ca2+ Dependent Formation/Collapse of Cylindrical Ca2+-ATPase Crystals in Scallop Sarcoplasmic Reticulum (SR) Vesicles: A Possible Dynamic Role of SR in Regulation of Muscle Contraction. International Journal of Molecular Sciences. 2023; 24(8):7080. https://doi.org/10.3390/ijms24087080
Chicago/Turabian StyleNakamura, Jun, Yuusuke Maruyama, Genichi Tajima, Satoshi Hayakawa, Makiko Suwa, and Chikara Sato. 2023. "Ca2+ Dependent Formation/Collapse of Cylindrical Ca2+-ATPase Crystals in Scallop Sarcoplasmic Reticulum (SR) Vesicles: A Possible Dynamic Role of SR in Regulation of Muscle Contraction" International Journal of Molecular Sciences 24, no. 8: 7080. https://doi.org/10.3390/ijms24087080