
Contributions of TOR Signaling on Photosynthesis


1
School of Life Sciences, Liaocheng University, Liaocheng 252000, China
2
Department of Integrative Agriculture, College of Food and Agriculture, United Arab Emirates University, Al Ain 15551, United Arab Emirates
3
Institute of Urban Agriculture, Chinese Academy of Agricultural Sciences, Chengdu 610213, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(16), 8959; https://doi.org/10.3390/ijms22168959
Submission received: 30 June 2021
/
Revised: 22 July 2021
/
Accepted: 29 July 2021
/
Published: 20 August 2021
(This article belongs to the Special Issue Photosynthesis 2.0)
{kind=link}
{kind=link}
Abstract
:The target of rapamycin (TOR) protein kinase is an atypical Ser/Thr protein kinase and evolutionally conserved among yeasts, plants, and mammals. TOR has been established as a central hub for integrating nutrient, energy, hormone, and environmental signals in all the eukaryotes. Despite the conserved functions across eukaryotes, recent research has shed light on the multifaceted roles of TOR signaling in plant-specific functional and mechanistic features. One of the most specific features is the involvement of TOR in plant photosynthesis. The recent development of tools for the functional analysis of plant TOR has helped to uncover the involvement of TOR signaling in several steps preceding photoautotrophy and maintenance of photosynthesis. Here, we present recent novel findings relating to TOR signaling and its roles in regulating plant photosynthesis, including carbon nutrient sense, light absorptions, and leaf and chloroplast development. We also provide some gaps in our understanding of TOR function in photosynthesis that need to be addressed in the future.
1. The TOR Complex in Plants
During the long-term evolution from the Last Eukaryotic Common Ancestor (LECA), plants have diverged from animals and other eukaryotes in many ways. Their most prominent feature is the ability to using light energy to generate organic molecules (e.g., glucose (C6H12O6), sugars, starch) from carbon dioxide (CO2) and water (H2O) and release molecular oxygen (O2) into the atmosphere, which is also called photosynthesis [1]. Photosynthesis is an essential process and facilitates the expansion of plants broadly across all of the Earth’s biomes. With limited agricultural land and increasing human population, it is essential to decipher the key regulators involved in photosynthesis and enhance photosynthetic activities [1,2,3,4].
Living organisms possess many mechanisms to sense when nutrient availability and environmental cues are conducive to growth and development [5]. Through identification of different external or internal cues, organisms adjust their metabolism, growth, and development to survive harsh conditions. The target of rapamycin (TOR) kinase signaling cascade is a core component of nutrient sensing in all eukaryotic organisms. Recently, McCready et al. explained the importance of TOR kinase in plant development as a whole [6]. In addition, the role of the TOR signaling plants’ defense against pathogen and related metabolisms was studied recently [7]. When eukaryotes are in favorable conditions, TOR signaling pathway is active, which promotes anabolic processes and inhibits catabolism and protein degradation, but when conditions are unfavorable for growth, TOR is inactivated and promotes catabolic processes [8,9]. Xiong and Sheen explained the glucose-TOR signaling in the transcriptional control of the cell cycle and enumerated the importance of leaf photosynthesis-derived glucose in activating TOR [10].
Tremendous progress has been made in uncovering the molecular and cellular functions of TOR in regulating plant photosynthesis. In this paper, we first present a broad overview of recent developments in the plant TOR complex. Secondly, we highlight the impact of TOR signaling on plant photosynthesis, focusing on findings on the regulation of TOR in sensing C nutrient derived from photosynthesis, light absorptions, and leaf and chloroplast development. Finally, we present some pressing questions to be addressed as we move onward in the study of TOR signaling in plant photosynthesis.
Rapamycin (RAP) is an immunosuppressive macrolide antibiotic produced by the soil bacterium Streptomyces hygroscopicus on Easter Island, which stops yeast division and produces cells with an N starvation phenotype [11]. Through screening for genetic mutants resistant to rapamycin, FK506-binding protein 12 (FKBP12) together with TOR were first identified, and TOR emerged as a central regulatory hub in all eukaryotic organisms, including yeasts, animals, and plants [12,13,14,15,16]. TOR is a conserved serine/threonine-protein kinase and belongs to the phosphatidylinositol 3-kinase-related kinase family and has been found in all sequenced plant and algal genomes [11,17,18]. The TOR protein contains five conserved domains, including HEAT (Huntington, Elongation Factor 3 regulatory, subunit A of PP2A, TOR1) repeats, FAT (FRAP-ATM-TTRAP) domain, FRB (FKBP rapamycin-binding) domain, kinase domain and FATC domain (Figure 1). The HEAT repeats mediate protein–protein interactions and membrane associations; the FAT domain, together with the FATC domains, contributes to protein interactions and kinase activation; the FRB domain is targeted by the inhibitory FKBP12–rapamycin complex [19].
