
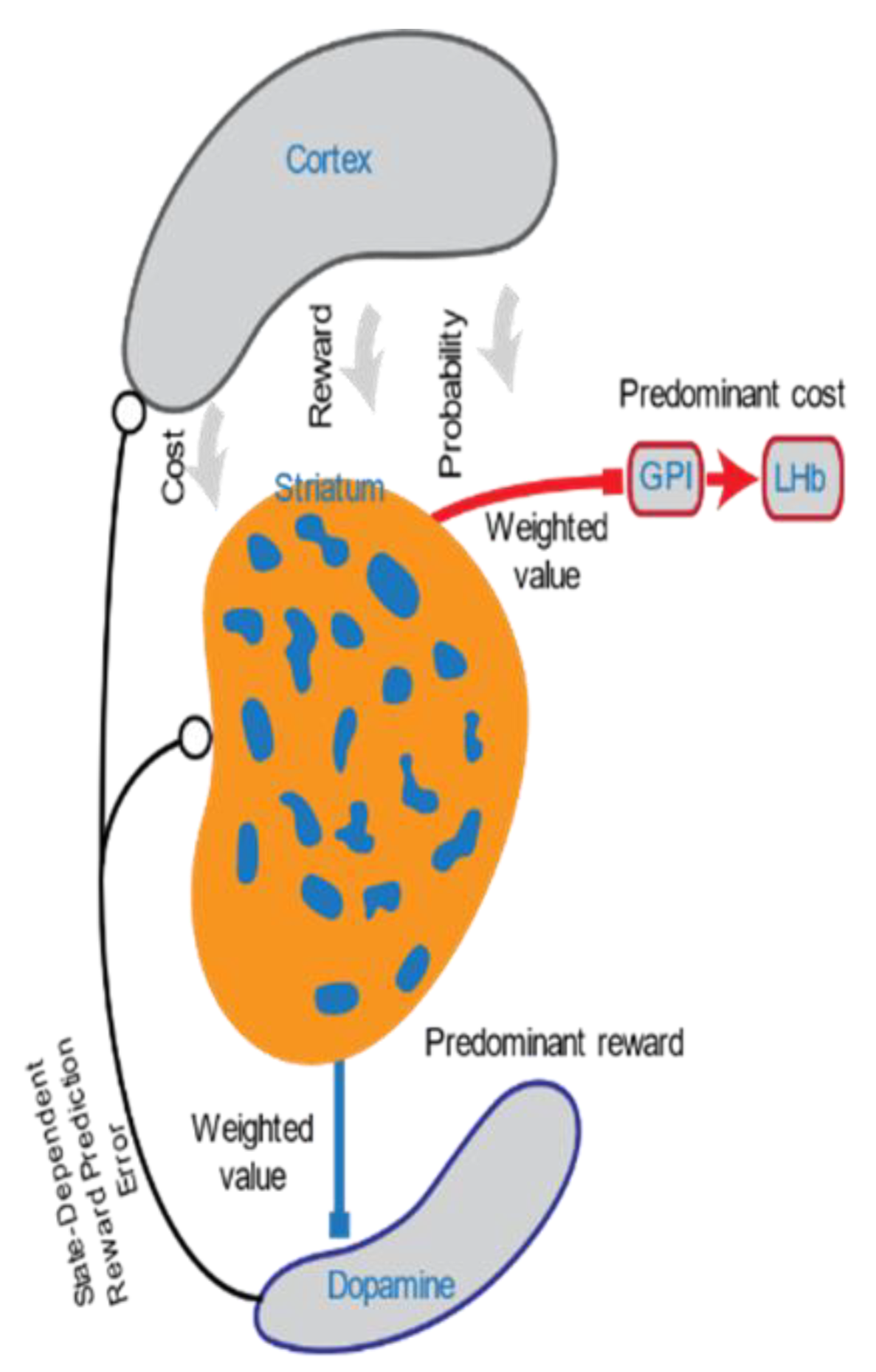
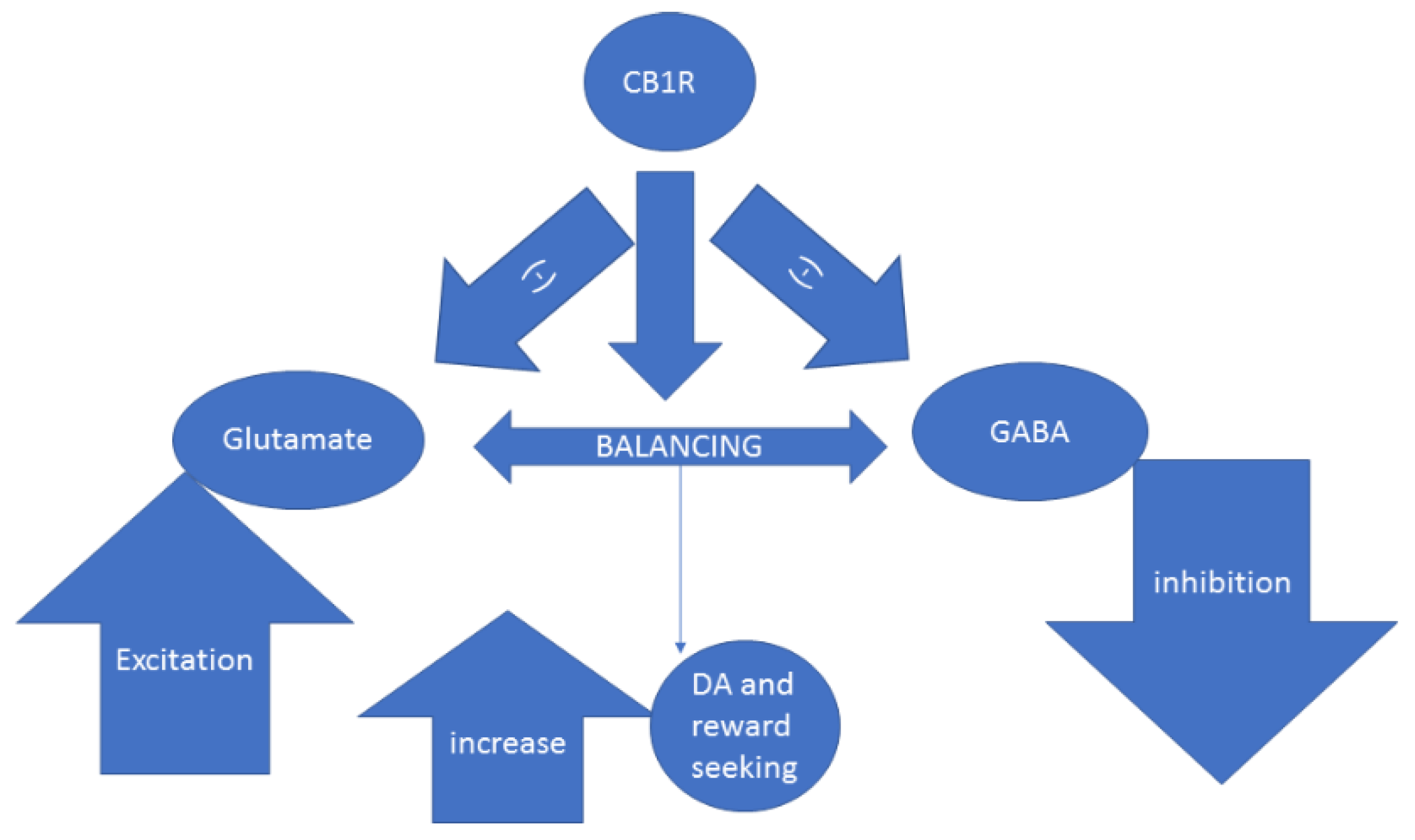
Discovering the Lost Reward: Critical Locations for Endocannabinoid Modulation of the Cortico–Striatal Loop That Are Implicated in Major Depression



{kind=link}
{kind=link}
Abstract
:1. Major Depression: Diagnostic Criteria, Prevalence and Etiology
2. Dopamine and Major Depression
2.1. Location of DA Activity within Subcortical Circuits
2.2. Top-Down Bottom-Up Cortical DA Connectivity with Subcortical Circuits
2.3. The Excitation-Inhibition Imbalance
3. The Reward Circuits and Major Depression
3.1. The Striatum and the Mesolimbic Area
3.2. The Striosomal Compartment of Striatum
4. The Endocannabinoid System and Major Depression
5. The Crosstalk between Dopamine, Striosomes and Endocannabinoid Receptors
6. The CB1 Crosstalk with the Striosomal Activity on the Molecular Level
7. Vulnerability of Dopamine and Endocannabinoid Systems within the Reward Circuits: Implications for Major Depression
8. Insufficiency of Past and Current Pharmacological Treatments for Major Depression: Implications for Novel Treatment with CBD
9. For Further Research
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kessler, R.C.; Bromet, E.J. The Epidemiology of Depression Across Cultures. Annu. Rev. Public Health 2013, 34, 119–138. [Google Scholar] [CrossRef] [Green Version]
- Collins, P.Y.; Patel, V.; Joestl, S.S.; March, D.; Insel, T.R.; Daar, A.S.; Bordin, I.A.; Costello, E.J.; Durkin, M.; Fairburn, C.G.; et al. Grand challenges in global mental health. Nat. Cell Biol. 2011, 475, 27–30. [Google Scholar] [CrossRef] [PubMed]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association Publishing: Washington, DC, USA, 2013. [Google Scholar] [CrossRef]
- Athira, K.V.; Bandopadhyay, S.; Samudrala, P.K.; Naidu, V.; Lahkar, M.; Chakravarty, S. An Overview of the Heterogeneity of Major Depressive Disorder: Current Knowledge and Future Prospective. Curr. Neuropharmacol. 2020, 18, 168–187. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, C.L.; Frank, E.; Kupfer, D.J. Social Zeitgebers and Biological Rhythms. Arch. Gen. Psychiatry 1988, 45, 948–952. [Google Scholar] [CrossRef] [PubMed]
- Visentin, A.P.V.; Colombo, R.; Scotton, E.; Fracasso, D.S.; Da Rosa, A.R.; Branco, C.S.; Salvador, M. Targeting Inflammatory-Mitochondrial Response in Major Depression: Current Evidence and Further Challenges. Oxidative Med. Cell. Longev. 2020, 2020, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Li, J.; Gui, X.; Shi, X.; Bao, Z.; Han, H.; Li, M.D. Updated review of research on the gut microbiota and their relation to depression in animals and human beings. Mol. Psychiatry 2020, 25, 2759–2772. [Google Scholar] [CrossRef]
- Beurel, E.; Toups, M.; Nemeroff, C.B. The Bidirectional Relationship of Depression and Inflammation: Double Trouble. Neuron 2020, 107, 234–256. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, M.; Gu, S.; Wang, F.; Yuan, B. Early Life Stress Induced DNA Methylation of Monoamine Oxidases Leads to Depressive-Like Behavior. Front. Cell Dev. Biol. 2020, 8, 8. [Google Scholar] [CrossRef]
- Coelho, F.M.D.C.; Pinheiro, R.T.; Silva, R.A.; de Ávila Quevedo, L.; Souza, L.D.D.M.; Castelli, R.D.; De Matos, M.B.; Pinheiro, K.A.T. Major Depressive Disorder during Teenage Pregnancy: Socio-demographic, Obstetric and Psychosocial Correlates. Rev. Bras. Psiquiatr. 2013, 35, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Menezes, I.C.; Baes, C.V.W.; Lacchini, R.; Juruena, M.F. Genetic biomarkers for differential diagnosis of major depressive disorder and bipolar disorder: A systematic and critical review. Behav. Brain Res. 2019, 358, 29–38. [Google Scholar] [CrossRef]
- Faye, C.; McGowan, J.C.; Denny, C.A.; David, D.J. Neurobiological Mechanisms of Stress Resilience and Implications for the Aged Population. Curr. Neuropharmacol. 2018, 16, 234–270. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, A.M.; Sullivan, P.F.; Lewis, C.M. Uncovering the Genetic Architecture of Major Depression. Neuron 2019, 102, 91–103. [Google Scholar] [CrossRef]
- Wohlschläger, A.; Karne, H.; Jordan, D.; Lowe, M.J.; Jones, S.E.; Anand, A. Spectral Dynamics of Resting State fMRI Within the Ventral Tegmental Area and Dorsal Raphe Nuclei in Medication-Free Major Depressive Disorder in Young Adults. Front. Psychiatry 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Chen, A.L.C.; Oscar-Berman, M.; Chen, T.J.H.; Lubar, J.; White, N.; Lubar, J.; Bowirrat, A.; Braverman, E.; Schoolfield, J.; et al. Generational Association Studies of Dopaminergic Genes in Reward Deficiency Syndrome (RDS) Subjects: Selecting Appropriate Phenotypes for Reward Dependence Behaviors. Int. J. Environ. Res. Public Health 2011, 8, 4425–4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dremencov, E.; Gispan-Herman, I.; Rosenstein, M.; Mendelman, A.; Overstreet, D.H.; Zohar, J.; Yadid, G. The serotonin–dopamine interaction is critical for fast-onset action of antidepressant treatment: In vivo studies in an animal model of depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2004, 28, 141–147. [Google Scholar] [CrossRef]
