
Photobiomodulation Therapy and the Glymphatic System: Promising Applications for Augmenting the Brain Lymphatic Drainage System



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
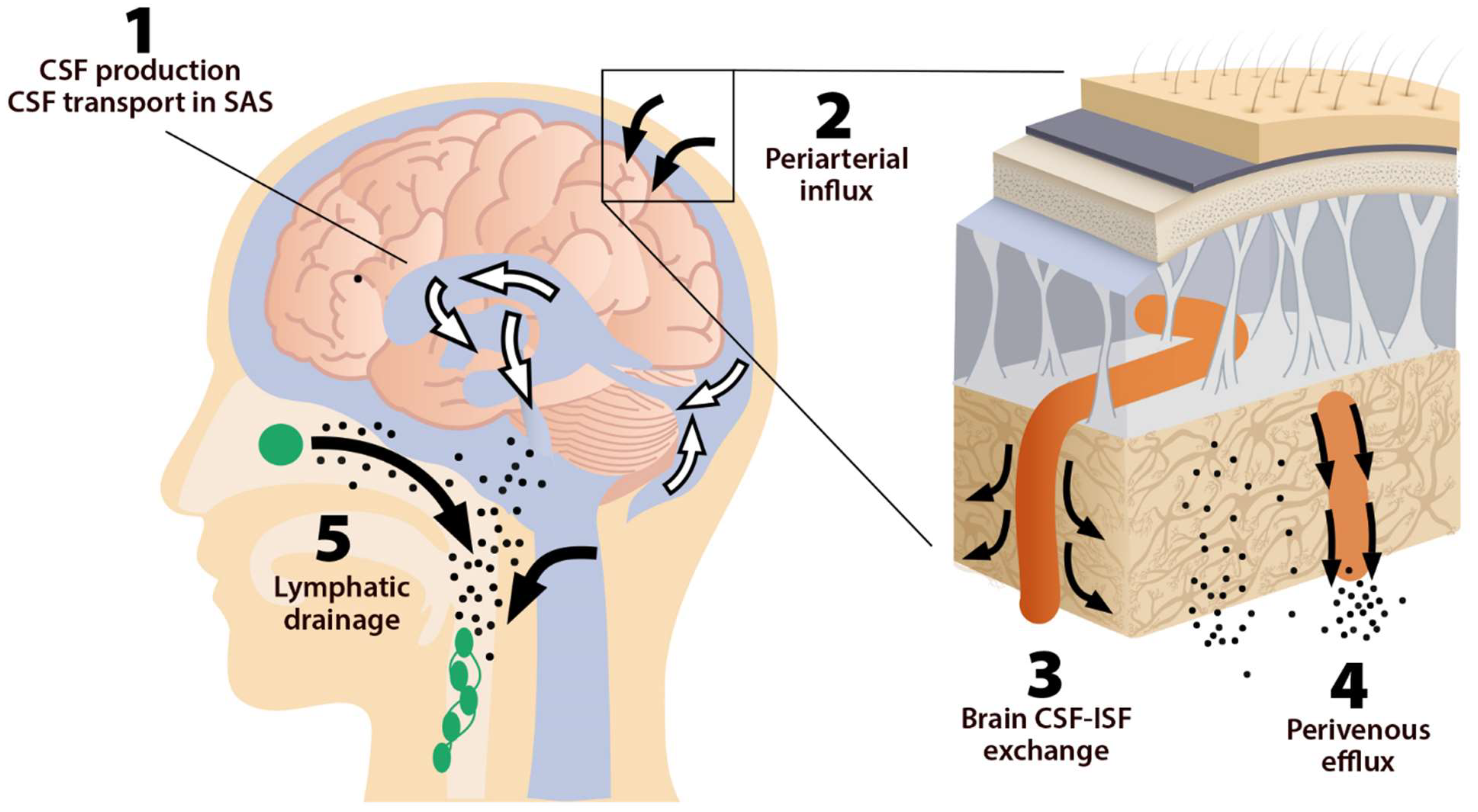
2. Brain Glymphatic Drainage System
2.1. The System, Its Components, and Pathways
2.2. MLVs, Olfactory/Cervical Lymphatic Drainage Route, and Their Association with CSF Circulation
2.3. Sleep and Clearance of the Brain
3. PBM Therapy
3.1. Evidence on Potential Effects of PBM on the Brain Drainage System
3.2. PBM and Nitric Oxide
3.3. PBM and Neuroprotection
3.4. Intranasal and Systemic PBM Therapies and Their Effects on the Brain Drainage System
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamblin, M.R. Mechanisms and Mitochondrial Redox Signaling in Photobiomodulation. Photochem. Photobiol. 2018, 94, 199–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Freitas, L.F.; Hamblin, M.R. Proposed mechanisms of photobiomodulation or low-level light therapy. IEEE J. Sel. Top Quan-Tum Electron 2016, 22, 348–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Lima, F. Neuroprotection and Neurocognitive Augmentation by Photobiomodulation. In Contemporary Clinical Neuroscience; Springer: Singapore, 2021; pp. 165–207. [Google Scholar]
- Salehpour, F.; Mahmoudi, J.; Kamari, F.; Sadigh-Eteghad, S.; Rasta, S.H.; Hamblin, M.R. Brain Photobiomodulation Therapy: A Narrative Review. Mol. Neurobiol. 2018, 55, 6601–6636. [Google Scholar] [CrossRef] [PubMed]
- Cassano, P.; Petrie, S.R.; Hamblin, M.R.; Henderson, T.A.; Iosifescu, D.V. Review of transcranial photobiomodulation for major depressive disorder: Targeting brain metabolism, inflammation, oxidative stress, and neurogenesis. Neurophotonics 2016, 3, 031404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamblin, M.R. Shining light on the head: Photobiomodulation for brain disorders. BBA Clin. 2016, 6, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldieraro, M.A.; Cassano, P. Transcranial and systemic photobiomodulation for major depressive disorder: A systematic review of efficacy, tolerability and biological mechanisms. J. Affect. Disord. 2019, 243, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R.; Huang, Y.-Y. Photobiomodulation in the Brain: Low-Level Laser (Light) Therapy in Neurology and Neuro-science. Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Da Mesquita, S.; Louveau, A.; Vaccari, A.; Smirnov, I.; Cornelison, R.C.; Kingsmore, K.M.; Contarino, C.; Onengut-Gumuscu, S.; Farber, E.; Raper, D.; et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature 2018, 560, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Postnov, D.; Kurths, J. Blood–Brain Barrier, Lymphatic Clearance, and Recovery: Ariadne’s Thread in Labyrinths of Hypotheses. Int. J. Mol. Sci. 2018, 19, 3818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louveau, A.; Smirnov, I.; Keyes, T.J.; Eccles, J.D.; Rouhani, S.J.; Peske, J.D.; Derecki, N.C.; Castle, D.; Mandell, J.W.; Lee, K.S.; et al. Structural and functional features of central nervous system lymphatic vessels. Nature 2015, 523, 337–341. [Google Scholar] [CrossRef]
- Dunn, G.P.; Okada, H. Principles of immunology and its nuances in the central nervous system: Figure 1. Neuro-Oncology 2015, 17, vii3–vii8. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.-L.; Wang, L.-H.; Yang, T.; Sun, J.-Y.; Mao, L.-L.; Yang, M.-F.; Yuan, H.; Colvin, R.A.; Yang, X.-Y. Lymphatic drainage system of the brain: A novel target for intervention of neurological diseases. Prog. Neurobiol. 2018, 163–164, 118–143. [Google Scholar] [CrossRef] [PubMed]