In yeast and mammalian systems, TOR exists in two structurally and functionally distinct protein complexes: the rapamycin-sensitive TOR complex 1 (TORC1) and the rapamycin-insensitive TOR complex 2 (TORC2) [20,21]. The core components of TORC1 include TOR, regulatory-associated protein of mTOR (RAPTOR)/KOG1, and lethal with SEC13 protein 8 (LST8), while TORC2 includes TOR, SAPK-interacting protein 1 (SIN1)/AVO1, rapamycin-insensitive companion of mTOR (RICTOR)/AVO3, and LST8. In mammalian systems, they have only one TOR gene. In yeast (Saccharomyces cerevisiae), two TOR genes, TOR1 and TOR2, were identified. Either TOR1 or TOR2 can form the TORC1 complex, while only TOR2 can form the TORC2 complex [8].
Research about TOR signaling in plants lags behind that in yeast and mammals. In plant systems examined to date, most of them have only one TOR gene; two TOR genes are identified in polyploids Glycine max, Populus trichocarpa, and Brassica rapa [17,22]. Four TOR genes are identified in allotetraploid cotton Gossypium hirsutum [23]. These results suggest that different plant species might have evolved unique TOR complexes with specific functions. Intriguingly, TORC2 complex components SIN1 and RICTOR appear to be absent in the plant lineage, indicating that plants may not form a conserved TORC2 [8,24].
Scientists revealed that plants were insensitive to rapamycin and could not form a complex with RAP, a small 12 kDa protein FKBP12 and the FRB domain of TOR [13,21,25]. However, the growth of Arabidopsis seedlings or cells in liquid culture was inhibited by RAP [26,27]. To elucidate the function of AtTOR in plants’ various cellular and developmental processes, rapamycin-sensitive transgenic A. thaliana lines (BP12) expressing yeast FK506 Binding Protein12 were developed and used [28,29,30,31,32,33]. In addition, a new generation of ATP-competitive chemical inhibitors specific to TOR kinase, such as AZD8055, Torin1, Torin2, and KU0063794, have been applied for TOR studies in plants [28,32,33].
In the model plant Arabidopsis thaliana, one copy of TOR gene, two copies of Raptor (RaptorA and RaptorB) genes, and two copies of LST8 (LST8-1 and LST8-2) genes exist [9,17]. Studies suggested that the null tor mutant is embryo-lethal, the raptora/b double mutant is unable to maintain post-embryonic meristem-driven growth, and the lst8-1 mutant exhibits modest dwarf growth and early senescence phenotypes [9,34,35,36]. These results revealed that Arabidopsis TOR, Raptor, and LST8 all play a pivotal role in regulating multiple plant growth and development, and TOR appears to regulate a much broader spectrum of biological functions than Raptor or LST8.
2. TOR: A Key Regulator in Photosynthesis
Extensive studies have demonstrated that TOR senses and integrates nutrient, hormone, light, energy, and other environmental cues to orchestrate growth and development. As a serine-threonine protein kinase, TOR mediates changes in protein phosphorylation and modulates a myriad of important cellular and metabolic processes, including nutrient assimilation, nutrient transport, ribosome biogenesis, cell division, cell expansion, polysome integrity, metabolism, autophagy, and development [14,17,31,37,38,39]. Photosynthesis functions as one of the most important anabolic processes in plants. Recent research suggest that TOR plays an indispensable role in plant photosynthesis [40]. Plants with TOR dysfunctions showed severe defects in chloroplasts and photosynthesis [23,36,41,42,43,44]. Transcriptome analysis in TOR-inhibited plants revealed a broad regulation of plant photosynthesis-related genes [28,30,41,45]. Intriguingly, activation of TOR leading to defects in chloroplast development compared with wild type, further supports the pivotal roles of TOR in regulating plant photosynthesis [46,47]. In the present review, we summarize recent advances that address the functions of TOR in regulating plant photoautotrophy and maintenance of photosynthesis (Figure 2).