- Lavi-Avnon, Y.; Weller, A.; Finberg, J.P.M.; Gispan-Herman, I.; Kinor, N.; Stern, Y.; Schroeder, M.; Gelber, V.; Bergman, S.Y.; Overstreet, D.H.; et al. The reward system and maternal behavior in an animal model of depression: A microdialysis study. Psychopharmacology 2007, 196, 281–291. [Google Scholar] [CrossRef]
- Yadid, G.; Friedman, A. Dynamics of the dopaminergic system as a key component to the understanding of depression. Prog. Brain Res. 2008, 172, 265–286. [Google Scholar] [CrossRef]
- Dremencov, E.; Newman, M.E.; Kinor, N.; Blatman-Jan, G.; Schindler, C.J.; Overstreet, D.H.; Yadid, G. Hyperfunctionality of serotonin-2C receptor-mediated inhibition of accumbal dopamine release in an animal model of depression is reversed by antidepressant treatment. Neuropharmacology 2005, 48, 34–42. [Google Scholar] [CrossRef]
- Zangen, A.; Nakash, R.; Overstreet, D.; Yadid, G. Association between depressive behavior and absence of serotonin-dopamine interaction in the nucleus accumbens. Psychopharmacology 2001, 155, 434–439. [Google Scholar] [CrossRef]
- Dremencov, E.; Weizmann, Y.; Kinor, N.; Gispan-Herman, I.; Yadid, G. Modulation of Dopamine Transmission by 5HT2C and 5HT3 Receptors: A Role in the Antidepressant Response. Curr. Drug Targets 2006, 7, 165–175. [Google Scholar] [CrossRef]
- Schwerdt, H.N.; Amemori, K.; Gibson, D.J.; Stanwicks, L.L.; Yoshida, T.; Bichot, N.P.; DeSimone, R.; Langer, R.; Cima, M.J.; Graybiel, A.M. Dopamine and beta-band oscillations differentially link to striatal value and motor control. Sci. Adv. 2020, 6, eabb9226. [Google Scholar] [CrossRef] [PubMed]
- Bunney, W.E. Norepinephrine in Depressive Reactions. Arch. Gen. Psychiatry 1965, 13, 483–494. [Google Scholar] [CrossRef]
- Schildkraut, J.J.; Gordon, E.K.; Durell, J. Catecholamine metabolism in affective disorders: I. J. Psychiatr. Res. 1965, 3, 213–228. [Google Scholar] [CrossRef]
- Coppen, A. The Biochemistry of Affective Disorders. Br. J. Psychiatry 1967, 113, 1237–1264. [Google Scholar] [CrossRef]
- Belujon, P.; Grace, A.A. Restoring Mood Balance in Depression: Ketamine Reverses Deficit in Dopamine-Dependent Synaptic Plasticity. Biol. Psychiatry 2014, 76, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Tye, K.M.; Deisseroth, K. Optogenetic investigation of neural circuits underlying brain disease in animal models. Nat. Rev. Neurosci. 2012, 13, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, D.; Walsh, J.J.; Friedman, A.K.; Juarez, B.; Ku, S.M.; Koo, J.W.; Ferguson, D.; Tsai, H.-C.; E Pomeranz, L.; Christoffel, D.J.; et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nat. Cell Biol. 2012, 493, 532–536. [Google Scholar] [CrossRef]
- Savitz, J.; Hodgkinson, C.A.; Martin-Soelch, C.; Shen, P.-H.; Szczepanik, J.; Nugent, A.; Herscovitch, P.; Grace, A.A.; Goldman, D.; Drevets, W.C. The Functional DRD3 Ser9Gly Polymorphism (rs6280) Is Pleiotropic, Affecting Reward as Well as Movement. PLoS ONE 2013, 8, e54108. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-H.; Grace, A.A. Amygdala-Ventral Pallidum Pathway Decreases Dopamine Activity After Chronic Mild Stress in Rats. Biol. Psychiatry 2014, 76, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Moreines, J.L.; Owrutsky, Z.L.; A Grace, A. Involvement of Infralimbic Prefrontal Cortex but not Lateral Habenula in Dopamine Attenuation After Chronic Mild Stress. Neuropsychopharmacology 2016, 42, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Friedman, Y.; Dremencov, E.; Yadid, G. VTA Dopamine Neuron Bursting is Altered in an Animal Model of Depression and Corrected by Desipramine. J. Mol. Neurosci. 2008, 34, 201–209. [Google Scholar] [CrossRef]
- Tye, K.M.; Mirzabekov, J.J.; Warden, M.R.; Ferenczi, E.A.; Tsai, H.-C.; Finkelstein, J.; Kim, S.-Y.; Adhikari, A.; Thompson, K.R.; Andalman, A.S.; et al. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nat. Cell Biol. 2012, 493, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.; Frankel, M.; Flaumenhaft, Y.; Merenlender, A.; Pinhasov, A.; Feder, Y.; Taler, M.; Gil-Ad, I.; Abeles, M.; Yadid, G. Programmed Acute Electrical Stimulation of Ventral Tegmental Area Alleviates Depressive-Like Behavior. Neuropsychopharmacology 2008, 34, 1057–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazit, T.; Friedman, A.; Lax, E.; Samuel, M.; Zahut, R.; Katz, M.; Abraham, L.; Tischler, H.; Teicher, M.; Yadid, G. Programmed deep brain stimulation synchronizes VTA gamma band field potential and alleviates depressive-like behavior in rats. Neuropharmacology 2015, 91, 135–141. [Google Scholar] [CrossRef]
- Dunlop, B.W.; Nemeroff, C.B. The Role of Dopamine in the Pathophysiology of Depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef]