- Zou, W.; Pu, T.; Feng, W.; Lu, M.; Zheng, Y.; Du, R.; Xiao, M.; Hu, G. Blocking meningeal lymphatic drainage aggravates Parkinson’s disease-like pathology in mice overexpressing mutated α-synuclein. Transl. Neurodegener. 2019, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, E.; Navolokin, N.; Shirokov, A.; Khlebtsov, B.; Dubrovsky, A.; Saranceva, E.; Abdurashitov, A.; Khorovodov, A.; Terskov, A.; Mamedova, A.; et al. Pilot study of transcranial photobiomodulation of lymphatic clearance of beta-amyloid from the mouse brain: Breakthrough strategies for non-pharmacologic therapy of Alzheimer’s disease. Biomed. Opt. Express 2019, 10, 4003–4017. [Google Scholar] [CrossRef] [PubMed]
- Zinchenko, E.; Klimova, M.; Mamedova, A.; Agranovich, I.; Blokhina, I.; Antonova, T.; Terskov, A.; Shirokov, A.; Navolokin, N.; Morgun, A.; et al. Photostimulation of Extravasation of Beta-Amyloid through the Model of Blood-Brain Barrier. Electronics 2020, 9, 1056. [Google Scholar] [CrossRef]
- Li, D.; Liu, S.; Yu, T.; Liu, Z.; Sun, S.; Bragin, D.; Navolokin, N.; Kurths, J.; Glushkovskaya-Semyachkina, O.; Zhu, D. Photostimulation of lymphatic clearance of red blood cells from the mouse brain after intraventricular hemor-rhage. bioRxiv. [CrossRef]
- Weller, R.O.; Djuanda, E.; Yow, H.-Y.; Carare, R.O. Lymphatic drainage of the brain and the pathophysiology of neurological disease. Acta Neuropathol. 2009, 117, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Matsumae, M.; Sato, O.; Hirayama, A.; Hayashi, N.; Takizawa, K.; Atsumi, H.; Sorimachi, T. Research into the physiology of cerebrospinal fluid reaches a new horizon: Intimate exchange between cerebrospinal fluid and interstitial fluid may contrib-ute to maintenance of homeostasis in the central nervous system. Neurol. Med. Chir. 2016, 56, 416–441. [Google Scholar] [CrossRef] [Green Version]
- Iliff, J.J.; Wang, M.; Liao, Y.; Plogg, B.A.; Peng, W.; Gundersen, G.A.; Benveniste, H.; Vates, G.E.; Deane, R.; Goldman, S.A.; et al. A Paravascular Pathway Facilitates CSF Flow Through the Brain Parenchyma and the Clearance of Interstitial Solutes, Including Amyloid β. Sci. Transl. Med. 2012, 4, 147ra111. [Google Scholar] [CrossRef] [Green Version]
- Benveniste, H.; Liu, X.; Koundal, S.; Sanggaard, S.; Lee, H.; Wardlaw, J. The Glymphatic System and Waste Clearance with Brain Aging: A Review. Gerontology 2018, 65, 106–119. [Google Scholar] [CrossRef]
- Bakker, E.; Bacskai, B.J.; Arbel-Ornath, M.; Aldea, R.; Bedussi, B.; Morris, A.; Weller, R.O.; Carare, R.O. Lymphatic Clearance of the Brain: Perivascular, Paravascular and Significance for Neurodegenerative Diseases. Cell. Mol. Neurobiol. 2016, 36, 181–194. [Google Scholar] [CrossRef] [Green Version]
- Engelhardt, B.; Carare, R.O.; Bechmann, I.; Flügel, A.; Laman, J.D.; Weller, R.O. Vascular, glial, and lymphatic immune gateways of the central nervous system. Acta Neuropathol. 2016, 132, 317–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carare, R.O.; Bernardes-Silva, M.; Newman, T.A.; Page, A.M.; Nicoll, J.A.R.; Perry, V.H.; Weller, R.O. Solutes, but not cells, drain from the brain parenchyma along basement membranes of capillaries and arteries: Significance for cerebral amyloid angiopathy and neuroimmunology. Neuropathol. Appl. Neurobiol. 2008, 34, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; Olveda, G.; Thomas, J.H.; Nedergaard, M.; Kelley, D.H. Flow of cerebrospinal fluid is driven by arterial pulsations and is reduced in hypertension. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.; Silva, J.; Ferreira, R.; Trigo, D. Glymphatic system, AQP4, and their implications in Alzheimer’s disease. Neurol. Res. Pract. 2021, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Shen, B.; Shen, W.-Q.; Chen, H.; Zheng, Y.-F.; Fei, J.-J. Dysfunction of the Glymphatic System Might Be Related to Iron Deposition in the Normal Aging Brain. Front. Aging Neurosci. 2020, 12, 445. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, M.K.; Mestre, H.; Nedergaard, M. The glymphatic pathway in neurological disorders. Lancet Neurol. 