2.1. Role of TOR in Sensing C Nutrient
Unlike yeast and animals, plants acquired the ability to live outside water, and their most prominent feature is probably their ability to synthesize organic carbon (C) molecules like glucose, sugars (fructose, glucose or sucrose) and starch by using light energy and releasing molecular oxygen into the atmosphere, which is also called photosynthesis. The ability to sense, assimilate, transport, and utilize C nutrients is vital for plant survival and growth. Plants decide whether to store sugars in source leaves or distribute these metabolites to sink organs, such as young organs, roots, or seeds to ensure the supply-demand balance of sugar in plants [48]. Evidence supports that TOR is involved in plant C nutrient sensing and serves as an intermediate for the regulation of plant growth by organic carbon (C) sugars [9,39].
At the photoautotrophic transition checkpoint, 15 mM glucose and sucrose can promote TOR signaling and control root meristem, root elongation, cotyledon expansion, and root hair production activation through glycolysis and mitochondrial bioenergetic. The requirement for glycolysis and mitochondrial bioenergetics to activate TOR protein kinase was monitored by the phosphorylation of its conserved substrates S6K. In this process, other sugars, such as fructose, galactose, and xylose, are much less effective [27,44]. Recent findings have illustrated that glucose is sufficient to activate TOR kinase in root apexes, whereas glucose and light signals are required for TOR activation in shoot apexes. In shoot apexes, exogenously auxin can replace light to activate TOR and promote leaf development. The small GTPase Rho-related protein 2 (ROP2) transduced light-auxin signal to activate TOR and promotes TOR-dependent phosphorylation and activation of transcription factor E2Fa/E2Fb, which eventually enhances root growth and leaf formation [27,44,49].
Glucose-activated TOR also controls the accumulation of the brassinosteroid (BR)-signaling transcription factor BZR1 (Brassinazole resistance 1). Inhibition of TOR by inducible RNAi reduced expression of BR-responsive genes and abolished sugar-promoted BZR1 accumulation. The growth inhibition caused by TOR inactivation can be partially recovered by exogenously applied BR and the BZR1 gain-of-function mutation bzr1-1D. The regulation of BZR1 accumulation by glucose-TOR allows carbon availability to control plant growth [50].
Through genetic approaches and chemical or physiological treatments to either promote or disrupt TOR activity, the results indicated that TOR restricted the plasmodesmata (PD) transport activity in leaves. Further study revealed that TOR was significantly more active in mature leaves photosynthesizing excess sugars (glucose) than in young and growing leaves [51].
In addition to glucose, sucrose is demonstrated to play a pivotal role in TOR activation and development of shoot apical meristem. Research suggested that sucrose was essential to promote leaf expansion even in the cop1 mutants, which exhibited constitutive photomorphogenesis with open cotyledons and a short hypocotyl. TOR protein plays a key role in integrating energy and light signaling to promote stem cell activation in the shoot apical meristem [52].
2.2. Light and TOR Signaling Pathway
Plants are oxygenic photosynthesizers that use light energy to generate organic molecules from carbon dioxide (CO2) and water (H2O). The ability to absorb the light energy has a significant effect on the photosynthetic efficiency of plants. In addition, light-stimulated photomorphogenesis is a developmental process that transforms plants into a vegetative state, which is required for photosynthetic activity. Light regulates the expression pattern of genes that participated in cell proliferation, organ development, and differentiation of photosynthetic cells. Advancing research has shown that light is intimately linked with TOR activation.
In Arabidopsis shoot apexes, both light signals and the glucose are indispensable for continuously promoting auxin accumulation for TOR activation to facilitate conversion to photoautotropic growth in Arabidopsis. Light-auxin signal can activate ROP2, which then promotes TOR kinase activity [49]. In deetiolating Arabidopsis seedlings, phytochrome A and cryptochromes perceive light to inactivate the negative regulator Constitutive photomorphogenesis 1 (COP1), and then leads to TOR activation and phosphorylation of downstream protein RPS6. The activation of TOR and RPS6 enhances cotyledon opening and transformation from young plant seedlings into the vegetative phase with photosynthetic activities [53]. This finding reveals the molecular mechanism of TOR activation in light-specific translational enhancement in plants de-etiolation.
In addition, the sugar requirement for TOR–S6K–RPS6 activation was investigated in older etiolated seedlings. RPS6 phosphorylation was elevated by light exposure for 2 h, but not for 0.5 h. However, when seedlings were grown with 1% sucrose, RPS6 phosphorylation was promoted by light as short as 0.5 h [54]. A recent study demonstrated that the impairment of TOR activity (through RAPTOR1B mutations or TOR kinase active site chemical inhibition) in Arabidopsis, reduced the accumulation of the photoreactive chlorophyll precursor protochlorophyllide in darkness, but increased greening rate of etiolated seedlings after exposure to light [55].