- Paret, C.; Kluetsch, R.; Zaehringer, J.; Ruf, M.; Demirakca, T.; Bohus, M.; Ende, G.; Schmahl, C. Alterations of amygdala-prefrontal connectivity with real-time fMRI neurofeedback in BPD patients. Soc. Cogn. Affect. Neurosci. 2016, 11, 952–960. [Google Scholar] [CrossRef] [Green Version]
- Hasler, G.; Fromm, S.; Carlson, P.J.; Luckenbaugh, D.A.; Waldeck, T.; Geraci, M.; Roiser, J.P.; Neumeister, A.; Meyers, N.; Charney, D.S.; et al. Neural Response to Catecholamine Depletion in Unmedicated Subjects With Major Depressive Disorder in Remission and Healthy Subjects. Arch. Gen. Psychiatry 2008, 65, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.; Homma, D.; Gibb, L.G.; Amemori, K.-I.; Rubin, S.J.; Hood, A.S.; Riad, M.H.; Graybiel, A.M. A Corticostriatal Path Targeting Striosomes Controls Decision-Making under Conflict. Cell 2015, 161, 1320–1333. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.; Hueske, E.; Drammis, S.M.; Arana, S.E.T.; Nelson, E.D.; Carter, C.W.; Delcasso, S.; Rodriguez, R.X.; Lutwak, H.; DiMarco, K.S.; et al. Striosomes Mediate Value-Based Learning Vulnerable in Age and a Huntington’s Disease Model. Cell 2020, 183, 918–934.e49. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.; Homma, D.; Bloem, B.; Gibb, L.G.; Amemori, K.-I.; Hu, D.; Delcasso, S.; Truong, T.F.; Yang, J.; Hood, A.S.; et al. Chronic Stress Alters Striosome-Circuit Dynamics, Leading to Aberrant Decision-Making. Cell 2017, 171, 1191–1205.e28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, K.C. The debate over dopamine’s role in reward: The case for incentive salience. Psychopharmacology 2007, 191, 391–431. [Google Scholar] [CrossRef]
- Schott, B.H.; Minuzzi, L.; Krebs, R.M.; Elmenhorst, D.; Lang, M.; Winz, O.H.; Seidenbecher, C.I.; Coenen, H.H.; Heinze, H.-J.; Zilles, K.; et al. Mesolimbic Functional Magnetic Resonance Imaging Activations during Reward Anticipation Correlate with Reward-Related Ventral Striatal Dopamine Release. J. Neurosci. 2008, 28, 14311–14319. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, S.J.; Pizzagalli, D.A.; Sproule, B.A.; Kennedy, S.H. Assessing anhedonia in depression: Potentials and pitfalls. Neurosci. Biobehav. Rev. 2016, 65, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Treadway, M.T.; Zald, D.H. Reconsidering anhedonia in depression: Lessons from translational neuroscience. Neurosci. Biobehav. Rev. 2011, 35, 537–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, K.R.; Whybrow, P.C.; Kringelbach, M.L. Reconceptualizing anhedonia: Novel perspectives on balancing the pleasure networks in the human brain. Front. Behav. Neurosci. 2015, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Rupprechter, S.; Romaniuk, L.; Series, P.; Hirose, Y.; Hawkins, E.; Sandu, A.-L.; Waiter, G.D.; McNeil, C.J.; Shen, X.; A Harris, M.; et al. Blunted medial prefrontal cortico-limbic reward-related effective connectivity and depression. Brain 2020, 143, 1946–1956. [Google Scholar] [CrossRef]
- Mao, L.-M.; Wang, J.Q. Alterations in mGlu5 receptor expression and function in the striatum in a rat depression model. J. Neurochem. 2018, 145, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Lener, M.S.; Kadriu, B.; Zarate, C.A. Ketamine and Beyond: Investigations into the Potential of Glutamatergic Agents to Treat Depression. Drugs 2017, 77, 381–401. [Google Scholar] [CrossRef] [Green Version]
- Corriger, A.; Pickering, G. Ketamine and depression: A narrative review. Drug Des. Dev. Ther. 2019, 13, 3051–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraus, C.; Rabl, U.; Vanicek, T.; Carlberg, L.; Popovic, A.; Spies, M.; Bartova, L.; Gryglewski, G.; Papageorgiou, K.; Lanzenberger, R.; et al. Administration of ketamine for unipolar and bipolar depression. Int. J. Psychiatry Clin. Pract. 2017, 21, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, R.-J.; Dwyer, J.M.; Banasr, M.; Lee, B.; Son, H.; Li, X.-Y.; Aghajanian, G.; Duman, R.S. Glutamate N-methyl-D-aspartate Receptor Antagonists Rapidly Reverse Behavioral and Synaptic Deficits Caused by Chronic Stress Exposure. Biol. Psychiatry 2011, 69, 754–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salagre, E.; Grande, I.; Solé, B.; Sanchez-Moreno, J.; Vieta, E. Vortioxetine: A new alternative for the treatment of major depressive disorder. Rev. Psiquiatr. Salud Ment. 2018, 11, 48–59. [Google Scholar] [CrossRef]
- Grace, A.A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat. Rev. Neurosci. 2016, 17, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Arias-Carrión, Ó.; Pöppel, E. Dopamine, learning, and reward-seeking behavior. Acta Neurobiol. Exp. 2007, 67, 481–488. Available online: https://www.researchgate.net/profile/Ernst_Poeppel/publication/5531082_Dopamine_learning_and_reward-seeking_behavior/links/54db98010cf23fe133ad631e.pdf (accessed on 15 March 2020).