2018, 17, 1016–1024. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, L.; Xu, H.; Wang, Y.; Liang, Q. The lymphatic drainage system of the CNS plays a role in lymphatic drain-age, immunity, and neuroinflammation in stroke. J. Leukoc. Biol. 2021, 110, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sawyer, I.A.; Sung, M.-H.; Sturgill, D.; Shevtsov, S.P.; Pegoraro, G.; Hakim, O.; Baek, S.; Hager, G.L.; Dundr, M. Cajal bodies are linked to genome conformation. Nat. Commun. 2016, 7, 10966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thrane, V.R.; Thrane, A.S.; Plog, B.A.; Thiyagarajan, M.; Iliff, J.J.; Deane, R.; Nagelhus, E.A.; Nedergaard, M. Paravascular microcirculation facilitates rapid lipid transport and astrocyte signaling in the brain. Sci. Rep. 2013, 3, srep02582. [Google Scholar] [CrossRef]
- Achariyar, T.M.; Li, B.; Peng, W.; Verghese, P.B.; Shi, Y.; McConnell, E.; Benraiss, A.; Kasper, T.; Song, W.; Takano, T.; et al. Glymphatic distribution of CSF-derived apoE into brain is isoform specific and suppressed during sleep deprivation. Mol. Neurodegener. 2016, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Kylkilahti, T.M.; Berends, E.; Ramos, M.; Shanbhag, N.C.; Töger, J.; Bloch, K.M.; Lundgaard, I. Achieving brain clearance and preventing neurodegenerative diseases—A glymphatic perspective. Br. J. Pharmacol. 2021, 41, 2137–2149. [Google Scholar] [CrossRef] [PubMed]
- Baranello, R.J.; Bharani, K.L.; Padmaraju, V.; Chopra, N.; Lahiri, D.K.; Greig, N.H.; Pappolla, M.A.; Sambamurti, K. Amy-loid-beta protein clearance and degradation (ABCD) pathways and their role in Alzheimer’s disease. Curr. Alzheimer Res. 2015, 12, 32–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, A.; Iadecola, C. Impaired Aβ clearance: A potential link between atherosclerosis and Alzheimer’s disease. Front. Aging Neurosci. 2015, 7, 115. [Google Scholar] [CrossRef] [PubMed]
- Nedergaard, M.; Goldman, S.A. Glymphatic failure as a final common pathway to dementia. Science 2020, 370, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.C.; Wyler, S.C.; Wan, R.; Castorena, C.M.; Ahmed, N.; Mathew, D.; Lee, S.; Liu, C.; Elmquist, J.K. The atypical anti-psychotic olanzapine causes weight gain by targeting serotonin receptor 2C. J. Clin. Investig. 2017, 127, 3402–3406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, J.H.; Cho, H.; Kim, J.-H.; Kim, S.H.; Ham, J.-S.; Park, I.; Suh, S.H.; Hong, S.P.; Song, J.-H.; Hong, Y.-K.; et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Louveau, A.; Herz, J.; Alme, M.N.; Salvador, A.F.; Dong, M.Q.; Viar, K.E.; Herod, S.G.; Knopp, J.; Setliff, J.C.; Lupi, A.; et al. CNS lymphatic drainage and neuroinflammation are regulated by meningeal lymphatic vasculature. Nat. Neurosci. 2018, 21, 1380–1391. [Google Scholar] [CrossRef] [PubMed]
- King, H.H. Lymphatic Vessels Found in the Brain—Osteopathic Considerations. J. Am. Osteopat. Assoc. 2015, 115, 627. [Google Scholar] [CrossRef] [Green Version]
- Dupont, G.; Iwanaga, J.; Yilmaz, E.; Tubbs, R.S. Connections between amyloid beta and the meningeal lymphatics as a possi-ble route for clearance and therapeutics. Lymphat. Res. Biol. 2020, 18, 2–6. [Google Scholar]
- Wang, X.; Tian, H.; Liu, H.; Liang, D.; Qin, C.; Zhu, Q.; Wang, X. Impaired Meningeal Lymphatic Flow in NMOSD Patients with Acute Attack. Front. Immunol. 2021, 12, 2239. [Google Scholar] [CrossRef]
- Ding, X.-B.; Wang, X.-X.; Xia, D.-H.; Liu, H.; Tian, H.-Y.; Fu, Y.; Chen, Y.-K.; Qin, C.; Wang, J.-Q.; Xiang, Z.; et al. Impaired meningeal lymphatic drainage in patients with idiopathic Parkinson’s disease. Nat. Med. 2021, 27, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Ghandili, M.; Munakomi, S. Neuroanatomy, Putamen. StatPearls [Internet]. 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK542170/ (accessed on 25 December 2021).