2.3. TOR Regulates Leaf and Chloroplast Development
Plant chloroplasts are essential organelles that carry out oxygenic photosynthesis. Several studies revealed the involvement of TOR signaling in the biogenesis and maturation of chloroplasts [23,29,33,45,48,55,56]. The suppression of TOR by active-site TOR inhibitor AZD8055 in Arabidopsis eliminated greening and cotyledon expansion and remodeled the expression profile of genes associated with photosynthesis, involved in chlorophyll biosynthesis, light reactions, and CO2 fixation [45]. The transcription and translation of nuclear-encoded mRNAs coding for plastidic ribosomal proteins were down-regulated in TOR inactivation plants (ethanol-inducible TOR RNAi lines), which could explain the chlorotic phenotype of the TOR silenced plants [48]. Recent findings also uncovered the genes involved in plastid biogenesis were particularly sensitive to TOR inhibition by AZD8055 treatment [57]. Analysis of the C. reinhardtii proteome and phosphoproteome under TOR inactivation revealed that the levels of most of the proteins related to the Calvin cycle were decreased, suggesting the involvement of TOR in photosynthesis [58].
Inhibition of TOR activity resulted in reduced chloroplast number and size, and thus impaired photoautotrophic growth in Arabidopsis [33]. In addition, scientists identified the brassinosteroid insensitive 2 (BIN2) as a novel phosphorylation target of TOR-S6K2 and uncovered the pivotal role of TOR-S6K2-BIN2 in heterotrophic to photoautotrophic transition [33]. In addition, TOR inhibition caused altered chloroplast morphology, increased non-photochemical quenching (NPQ), damage to photosystem II (PS II) reaction centers and inhibited efficient state transitions between PSII and PSI in alga [59]. In allotetraploid cotton Gossypium hirsutum, pharmacological experiments with TOR inhibitor AZD8055 and silencing GhTOR genes by virus-induced gene silencing (VIGS) resulted in a significant delay in the transition from heterotrophic to photoautotrophic growth [23].
Ectopic expression of full-length AtTOR increased plant photosynthetic efficiency and chlorophyll content in rice [60]. Type 2A phosphatase-associated protein of 46 kDa (TAP46), a regulatory subunit of protein phosphatase 2A, is a direct TORC1 phosphorylation substrate. Transgenic plants overexpressing TAP46 exhibited increased hypocotyl length and enlarged leaves [61,62]. Ribosomal protein S6 kinase (S6K) functions as a key component in the target of rapamycin (TOR) pathway. In rice (Oryza sativa L.), suppression of S6K1 caused pale yellow-green leaves and defective thylakoid grana architecture. Genetic and pharmacological evidence further demonstrated that S6K1 was vital for galactolipid biosynthesis for the thylakoid membrane [56]. Arabidopsis raptor1b mutation resulted in decreased CO2 assimilation rate and increased stomatal conductance; however, the chloroplast development or photosynthetic electron transport efficiency was not affected [42].
In Arabidopsis, TOR functioned as a positive regulator of chlorophyll biosynthesis and metabolism via TRIN1 (tor-inhibitor insensitive 1)/ABI4 (abscisic acid-insensitive 4) at the photoautotrophic stage. The precise molecular connection between TOR and ABI4 remains to be elucidated [29]. The TOR complex components lst8 and raptor mutants as well as wild-type Arabidopsis seedlings under TOR inhibition by AZD showed diminished ABA hormone accumulation. The diminished ABA hormone accumulation was correlated with decreased transcript levels of ABA biosynthetic enzymes including zeaxanthin epoxidase (ZEP), the 9-cis-epoxycarotenoid dioxygenase NCED3 and aldehyde oxidases AAO3, and increased transcript amount of the abscisic acid catabolism gene cytochrome P450 monooxygenases CYP707A2 [63]. Furthermore, an in vitro protein kinase screen identified that TOR kinase phosphorylated ABA receptors PYL at a conserved serine residue S119 [64]. These results suggested the reciprocal regulations between TOR and ABA signaling to balance plant growth and stress responses, consistent with the role of TOR and ABA in seedling development and greening of cotyledons and leaves [39].