- Szczypiński, J.J.; Gola, M. Dopamine dysregulation hypothesis: The common basis for motivational anhedonia in major depressive disorder and schizophrenia? Rev. Neurosci. 2018, 29, 727–744. [Google Scholar] [CrossRef]
- Piray, P.; den Ouden, H.E.; van der Schaaf, M.E.; Toni, I.; Cools, R. Dopaminergic Modulation of the Functional Ventrodorsal Ar-chitecture of the Human Striatum. Cereb. Cortex 2017, 27, 485–495. [Google Scholar] [PubMed] [Green Version]
- Knutson, B.; Westdorp, A.; Kaiser, E.; Hommer, D. FMRI Visualization of Brain Activity during a Monetary Incentive Delay Task. Neuroimage 2000, 12, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Benningfield, M.M.; Blackford, J.U.; Ellsworth, M.E.; Samanez-Larkin, G.R.; Martin, P.R.; Cowan, R.L.; Zald, D.H. Caudate responses to reward anticipation associated with delay discounting behavior in healthy youth. Dev. Cogn. Neurosci. 2014, 7, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haber, S.N.; Knutson, B. The Reward Circuit: Linking Primate Anatomy and Human Imaging. Neuropsychopharmacology 2009, 35, 4–26. [Google Scholar] [CrossRef] [Green Version]
- Berridge, K.C.; E Robinson, T.; Aldridge, J.W. Dissecting components of reward: ‘Liking’, ‘wanting’, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Kringelbach, M.L.; Berridge, K.C. Towards a functional neuroanatomy of pleasure and happiness. Trends Cogn. Sci. 2009, 13, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Schultz, W. Multiple reward signals in the brain. Nat. Rev. Neurosci. 2000, 1, 199–207. [Google Scholar] [CrossRef]
- Schultz, W.; Tremblay, L.; Hollerman, J.R. Reward Processing in Primate Orbitofrontal Cortex and Basal Ganglia. Cereb. Cortex 2000, 10, 272–284. [Google Scholar] [CrossRef]
- Takahashi, T.; Yűcel, M.; Lorenzetti, V.; Nakamura, K.; Whittle, S.; Walterfang, M.; Suzuki, M.; Pantelis, C.; Allen, N.B. Midline brain structures in patients with current and remitted major depression. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2009, 33, 1058–1063. [Google Scholar] [CrossRef] [PubMed]
- Knutson, B.; Taylor, J.; Kaufman, M.; Peterson, R.; Glover, G. Distributed Neural Representation of Expected Value. J. Neurosci. 2005, 25, 4806–4812. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M.; Zatorre, R.J.; Dagher, A.; Evans, A.C.; Jones-Gotman, M. Changes in brain activity related to eating chocolate: From pleasure to aversion. Brain 2001, 124, 1720–1733. [Google Scholar] [CrossRef] [Green Version]
- Richards, J.M.; Plate, R.C.; Ernst, M. A systematic review of fMRI reward paradigms used in studies of adolescents vs. adults: The impact of task design and implications for understanding neurodevelopment. Neurosci. Biobehav. Rev. 2013, 37, 976–991. [Google Scholar] [CrossRef] [Green Version]
- Nusslock, R.; Alloy, L.B. Reward processing and mood-related symptoms: An RDoC and translational neuroscience perspective. J. Affect. Disord. 2017, 216, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Alloy, L.B.; Olino, T.; Freed, R.D.; Nusslock, R. Role of Reward Sensitivity and Processing in Major Depressive and Bipolar Spectrum Disorders. Behav. Ther. 2016, 47, 600–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyropoulos, S.V.; Nutt, D.J. Anhedonia revisited: Is there a role for dopamine-targeting drugs for depression? J. Psychopharmacol. 2013, 27, 869–877. [Google Scholar] [CrossRef]
- Pechtel, P.; Dutra, S.J.; Goetz, E.L.; Pizzagalli, D.A. Blunted reward responsiveness in remitted depression. J. Psychiatr. Res. 2013, 47, 1864–1869. [Google Scholar] [CrossRef] [Green Version]
- Roth-Deri, I.; Friedman, A.; Abraham, L.; Lax, E.; Flaumenhaft, Y.; Dikshtein, Y.; Yadid, G. Antidepressant treatment facilitates dopamine release and drug seeking behavior in a genetic animal model of depression. Eur. J. Neurosci. 2009, 30, 485–492. [Google Scholar] [CrossRef]
- Dias-Ferreira, E.; Sousa, J.C.; Melo, I.; Morgado, P.; Mesquita, A.R.; Cerqueira, J.J.; Costa, R.M.; Sousa, N. Chronic Stress Causes Frontostriatal Reorganization and Affects Decision-Making. Science 2009, 325, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Burton, A.C.; Nakamura, K.; Roesch, M.R. From ventral-medial to dorsal-lateral striatum: Neural correlates of reward-guided decision-making. Neurobiol. Learn. Mem. 2015, 117, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Thomason, M.E.; Marusak, H.A. Within-subject neural reactivity to reward and threat is inverted in young adolescents. Psychol. Med. 2017, 47, 1549–1560. [Google Scholar] [CrossRef]
- Yang, J.-H.; Liao, R.-M. Dissociable contribution of nucleus accumbens and dorsolateral striatum to the acquisition of risk choice behavior in the rat. Neurobiol. Learn. Mem. 2015, 126, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Jacquet, M.; Lecourtier, L.; Cassel, R.; Loureiro, M.; Cosquer, B.; Escoffier, G.; Migliorati, M.; Cassel, J.-C.; Roman, F.; Marchetti, E. Dorsolateral striatum and dorsal hippocampus: A serial contribution to acquisition of cue-reward associations in rats. Behav. Brain Res. 2013, 239, 94–103. [Google Scholar] [CrossRef]
- Thapa, R.; Gruber, A.J. Lesions of ventrolateral striatum eliminate lose-shift but not win-stay behaviour in rats. Neurobiol. Learn. Mem. 2018, 155, 446–451. [Google Scholar] [CrossRef]
- Cox, J.; Witten, I.B. Striatal circuits for reward learning and decision-making. Nat. Rev. Neurosci. 2019, 20, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Stalnaker, T.A.; Calhoon, G.G.; Ogawa, M.; Roesch, M.R.; Schoenbaum, G. Neural correlates of stimulus-response and response-outcome associations in dorsolateral versus dorsomedial striatum. Front. Integr. Neurosci. 2010, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voorn, P.; Vanderschuren, L.J.; Groenewegen, H.J.; Robbins, T.W.; Pennartz, C.M. Putting a spin on the dorsal–ventral divide of the striatum. Trends Neurosci. 2004, 27, 468–474. [Google Scholar] [CrossRef]
- Tepper, J.M.; Bolam, J.P. Functional diversity and specificity of neostriatal interneurons. Curr. Opin. Neurobiol. 2004, 14, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Dudman, J.T.; Gerfen, C.R. The Basal Ganglia. In The Rat Nervous System, 4th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 391–440. [Google Scholar] [CrossRef]