- Tripathi, R. Tracing the bulk outflow route of cerebrospinal fluid by transmission and scanning electron microscopy. Brain Res. 1974, 80, 503–506. [Google Scholar] [CrossRef]
- Kida, S.; Pantazis, A.; Weller, R.O. CSF drains directly from the subarachnoid space into nasal lymphatics in the rat. Anatomy, histology and immunological significance. Neuropathol. Appl. Neurobiol. 1993, 19, 480–488. [Google Scholar] [CrossRef] [PubMed]
- De Leon, M.J.; Li, Y.; Okamura, N.; Tsui, W.H.; Saint-Louis, L.A.; Glodzik, L.; Osorio, R.; Fortea, J.; Butler, T.; Pirraglia, E.; et al. Cerebrospinal Fluid Clearance in Alzheimer Disease Measured with Dynamic PET. J. Nucl. Med. 2017, 58, 1471–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Ries, M.; Decker, Y.; Müller, A.; Riner, C.; Bücker, A.; Fassbender, K.; Detmar, M.; Proulx, S.T. Rapid lymphatic efflux limits cerebrospinal fluid flow to the brain. Acta Neuropathol. 2019, 137, 151–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norwood, J.N.; Zhang, Q.; Card, D.; Craine, A.; Ryan, T.M.; Drew, P.J. Anatomical basis and physiological role of cerebro-spinal fluid transport through the murine cribriform plate. elife 2019, 8, e44278. [Google Scholar] [CrossRef] [PubMed]
- Wostyn, P. COVID-19 and chronic fatigue syndrome: Is the worst yet to come? Med. Hypotheses 2021, 146, 110469. [Google Scholar] [CrossRef] [PubMed]
- Rash, J.E.; Davidson, K.G.V.; Kamasawa, N.; Yasumura, T.; Kamasawa, M.; Zhang, C.; Michaels, R.; Restrepo, D.; Ottersen, O.P.; Olson, C.O.; et al. Ultrastructural localization of connexins (Cx36, Cx43, Cx45), glutamate receptors and aquaporin-4 in rodent olfactory mucosa, olfactory nerve and olfactory bulb. J. Neurocytol. 2005, 34, 307–341. [Google Scholar] [CrossRef] [Green Version]
- Brady, M.; Rahman, A.; Combs, A.; Venkatraman, C.; Kasper, R.T.; McQuaid, C.; Kwok, W.-C.E.; Wood, R.W.; Deane, R. Cerebrospinal fluid drainage kinetics across the cribriform plate are reduced with aging. Fluids Barriers CNS 2020, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Ineichen, B.V.; Detmar, M.; Proulx, S.T. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauglund, N.L.; Pavan, C.; Nedergaard, M. Cleaning the sleeping brain–the potential restorative function of the glymphatic system. Curr. Opin. Physiol. 2020, 15, 1–6. [Google Scholar] [CrossRef]
- Benveniste, H.; Heerdt, P.M.; Fontes, M.; Rothman, D.L.; Volkow, N.D. Glymphatic System Function in Relation to Anesthesia and Sleep States. Anesthesia Analg. 2019, 128, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Fultz, N.E.; Bonmassar, G.; Setsompop, K.; Stickgold, R.A.; Rosen, B.R.; Polimeni, J.R.; Lewis, L.D. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 2019, 366, 628–631. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, O.C.; Van Der Werf, Y.D. The sleeping brain: Harnessing the power of the glymphatic system through lifestyle choic-es. Brain Sci. 2020, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Abdurashitov, A.; Dubrovsky, A.; Klimova, M.; Agranovich, I.; Terskov, A.; Shirokov, A.; Vinnik, V.; Kuzmina, A.; Lezhnev, N.; et al. Photobiomodulation of lymphatic drainage and clearance: Perspective strategy for augmentation of meningeal lymphatic functions. Biomed. Opt. Express 2020, 11, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Holth, J.K.; Fritschi, S.K.; Wang, C.; Pedersen, N.P.; Cirrito, J.R.; Mahan, T.E.; Finn, M.B.; Manis, M.; Geerling, J.C.; Fuller, P.M.; et al. The sleep-wake cycle regulates brain interstitial fluid tau in mice and CSF tau in humans. Science 2019, 363, 880–884. [Google Scholar] [CrossRef] [PubMed]
- Raggi, A.; Ferri, R. Sleep disorders in neurodegenerative diseases. Eur. J. Neurol. 2010, 17, 1326–1338. [Google Scholar] [CrossRef] [PubMed]
- Cuddapah, V.; Zhang, S.L.; Sehgal, A. Regulation of the Blood–Brain Barrier by Circadian Rhythms and Sleep. Trends Neurosci. 2019, 42, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yehuda, S.; Sredni, B.; Carasso, R.L.; Kenigsbuch-Sredni, D. REM sleep deprivation in rats results in inflammation and inter-leukin-17 elevation. J. Interferon Cytokine Res. 2009, 29, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Alvarado, G.; Becerril-Villanueva, E.; De Oca, A.C.-M.; Domínguez-Salazar, E.; Salinas-Jazmín, N.; Pérez-Tapia, S.; Pavon, L.; Velázquez-Moctezuma, J.; Gómez-González, B. The yin/yang of inflammatory status: Blood-brain barrier regula-tion during sleep. Brain Behav. Immun. 2018, 69, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-X.; He, X.; Wu, D.; Zhang, Q.; Yang, C.; Liang, F.-Y.; He, X.-F.; Dai, G.-Y.; Pei, Z.; Lan, Y.; et al. Continuous theta burst stimulation facilitates the clearance efficiency of the glymphatic pathway in a mouse model of sleep deprivation. Neurosci. Lett. 2017, 653, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.R.; Carroll, J.D.; De Freitas, L.F.; Huang, Y.-Y.; Ferraresi, C. Low-Level Light Therapy: Photobiomodulation; SPIE: Bellingham, WA, USA, 2018. [Google Scholar]
- Hamblin, M.R.; Agrawal, T.; de Sousa, M. Handbook of Low-Level Laser Therapy; Jenny Stanford Publishing: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Karu, T.I. Mitochondrial Signaling in Mammalian Cells Activated by Red and Near-IR Radiation. Photochem. Photobiol. 2008, 84, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Karu, T. Photobiology of Low-power Laser Effects. Health Phys. 1989, 56, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Karu, T.; Kolyakov, S. Exact Action Spectra for Cellular Responses Relevant to Phototherapy. Photomed. Laser Surg. 2005, 23, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong-Riley, M.T.; Liang, H.L.; Eells, J.T.; Chance, B.; Henry, M.M.; Buchmann, E.; Kane, M.; Whelan, H.T. Photobiomodula-tion directly benefits primary neurons functionally inactivated by toxins: Role of cytochrome c oxidase. J. Biol. Chem. 2005, 280, 4761–4771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana-Blank, L.; Rodríguez-Santana, E.; Santana-Rodriguez, K. Theoretic, experimental, clinical bases of the water oscilla-tor hypothesis in near-infrared photobiomodulation. Photomed. Laser Surg. 2010, 28, S41–S52. [Google Scholar] [CrossRef] [PubMed]
- Karu, T.I. Cellular and Molecular Mechanisms of Photobiomodulation (Low-Power Laser Therapy). IEEE J. Sel. Top. Quantum Electron. 2013, 20, 143–148. [Google Scholar] [CrossRef]
- Ganeshan, V.; Skladnev, N.V.; Kim, J.Y.; Mitrofanis, J.; Stone, J.; Johnstone, D.M. Pre-conditioning with remote photobio-modulation modulates the brain transcriptome and protects against MPTP insult in mice. Neuroscience 2019, 400, 85–97. [Google Scholar] [CrossRef]
- George, S.; Hamblin, M.R.; Abrahamse, H. Photobiomodulation-Induced Differentiation of Immortalized Adipose Stem Cells to Neuronal Cells. Lasers Surg. Med. 2020, 52, 1032–1040. [Google Scholar] [CrossRef]
- Chung, H.; Dai, T.; Sharma, S.K.; Huang, Y.-Y.; Carroll, J.D.; Hamblin, M.R. The nuts and bolts of low-level laser (light) ther-apy. Ann. Biomed. Eng. 2012, 40, 516–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezani, F.; Neshasteh-Riz, A.; Ghadaksaz, A.; Fazeli, S.M.; Janzadeh, A.; Hamblin, M.R. Mechanistic aspects of photobi-omodulation therapy in the nervous system. Lasers Med. Sci. 2021, 1–8, Epub ahead of print. [Google Scholar] [CrossRef]
- Salehpour, F.; Gholipour-Khalili, S.; Farajdokht, F.; Kamari, F.; Walski, T.; Hamblin, M.R.; DiDuro, J.O.; Cassano, P. Thera-peutic potential of intranasal photobiomodulation therapy for neurological and neuropsychiatric disorders: A narrative re-view. Rev. Neurosci. 2020, 31, 269–286. [Google Scholar] [CrossRef] [PubMed]