- Kreitzer, A.C.; Berke, J.D. Investigating striatal function through cell-type-specific manipulations. Neuroscience 2011, 198, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Nagayoshi, I.; Nishi, A.; Fukuda, T. Three divisions of the mouse caudal striatum differ in the proportions of dopamine D1 and D2 receptor-expressing cells, distribution of dopaminergic axons, and composition of cholinergic and GABAergic interneurons. Brain Struct. Funct. 2019, 224, 2703–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.G.; Marshall, J.D.; Ahanonu, B.; Wu, Y.-W.; Kim, T.H.; Grewe, B.F.; Zhang, Y.; Li, J.Z.; Ding, J.B.; Ehlers, M.D.; et al. Diametric neural ensemble dynamics in parkinsonian and dyskinetic states. Nat. Cell Biol. 2018, 557, 177–182. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Ding, J.; Day, M.; Wang, Z.; Shen, W. D1 and D2 dopamine-receptor modulation of striatal glutamatergic signaling in striatal medium spiny neurons. Trends Neurosci. 2007, 30, 228–235. [Google Scholar] [CrossRef]
- Graybiel, A.M.; Ragsdale, C.W. Histochemically distinct compartments in the striatum of human, monkeys, and cat demonstrated by acetylthiocholinesterase staining. Proc. Natl. Acad. Sci. USA 1978, 75, 5723–5726. [Google Scholar] [CrossRef] [Green Version]
- Eblen, F.; Graybiel, A. Highly restricted origin of prefrontal cortical inputs to striosomes in the macaque monkey. J. Neurosci. 1995, 15, 5999–6013. [Google Scholar] [CrossRef] [Green Version]
- Gerfen, C.R. The neostriatal mosaic: Compartmentalization of corticostriatal input and striatonigral output systems. Nat. Cell Biol. 1984, 311, 461–464. [Google Scholar] [CrossRef]
- Smith, J.B.; Klug, J.R.; Ross, D.L.; Howard, C.D.; Hollon, N.G.; Ko, V.I.; Hoffman, H.; Callaway, E.M.; Gerfen, C.R.; Jin, X. Genetic-Based Dissection Unveils the Inputs and Outputs of Striatal Patch and Matrix Compartments. Neuron 2016, 91, 1069–1084. [Google Scholar] [CrossRef] [Green Version]
- Crittenden, J.R.; Tillberg, P.W.; Riad, M.H.; Shima, Y.; Gerfen, C.R.; Curry, J.; Housman, D.E.; Nelson, S.B.; Boyden, E.S.; Graybiel, A.M. Striosome–dendron bouquets highlight a unique striatonigral circuit targeting dopamine-containing neurons. Proc. Natl. Acad. Sci. USA 2016, 113, 11318–11323. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.; Amemori, S.; Chung, E.; Gibson, D.J.; Amemori, K.-I.; Graybiel, A.M. Predominant Striatal Input to the Lateral Habenula in Macaques Comes from Striosomes. Curr. Biol. 2019, 29, 51–61.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watabe-Uchida, M.; Zhu, L.; Ogawa, S.K.; Vamanrao, A.; Uchida, N. Whole-Brain Mapping of Direct Inputs to Midbrain Dopamine Neurons. Neuron 2012, 74, 858–873. [Google Scholar] [CrossRef] [Green Version]
- Graybiel, A.M. Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci. 1990, 13, 244–254. [Google Scholar] [CrossRef]
- Crittenden, J.R.; Graybiel, A.M. Basal Ganglia Disorders Associated with Imbalances in the Striatal Striosome and Matrix Compartments. Front. Neuroanat. 2011, 5, 59. [Google Scholar] [CrossRef] [Green Version]
- Rajakumar, N.; Elisevich, K.; Flumerfelt, B.A. Compartmental origin of the striato-entopeduncular projection in the rat. J. Comp. Neurol. 1993, 331, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Stephenson-Jones, M.; Kardamakis, A.A.; Robertson, B.; Grillner, S. Independent circuits in the basal ganglia for the evaluation and selection of actions. Proc. Natl. Acad. Sci. USA 2013, 110, E3670–E3679. [Google Scholar] [CrossRef] [Green Version]
- Fujiyama, F.; Sohn, J.; Nakano, T.; Furuta, T.; Nakamura, K.C.; Matsuda, W.; Kaneko, T. Exclusive and common targets of neostriatofugal projections of rat striosome neurons: A single neuron-tracing study using a viral vector. Eur. J. Neurosci. 2011, 33, 668–677. [Google Scholar] [CrossRef] [PubMed]
- McGregor, M.M.; McKinsey, G.L.; Girasole, A.E.; Bair-Marshall, C.J.; Rubenstein, J.L.; Nelson, A.B. Functionally Distinct Connectivity of Developmentally Targeted Striosome Neurons. Cell Rep. 2019, 29, 1419–1428.e5. [Google Scholar] [CrossRef]
- Evans, R.C.; Twedell, E.L.; Zhu, M.; Ascencio, J.; Zhang, R.; Khaliq, Z.M. Functional Dissection of Basal Ganglia Inhibitory Inputs onto Substantia Nigra Dopaminergic Neurons. Cell Rep. 2020, 32. [Google Scholar] [CrossRef]
- Haber, S.N.; Fudge, J.L.; McFarland, N.R. Striatonigrostriatal Pathways in Primates Form an Ascending Spiral from the Shell to the Dorsolateral Striatum. J. Neurosci. 2000, 20, 2369–2382. [Google Scholar] [CrossRef]
- Lerner, T.N.; Shilyansky, C.; Davidson, T.J.; Evans, K.E.; Beier, K.T.; Zalocusky, K.A.; Crow, A.K.; Malenka, R.C.; Luo, L.; Tomer, R.; et al. Intact-Brain Analyses Reveal Distinct Information Carried by SNc Dopamine Subcircuits. Cell 2015, 162, 635–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tritsch, N.X.; Sabatini, B.L. Dopaminergic Modulation of Synaptic Transmission in Cortex and Striatum. Neuron 2012, 76, 33–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurd, Y.L.; Herman, M.M.; Hyde, T.M.; Bigelow, L.B.; Weinberger, D.R.; E Kleinman, J. Prodynorphin mRNA expression is increased in the patch vs matrix compartment of the caudate nucleus in suicide subjects. Mol. Psychiatry 1997, 2, 495–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kano, M.; Ohno-Shosaku, T.; Hashimotodani, Y.; Uchigashima, M.; Watanabe, M. Endocannabinoid-Mediated Control of Synaptic Transmission. Physiol. Rev. 2009, 89, 309–380. [Google Scholar] [CrossRef]
- Piomelli, D. The molecular logic of endocannabinoid signalling. Nat. Rev. Neurosci. 2003, 4, 873–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marco, E.M.; Echeverry-Alzate, V.; López-Moreno, J.A.; Giné, E.; Peñasco, S.; Viveros, M.P. Consequences of early life stress on the expression of endocannabinoid-related genes in the rat brain. Behav. Pharmacol. 2014, 25, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V. Endocannabinoid signaling in the brain: Biosynthetic mechanisms in the limelight. Nat. Neurosci. 2010, 14, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Hillard, C.J. The Endocannabinoid Signaling System in the CNS. In International Review of Neurobiology; Academic Press Inc.: Cambridge, MA, USA, 2015; Volume 125, pp. 1–47. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the Presence and Functional Expression of Cannabinoid CB2 Receptors in Brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef] [PubMed]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Identification and Functional Characterization of Brainstem Cannabinoid CB2 Receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [Green Version]