- Caldieraro, M.A.; Sani, G.; Bui, E.; Cassano, P. Long-term near-infrared photobiomodulation for anxious depression compli-cated by Takotsubo cardiomyopathy. J. Clin. Psychopharmacol. 2018, 38, 268–270. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Guo, Y.; Chu, X.; Jia, S.; Zheng, X.; Zhou, C. Effects of low power laser irradiation in nasal cavity on cerebral blood flow perfusion of patients with brain infarction. Chin. J. Rehabil. Med. 2003, 12, wpr-573842. [Google Scholar]
- Hennessy, M.; Hamblin, M.R. Photobiomodulation and the brain: A new paradigm. J. Opt. 2017, 19, 013003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamper, M.; Cassano, P.; Lombard, J. Treatment of Kleine-Levin Syndrome with Intranasal Photobiomodulation and Meth-ylene Blue. Cureus 2021, 13, e18596. [Google Scholar] [CrossRef] [PubMed]
- Salehpour, F.; Hamblin, M.R.; DiDuro, J.O. Rapid Reversal of Cognitive Decline, Olfactory Dysfunction, and Quality of Life Using Multi-Modality Photobiomodulation Therapy: Case Report. Photobiomodul. Photomed. Laser Surg. 2019, 37, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Klimova, M.; Iskra, T.; Bragin, D.; Abdurashitov, A.; Dubrovsky, A.; Khorovodov, A.; Terskov, A.; Blokhina, I.; Lezhnev, N.; et al. Transcranial Photobiomodulation of Clearance of Beta-Amyloid from the Mouse Brain: Effects on the Meningeal Lymphatic Drainage and Blood Oxygen Saturation of the Brain. Adv. Exp. Med. Biol. 2021, 1269, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Fedosov, I.; Shirokov, A.; Vodovozova, E.; Alekseeva, A.; Khorovodov, A.; Blokhina, I.; Terskov, A.; Mamedova, A.; Klimova, M.; et al. Photomodulation of lymphatic delivery of liposomes to the brain bypassing the blood-brain barrier: New perspectives for glioma therapy. Nanophotonics 2021, 10, 3215–3227. [Google Scholar] [CrossRef]
- Li, D.; Liu, S.; Yu, T.; Liu, Z.; Sun, S.; Bragin, D.; Shirokov, A.; Navolokin, N.; Bragina, O.; Kurths, J.; et al. Noninvasive technology of photostimulation of lymphatic clearance of red blood cells from the mouse brain after intraventricular hemorrhage. Res. Sq. 2021, 1–24. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Abdurashitov, A.; Klimova, M.; Dubrovsky, A.; Shirokov, A.; Fomin, A.; Terskov, A.; Agranovich, I.; Mamedova, A.; Khorovodov, A.; et al. Photostimulation of cerebral and peripheral lymphatic functions. Transl. Biophotonics 2020, 2, 201900036. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Postnov, D.; Lavrova, A.; Fedosov, I.; Borisova, E.; Nikolenko, V.; Penzel, T.; Kurths, J.; Tuchin, V. Biophotonic Strategies of Measurement and Stimulation of the Cranial and the Extracranial Lymphatic Drainage Function. IEEE J. Sel. Top. Quantum Electron. 2020, 27, 1–13. [Google Scholar] [CrossRef]
- Saucedo, C.L.; Courtois, E.C.; Wade, Z.S.; Kelley, M.N.; Kheradbin, N.; Barrett, D.W.; Gonzalez-Lima, F. Transcranial laser stimulation: Mitochondrial and cerebrovascular effects in younger and older healthy adults. Brain Stimul. 2021, 14, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Uozumi, Y.; Nawashiro, H.; Sato, S.; Kawauchi, S.; Shima, K.; Kikuchi, M. Targeted increase in cerebral blood flow by tran-scranial near-infrared laser irradiation. Lasers Surg. Med. 2010, 42, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Borghammer, P.; Cumming, P.; Østergaard, K.; Gjedde, A.; Rodell, A.; Bailey, C.J.; Vafaee, M.S. Cerebral oxygen metabolism in patients with early Parkinson’s disease. J. Neurol. Sci. 2012, 313, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Litscher, G.; Min, L.; Passegger, C.A.; Litscher, D.; Li, M.; Wang, M.; Ghaffari-Tabrizi-Wizsy, N.; Stelzer, I.; Feigl, G.; Gaischek, I.; et al. Transcranial Yellow, Red, and Infrared Laser and LED Stimulation: Changes of Vascular Parameters in a Chick Embryo Model. Integr. Med. Int. 2015, 2, 80–89. [Google Scholar] [CrossRef]
- Colombo, E.; Signore, A.; Aicardi, S.; Zekiy, A.; Utyuzh, A.; Benedicenti, S.; Amaroli, A. Experimental and clinical applica-tions of red and near-infrared photobiomodulation on endothelial dysfunction: A Review. Biomedicines 2021, 9, 274. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, U.H.; Mack, G.L. Low-level laser treatment with near-infrared light increases venous nitric oxide levels acutely: A single-blind, randomized clinical trial of efficacy. Am. J. Phys. Med. Rehabil. 2013, 92, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, D.; Keszler, A.; Lindemer, B.; Krolikowski, J.; Lohr, N.L. Red light stimulates vasodilation through extracellular vesicle trafficking. J. Photochem. Photobiol. B Biol. 2021, 220, 112212. [Google Scholar] [CrossRef]
- Lubart, R.; Eichler, M.; Lavi, R.; Friedman, H.; Shainberg, A. Low-Energy Laser Irradiation Promotes Cellular Redox Activity. Photomed. Laser Surg. 2005, 23, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M. The role of nitric oxide in low level light therapy. In Mechanisms for Low-Light Therapy III; International Society for Optics and Photonics: Bellingham, WA, USA, 2008; Volume 6846, p. 684602. [Google Scholar]
- Scallan, J.P.; Zawieja, S.D.; Castorena-Gonzalez, J.; Davis, M.J. Lymphatic pumping: Mechanics, mechanisms and malfunction. J. Physiol. 2016, 594, 5749–5768. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Liu, L.; Xing, D. Photobiomodulation by low-power laser irradiation attenuates Aβ-induced cell apoptosis through the Akt/GSK3β/β-catenin pathway. Free Radic. Biol. Med. 2012, 53, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; He, Z.; Xing, D. Low-Level Laser Therapy Rescues Dendrite Atrophy via Upregulating BDNF Expression: Implications for Alzheimer’s Disease. J. Neurosci. 2013, 33, 13505–13517. [Google Scholar] [CrossRef] [PubMed]
- Duggett, N.A.; Chazot, P.L. Low-Intensity Light Therapy (1068 nm) Protects CAD Neuroblastoma Cells from β-Amyloid-Mediated Cell Death. Biol. Med. 2014, 1, 2. [Google Scholar]
- Zhang, Z.; Shen, Q.; Wu, X.; Zhang, D.; Xing, D. Activation of PKA/SIRT1 signaling pathway by photobiomodulation thera-py reduces Aβ levels in Alzheimer’s disease models. Aging Cell 2020, 19, e13054. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Liu, Q.; Zhang, F.; Fu, Y.; Zhu, X.; Weng, X.; Wei, X. Microglia modulation with 1070-nm light attenuates Aβ burden and cognitive impairment in Alzheimer’s dis-ease mouse model. Light Sci. Appl. 2021, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.; Mei, Y.; Zhang, Y.; Tong, Z.; Cui, D.; Yang, J.; Wang, A.; Wang, R.; Fei, X.; Ai, L.; et al. New insight into Alzheimer’s disease: Light reverses Aβ-obstructed interstitial fluid flow and ameliorates memory decline in APP/PS1 mice. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2019, 5, 671–684. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.C.-Y.; Cheng, L.; Su, W.-J.; Zhang, Y.-W.; Shi, Y.; Liu, A.-H.; Zhang, L.-L.; Qian, Z.-Y. Randomized, Double-Blind, and Placebo-Controlled Clinic Report of Intranasal Low-Intensity Laser Therapy on Vascular Diseases. Int. J. Photoenergy 2012, 2012, 1–5. [Google Scholar] [CrossRef]
- Gao, X.; Zhi, P.-K.; Wu, X.-J. Low-energy semiconductor laser intranasal irradiation of the blood improves blood coagulation status in normal pregnancy at term. J. South. Med Univ. 2008, 28, 1400–1401. [Google Scholar]
- Elwood, P.; Pickering, J.; Gallacher, J.E.J. Cognitive function and blood rheology: Results from the Caerphilly cohort of older men. Age Ageing 2001, 30, 135–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Zhang, L.; Qin, C. The relationship between hemorheological changes and the anxiety and depression symptoms in schizophrenia. Clin. J. Hemorheol. 2004, 1. [Google Scholar]