- Cabral, G.A.; Raborn, E.S.; Griffin, L.; Dennis, J.; Marciano-Cabral, F. CB2 receptors in the brain: Role in central immune function. Br. J. Pharmacol. 2008, 153, 240–251. [Google Scholar] [CrossRef] [Green Version]
- Rana, T.; Behl, T.; Sehgal, A.; Mehta, V.; Singh, S.; Kumar, R.; Bungau, S. Integrating Endocannabinoid Signalling In Depression. J. Mol. Neurosci. 2021, 2021, 1–13. [Google Scholar] [CrossRef]
- Fitzgerald, M.L.; Shobin, E.; Pickel, V.M. Cannabinoid modulation of the dopaminergic circuitry: Implications for limbic and striatal output. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2012, 38, 21–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovinger, D.M.; Mathur, B.N. Endocannabinoids in striatal plasticity. Park. Relat. Disord. 2012, 18, S132–S134. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R.; Cabañero, D.; Martín-García, E. The endocannabinoid system in modulating fear, anxiety, and stress. Dialog. Clin. Neurosci. 2020, 22, 229–239. [Google Scholar] [CrossRef]
- Hwang, E.-S.; Kim, H.-B.; Lee, S.; Kim, M.-J.; Kim, K.-J.; Han, G.; Han, S.-Y.; Lee, E.-A.; Yoon, J.-H.; Kim, D.-O.; et al. Antidepressant-like effects of β-caryophyllene on restraint plus stress-induced depression. Behav. Brain Res. 2020, 380, 112439. [Google Scholar] [CrossRef]
- Lazary, J.; Eszlari, N.; Juhasz, G.; Bagdy, G. A functional variant of CB2 receptor gene interacts with childhood trauma and FAAH gene on anxious and depressive phenotypes. J. Affect. Disord. 2019, 257, 716–722. [Google Scholar] [CrossRef]
- Horder, J.; Harmer, C.J.; Cowen, P.J.; McCabe, C. Reduced neural response to reward following 7 days treatment with the cannabinoid CB1 antagonist rimonabant in healthy volunteers. Int. J. Neuropsychopharmacol. 2010, 13, 1103–1113. [Google Scholar] [CrossRef]
- Sanchis-Segura, C.; Cline, B.H.; Marsicano, G.; Lutz, B.; Spanagel, R. Reduced sensitivity to reward in CB1 knockout mice. Psychopharmacology 2004, 176, 223–232. [Google Scholar] [CrossRef]
- Gessa, G.; Melis, M.; Muntoni, A.; Diana, M. Cannabinoids activate mesolimbic dopamine neurons by an action on cannabinoid CB1 receptors. Eur. J. Pharmacol. 1998, 341, 39–44. [Google Scholar] [CrossRef]
- Coccurello, R. Anhedonia in depression symptomatology: Appetite dysregulation and defective brain reward processing. Behav. Brain Res. 2019, 372, 112041. [Google Scholar] [CrossRef] [PubMed]
- Rincón-Cortés, M.; Grace, A.A. Antidepressant effects of ketamine on depression-related phenotypes and dopamine dysfunction in rodent models of stress. Behav. Brain Res. 2020, 379, 112367. [Google Scholar] [CrossRef]
- Ferber, S.G.; Roth, T.L.; Weller, A. Epigenetic fragility of the endocannabinoid system under stress: Risk for mood disorders and pharmacogenomic implications. Epigenomics 2020, 12, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Ferber, S.G.; Trezza, V.; Weller, A. Early life stress and development of the endocannabinoid system: A bidirectional process in programming future coping. Dev. Psychobiol. 2019. [Google Scholar] [CrossRef]
- Lomazzo, E.; König, F.; Abassi, L.; Jelinek, R.; Lutz, B. Chronic stress leads to epigenetic dysregulation in the neuropeptide-Y and cannabinoid CB1 receptor genes in the mouse cingulate cortex. Neuropharmacology 2017, 113, 301–313. [Google Scholar] [CrossRef]
- Hayase, T. Putative Epigenetic Involvement of the Endocannabinoid System in Anxiety- and Depression-Related Behaviors Caused by Nicotine as a Stressor. PLoS ONE 2016, 11, e0158950. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Calvo, A.; Maroto, I.B.; Bajo-Grañeras, R.; Chiarlone, A.; Gaudioso, Á.; Ferrero, J.J.; Resel, E.; Sánchez-Prieto, J.; Rodríguez-Navarro, J.A.; Marsicano, G.; et al. Pathway-Specific Control of Striatal Neuron Vulnerability by Corticostriatal Cannabinoid CB1 Receptors—PubMed. Cereb. Cortex 2018, 28, 307–322. [Google Scholar] [CrossRef]
- Monory, K.; Blaudzun, H.; Massa, F.; Kaiser, N.; Lemberger, T.; Schütz, G.; Wotjak, C.T.; Lutz, B.; Marsicano, G. Genetic Dissection of Behavioural and Autonomic Effects of Δ9-Tetrahydrocannabinol in Mice. PLoS Biol. 2007, 5, e269. [Google Scholar] [CrossRef]
- Hermann, H.; Marsicano, G.; Lutz, B. Coexpression of the cannabinoid receptor type 1 with dopamine and serotonin receptors in distinct neuronal subpopulations of the adult mouse forebrain. Neuroscience 2002, 109, 451–460. [Google Scholar] [CrossRef]
- Centonze, D.; Battista, N.; Rossi, S.; Mercuri, N.B.; Finazzi-Agrò, A.; Bernardi, G.; Calabresi, P.; Maccarrone, M. A Critical Interaction between Dopamine D2 Receptors and Endocannabinoids Mediates the Effects of Cocaine on Striatal GABAergic Transmission. Neuropsychopharmacology 2004, 29, 1488–1497. [Google Scholar] [CrossRef] [Green Version]
- Katona, I.; Freund, T.F. Multiple Functions of Endocannabinoid Signaling in the Brain. Annu. Rev. Neurosci. 2012, 35, 529–558. [Google Scholar] [CrossRef] [Green Version]
- Bamford, N.S.; Zhang, H.; Schmitz, Y.; Wu, N.-P.; Cepeda, C.; Levine, M.S.; Schmauss, C.; Zakharenko, S.S.; Zablow, L.; Sulzer, D. Heterosynaptic Dopamine Neurotransmission Selects Sets of Corticostriatal Terminals. Neuron 2004, 42, 653–663. [Google Scholar] [CrossRef] [Green Version]
- Kreitzer, A.C. Physiology and Pharmacology of Striatal Neurons. Annu. Rev. Neurosci. 2009, 32, 127–147. [Google Scholar] [CrossRef]
- Castillo, P.E.; Younts, T.J.; Chávez, A.E.; Hashimotodani, Y. Endocannabinoid Signaling and Synaptic Function. Neuron 2012, 76, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Ruiz, J.; Moreno-Martet, M.; Rodríguez-Cueto, C.; Palomo-Garo, C.; Gómez-Cañas, M.; Valdeolivas, S.; Guaza, C.; Romero, J.; Guzmán, M.; Mechoulam, R.; et al. Prospects for cannabinoid therapies in basal ganglia disorders. Br. J. Pharmacol. 2011, 163, 1365–1378. [Google Scholar] [CrossRef] [Green Version]
- Blazquez, C.; Chiarlone, A.; Bellocchio, L.; Resel, E.; Pruunsild, P.; García-Rincón, D.; Sendtner, M.; Timmusk, T.; Lutz, B.; Galveroperh, I.; et al. The CB1 cannabinoid receptor signals striatal neuroprotection via a PI3K/Akt/mTORC1/BDNF pathway. Cell Death Differ. 2015, 22, 1618–1629. [Google Scholar] [CrossRef] [PubMed]
- Mievis, S.; Blum, D.; Ledent, C. Worsening of Huntington disease phenotype in CB1 receptor knockout mice. Neurobiol. Dis. 2011, 42, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Chiarlone, A.; Bellocchio, L.; Blázquez, C.; Resel, E.; Soria-Gómez, E.; Cannich, A.; Ferrero, J.J.; Sagredo, O.; Benito, C.; Romero, J.; et al. A restricted population of CB1 cannabinoid receptors with neuroprotective activity. Proc. Natl. Acad. Sci. USA 2014, 111, 8257–8262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirayama, Y. Neurochemistry of the Nucleus Accumbens and its Relevance to Depression and Antidepressant Action in Rodents. Curr. Neuropharmacol. 2006, 4, 277–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belujon, P.; Grace, A.A. Regulation of dopamine system responsivity and its adaptive and pathological response to stress. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S.; Morrison, J.H. The Brain on Stress: Vulnerability and Plasticity of the Prefrontal Cortex over the Life Course. Neuron 2013, 79, 16–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rademacher, D.J.; Hillard, C.J. Interactions between endocannabinoids and stress-induced decreased sensitivity to natural reward. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2007, 31, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubreucq, S.; Matias, I.; Cardinal, P.; Häring, M.; Lutz, B.; Marsicano, G.; Chaouloff, F. Genetic Dissection of the Role of Cannabinoid Type-1 Receptors in the Emotional Consequences of Repeated Social Stress in Mice. Neuropsychopharmacology 2012, 37, 1885–1900. [Google Scholar] [CrossRef] [Green Version]
- Knowland, D.; Lim, B.K. Circuit-based frameworks of depressive behaviors: The role of reward circuitry and beyond. Pharmacol. Biochem. Behav. 2018, 174, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Ottenheimer, D.; Richard, J.M.; Janak, P.H. Ventral pallidum encodes relative reward value earlier and more robustly than nucleus accumbens. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Gardner, E.L. Addiction and Brain Reward and Antireward Pathways. Asthma Soc. Psychol. Factors Psychosom. Syndr. 2011, 30, 22–60. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.S.; Tindell, A.J.; Aldridge, J.W.; Berridge, K.C. Ventral pallidum roles in reward and motivation. Behav. Brain Res. 2009, 196, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Bracci, E.; Centonze, D.; Bernardi, G.; Calabresi, P. Dopamine Excites Fast-Spiking Interneurons in the Striatum. J. Neurophysiol. 2002, 87, 2190–2194. [Google Scholar] [CrossRef] [Green Version]
- Laurikainen, H.; Tuominen, L.; Tikka, M.; Merisaari, H.; Armio, R.-L.; Sormunen, E.; Borgan, F.; Veronese, M.; Howes, O.; Haaparanta-Solin, M.; et al. Sex difference in brain CB1 receptor availability in man. Neuroimage 2019, 184, 834–842. [Google Scholar] [CrossRef] [Green Version]
- Akil, H.; Gordon, J.; Hen, R.; Javitch, J.; Mayberg, H.; McEwen, B.; Meaney, M.J.; Nestler, E.J. Treatment resistant depression: A multi-scale, systems biology approach. Neurosci. Biobehav. Rev. 2018, 84, 272–288. [Google Scholar] [CrossRef] [PubMed]
- Gaynes, B.N.; Warden, D.; Trivedi, M.H.; Wisniewski, S.R.; Fava, M.; Rush, A.J. What Did STAR*D Teach Us? Results From a Large-Scale, Practical, Clinical Trial for Patients With Depression. Psychiatr. Serv. 2009, 60, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.M.; Tekell, J.L.; Bowden, C.L.; Brannan, S.; Houston, J.P.; Berman, N.; Frazer, A. Onset and Early Behavioral Effects of Pharmacologically Different Antidepressants and Placebo in Depression. Neuropsychopharmacology 2003, 29, 566–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spijker, J.; Bijl, R.V.; De Graaf, R.; Nolen, W.A. Determinants of poor 1-year outcome of DSM-III-R major depression in the general population: Results of the Netherlands Mental Health Survey and Incidence Study (NEMESIS). Acta Psychiatr. Scand. 2001, 103, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Kringelbach, M.L. Pleasure Systems in the Brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef] [Green Version]
- Martin-Soelch, C. Is depression associated with dysfunction of the central reward system? Biochem. Soc. Trans. 2009, 37, 313–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shbiro, L.; Hen-Shoval, D.; Hazut, N.; Rapps, K.; Dar, S.; Zalsman, G.; Mechoulam, R.; Weller, A.; Shoval, G. Effects of cannabidiol in males and females in two different rat models of depression. Physiol. Behav. 2019, 201, 59–63. [Google Scholar] [CrossRef]
- Shoval, G.; Shbiro, L.; Hershkovitz, L.; Hazut, N.; Zalsman, G.; Mechoulam, R.; Weller, A. Prohedonic Effect of Cannabidiol in a Rat Model of Depression. Neuropsychobiology 2016, 73, 123–129. [Google Scholar] [CrossRef]
- Nutt, D.J.; Demyttenaere, K.; Janka, Z.; Aarre, T.; Bourin, M.; Canonico, P.L.; Carrasco, J.L.; Stahl, S. The other face of depression, reduced positive affect: The role of catecholamines in causation and cure. J. Psychopharmacol. 2006, 21, 461–471. [Google Scholar] [CrossRef]
- McCabe, C.; Cowen, P.J.; Harmer, C.J. Neural representation of reward in recovered depressed patients. Psychopharmacology 2009, 205, 667–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prisco, S.; Esposito, E. Differential effects of acute and chronic fluoxetine administration on the spontaneous activity of dopaminergic neurones in the ventral tegmental area. Br. J. Pharmacol. 1995, 116, 1923–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mascio, M.; Di Giovanni, G.; Di Matteo, V.; Prisco, S.; Esposito, E. Selective serotonin reuptake inhibitors reduce the spontaneous activity of dopaminergic neurons in the ventral tegmental area. Brain Res. Bull. 1998, 46, 547–554. [Google Scholar] [CrossRef]
- Dremencov, E.; El Mansari, M.; Blier, P. Effects of sustained serotonin reuptake inhibition on the firing of dopamine neurons in the rat ventral tegmental area. J. Psychiatry Neurosci. 2009, 34, 223–229. [Google Scholar] [PubMed]