- Chakraborty, S.; Davis, M.J.; Muthuchamy, M. Emerging trends in the pathophysiology of lymphatic contractile function. Semin. Cell Dev. Biol. 2015, 38, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnstone, D.M.; Gordon, L.C. Remote photobiomodulation: An emerging strategy for neuroprotection. Neural Regen. Res. 2019, 14, 2086–2087. [Google Scholar] [CrossRef] [PubMed]
- Caldieraro, M.A.; Salehpour, F.; Cassano, P. Transcranial and Systemic Photobiomodulation for the Enhancement of Mito-Chondrial Metabolism in Depression, in Clinical Bioenergetics; Elsevier: Amsterdam, The Netherlands, 2021; pp. 635–651. [Google Scholar] [CrossRef]
- Farfara, D.; Tuby, H.; Trudler, D.; Doron-Mandel, E.; Maltz, L.; Vassar, R.J.; Frenkel, D.; Oron, U. Low-Level Laser Therapy Ameliorates Disease Progression in a Mouse Model of Alzheimer’s Disease. J. Mol. Neurosci. 2015, 55, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Blivet, G.; Meunier, J.; Roman, F.J.; Touchon, J. Neuroprotective effect of a new photobiomodulation technique against Aβ 25–35 peptide–induced toxicity in mice: Novel hypothesis for therapeutic approach of Alzheimer’s disease suggested. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Mitrofanis, J.; Stone, J.; Johnstone, D.M. Remote tissue conditioning is neuroprotective against MPTP insult in mice. IBRO Rep. 2018, 4, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, D.M.; El Massri, N.; Moro, C.; Spana, S.; Wang, X.S.; Torres, N.; Mitrofanis, J. Indirect application of near infrared light induces neuroprotection in a mouse model of parkinsonism–an abscopal neuroprotective effect. Neuroscience 2014, 274, 93–101. [Google Scholar] [CrossRef]
- Tuby, H.; Maltz, L.; Oron, U. Implantation of Low-Level Laser Irradiated Mesenchymal Stem Cells into the Infarcted Rat Heart Is Associated with Reduction in Infarct Size and Enhanced Angiogenesis. Photomed. Laser Surg. 2009, 27, 227–233. [Google Scholar] [CrossRef]
- Tuby, H.; Maltz, L.; Oron, U. Induction of autologous mesenchymal stem cells in the bone marrow by low-level laser therapy has profound beneficial effects on the infarcted rat heart. Lasers Surg. Med. 2011, 43, 401–409. [Google Scholar] [CrossRef]
- Matsushita, T.; Kibayashi, T.; Katayama, T.; Yamashita, Y.; Suzuki, S.; Kawamata, J.; Honmou, O.; Minami, M.; Shimohama, S. Mesenchymal stem cells transmigrate across brain microvascular endothelial cell monolayers through transiently formed inter-endothelial gaps. Neurosci. Lett. 2011, 502, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Salehpour, F.; Khademi, M.; Hamblin, M.R. Photobiomodulation Therapy for Dementia: A Systematic Review of Pre-Clinical and Clinical Studies. J. Alzheimer’s Dis. 2021, 83, 1431–1452. [Google Scholar] [CrossRef] [PubMed]
- Berman, M.H.; Nichols, T.W. Treatment of Neurodegeneration: Integrating Photobiomodulation and Neurofeedback in Alz-heimer’s Dementia and Parkinson’s: A Review. Photobiomodul. Photomed. Laser Surg. 2019, 37, 623–634. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salehpour, F.; Khademi, M.; Bragin, D.E.; DiDuro, J.O. Photobiomodulation Therapy and the Glymphatic System: Promising Applications for Augmenting the Brain Lymphatic Drainage System. Int. J. Mol. Sci. 2022, 23, 2975. https://doi.org/10.3390/ijms23062975
Salehpour F, Khademi M, Bragin DE, DiDuro JO. Photobiomodulation Therapy and the Glymphatic System: Promising Applications for Augmenting the Brain Lymphatic Drainage System. International Journal of Molecular Sciences. 2022; 23(6):2975. https://doi.org/10.3390/ijms23062975
Chicago/Turabian StyleSalehpour, Farzad, Mahsa Khademi, Denis E. Bragin, and Joseph O. DiDuro. 2022. "Photobiomodulation Therapy and the Glymphatic System: Promising Applications for Augmenting the Brain Lymphatic Drainage System" International Journal of Molecular Sciences 23, no. 6: 2975. https://doi.org/10.3390/ijms23062975