- Juckel, G.; Schlagenhauf, F.; Koslowski, M.; Filonov, D.; Wüstenberg, T.; Villringer, A.; Knutson, B.; Kienast, T.; Gallinat, J.; Wrase, J.; et al. Dysfunction of ventral striatal reward prediction in schizophrenic patients treated with typical, not atypical, neuroleptics. Psychopharmacology 2006, 187, 222–228. [Google Scholar] [CrossRef]
- Schlagenhauf, F.; Sterzer, P.; Schmack, K.; Ballmaier, M.; Rapp, M.; Wrase, J.; Juckel, G.; Gallinat, J.; Heinz, A. Reward Feedback Alterations in Unmedicated Schizophrenia Patients: Relevance for Delusions. Biol. Psychiatry 2009, 65, 1032–1039. [Google Scholar] [CrossRef]
- Seeman, P. Atypical Antipsychotics: Mechanism of Action. Can. J. Psychiatry 2002, 47, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Belujon, P.; Jakobowski, N.L.; Dollish, H.K.; A Grace, A. Withdrawal from Acute Amphetamine Induces an Amygdala-Driven Attenuation of Dopamine Neuron Activity: Reversal by Ketamine. Neuropsychopharmacology 2015, 41, 619–627. [Google Scholar] [CrossRef] [Green Version]
- Norris, C.; Loureiro, M.; Kramar, C.; Zunder, J.; Renard, J.; Rushlow, W.; LaViolette, S.R. Cannabidiol Modulates Fear Memory Formation Through Interactions with Serotonergic Transmission in the Mesolimbic System. Neuropsychopharmacology 2016, 41, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Seeman, P. Cannabidiol is a partial agonist at dopamine D2High receptors, predicting its antipsychotic clinical dose. Transl. Psychiatry 2016, 6, e920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, A.C.; Moreira, F.A.; Gomes, F.V.; Del Bel, E.A.; Guimarães, F.S. Multiple mechanisms involved in the large-spectrum therapeutic potential of cannabidiol in psychiatric disorders. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3364–3378. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; Buuse, M.V.D. Is the mTOR-signalling cascade disrupted in Schizophrenia? J. Neurochem. 2014, 129, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Pham, X.; Zhang, L.; Chen, P.-L.; Burzynski, G.M.; McGaughey, D.M.; He, S.; McGrath, J.A.; Wolyniec, P.S.; Fallin, M.D.; et al. Functional Variants in DPYSL2 Sequence Increase Risk of Schizophrenia and Suggest a Link to mTOR Signaling. G3 Genes Genomes Genet. 2015, 5, 61–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renard, J.; Loureiro, M.; Rosen, L.G.; Zunder, J.; De Oliveira, C.; Schmid, S.; Rushlow, W.J.; LaViolette, S.R. Cannabidiol Counteracts Amphetamine-Induced Neuronal and Behavioral Sensitization of the Mesolimbic Dopamine Pathway through a Novel mTOR/p70S6 Kinase Signaling Pathway. J. Neurosci. 2016, 36, 5160–5169. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.B.; Burnett, A.; Hall, B.; Parker, K.K. Agonistic Properties of Cannabidiol at 5-HT1a Receptors. Neurochem. Res. 2005, 30, 1037–1043. [Google Scholar] [CrossRef]
- Bisogno, T.; Hanuš, L.; De Petrocellis, L.; Tchilibon, S.; E Ponde, D.; Brandi, I.; Moriello, A.S.; Davis, J.B.; Mechoulam, R.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001, 134, 845–852. [Google Scholar] [CrossRef]
- Hudson, R.; Rushlow, W.; LaViolette, S.R. Phytocannabinoids modulate emotional memory processing through interactions with the ventral hippocampus and mesolimbic dopamine system: Implications for neuropsychiatric pathology. Psychopharmacology 2018, 235, 447–458. [Google Scholar] [CrossRef]
- Kusmider, M.; Faron-Górecka, A.; Solich, J.; Pabian, P.; Dziedzicka-Wasylewska, M. Time-course of changes in key catecholaminergic receptors and trophic systems in rat brain after antidepressant administration. Neurochem. Int. 2020, 141, 104885. [Google Scholar] [CrossRef]
- Salort, G.; Hernández-Hernández, E.; García-Fuster, M.J.; García-Sevilla, J.A. Regulation of cannabinoid CB1 and CB2 receptors, neuroprotective mTOR and pro-apoptotic JNK1/2 kinases in postmortem prefrontal cortex of subjects with major depressive disorder. J. Affect. Disord. 2020, 276, 626–635. [Google Scholar] [CrossRef]
- Xue, S.-S.; Xue, F.; Ma, Q.-R.; Wang, S.-Q.; Wang, Y.; Tan, Q.-R.; Wang, H.-N.; Zhou, C.-H.; Peng, Z.-W. Repetitive high-frequency transcranial magnetic stimulation reverses depressive-like behaviors and protein expression at hippocampal synapses in chronic unpredictable stress-treated rats by enhancing endocannabinoid signaling. Pharmacol. Biochem. Behav. 2019, 184, 172738. [Google Scholar] [CrossRef]
- Shuto, T.; Kuroiwa, M.; Sotogaku, N.; Kawahara, Y.; Oh, Y.-S.; Jang, J.-H.; Shin, C.-H.; Ohnishi, Y.N.; Hanada, Y.; Miyakawa, T.; et al. Obligatory roles of dopamine D1 receptors in the dentate gyrus in antidepressant actions of a selective serotonin reuptake inhibitor, fluoxetine. Mol. Psychiatry 2020, 25, 1229–1244. [Google Scholar] [CrossRef] [PubMed]
- Arnsten, A.F.T. Stress weakens prefrontal networks: Molecular insults to higher cognition. Nat. Neurosci. 2015, 18, 1376–1385. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldstein Ferber, S.; Weller, A.; Yadid, G.; Friedman, A. Discovering the Lost Reward: Critical Locations for Endocannabinoid Modulation of the Cortico–Striatal Loop That Are Implicated in Major Depression. Int. J. Mol. Sci. 2021, 22, 1867. https://doi.org/10.3390/ijms22041867
Goldstein Ferber S, Weller A, Yadid G, Friedman A. Discovering the Lost Reward: Critical Locations for Endocannabinoid Modulation of the Cortico–Striatal Loop That Are Implicated in Major Depression. International Journal of Molecular Sciences. 2021; 22(4):1867. https://doi.org/10.3390/ijms22041867
Chicago/Turabian StyleGoldstein Ferber, Sari, Aron Weller, Gal Yadid, and Alexander Friedman. 2021. "Discovering the Lost Reward: Critical Locations for Endocannabinoid Modulation of the Cortico–Striatal Loop That Are Implicated in Major Depression" International Journal of Molecular Sciences 22, no. 4: 1867. https://doi.org/10.3390/ijms22041867
APA StyleGoldstein Ferber, S., Weller, A., Yadid, G., & Friedman, A. (2021). Discovering the Lost Reward: Critical Locations for Endocannabinoid Modulation of the Cortico–Striatal Loop That Are Implicated in Major Depression. International Journal of Molecular Sciences, 22(4), 1867. https://doi.org/10.3390/ijms22041867