Scientists revealed that inhibition of TOR increased the expression level of a nuclear-encoded chloroplast RelA-SpoT homolog (RSH) gene CmRSH4b. CmRSH4b encoded the guanosine 3′-diphosphate 5′-diphosphate (ppGpp) synthetases and regulated chloroplast rRNA transcription in the unicellular red alga Cyanidioschyzon merolae [65]. Researchers examined changes in oxidation during TOR inhibition in green alga Chlamydomonas reinhardtii. A reduction in photosynthetic electron flow and twenty proteins related with photosynthesis were identified with altered oxidation status under TOR inhibition [66].
3. Conclusions and Future Prospects
The analysis of the recent research progress and key findings on the TOR signaling pathways in plant photosynthesis suggests that TOR is a crucial signal integrator to transduce nutrient and environmental inputs into physiological, molecular, and developmental responses for growth. TOR kinase senses C nutrients and is activated by the availability of sugar produced by light-stimulated photosynthesis. Active TOR protein enhances plant photomorphogenesis and leaf development to make sure the maintenance of photosynthesis.
Compared with measurable progress made in the TOR signaling of mammalians and yeasts, our understanding of plant and algal TOR is limited. Although these findings in the present study clearly highlight a role for TOR in plant photosynthesis, several questions remain to be answered. Do plants possess unique TOR complex involved in plant specific life process photosynthesis? How do plants sense upstream signals for sugars and light? Plant hormones auxin, ABA and BR are involved in the regulation of TOR signaling and photosynthesis. What is the molecular mechanism of these hormones on TOR? How does TOR integrated into the known canonical hormone signaling? Are other hormones such as salicylic acid, jasmonic acid and ethylene involved in TOR regulation? In addition, these findings highlight a role for TOR in leaf and chloroplast development. What are the downstream effectors that mediate TOR function in this context?
In future studies, it is necessary to decipher the underpinning molecular mechanism of TOR in regulating plant photosynthesis. Genetic and biochemical analyses combined with currently available and future data from phosphoproteomic, transcriptomic and metabolomic are effective approaches to identify TOR functions and discover TOR up- and down-stream effectors.
With limited agricultural land and increasing human population, it is essential to enhance crop productivity. Increasing the photosynthesis rate of plants has been recently revitalized as one of the approaches for increasing grain crop yields and solving world food crises [1,2,3,4]. Understanding the regulation of TOR signaling in plants photosynthesis may be essential for crop improvement in the future.
Funding
This work is supported by grants from the National Natural Science Foundation of China (U1804231, 31972469, 31672206), State Key Laboratory of Cotton Biology Open Fund (CB2021A15), the Agricultural Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (34-IUA-02), the Sichuan Science and Technology Program (2020JDRC0044), and local financial funds of the National Agricultural Science & Technology Center (NASC), Chengdu (NASC2020AR08; NASC2021KR03).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, history and modelling. Ann. Bot. 2020, 126, 511–537. [Google Scholar] [CrossRef]
- Flexas, J.; Carriquí, M. Photosynthesis and photosynthetic efficiencies along the terrestrial plant’s phylogeny: Lessons for improving crop photosynthesis. Plant J. 2019, 101, 964–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundgren, M.R. C2 photosynthesis: A promising route towards crop improvement? New Phytol. 2020, 228, 1734–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, T.R.; Rufty, T.W.; Lewis, R.S. Increasing Photosynthesis: Unlikely Solution for World Food Problem. Trends Plant Sci. 2019, 24, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Velásquez, A.C.; Castroverde, C.D.; He, S.Y. Plant–Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCready, K.; Spencer, V.; Kim, M. The Importance of TOR Kinase in Plant Development. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Filipe, O.; Hoffman, G.; Seifi, H.S.; Haeck, A.; Canlas, P.; Van Bockhaven, J.; De Waele, E.; Demeestere, K.; Ronald, P.; et al. Target of rapamycin signaling orchestrates growth–defense trade-offs in plants. New Phytol. 2017, 217, 305–319. [Google Scholar] [CrossRef] [Green Version]
- Burkart, G.M.; Brandizzi, F. A Tour of TOR Complex Signaling in Plants. Trends Biochem. Sci. 2021, 46, 417–428. [Google Scholar] [CrossRef]
- Ingargiola, C.; Duarte, G.T.; Robaglia, C.; Leprince, A.-S.; Meyer, C. The Plant Target of Rapamycin: A Conduc TOR of Nutrition and Metabolism in Photosynthetic Organisms. Genes 2020, 11, 1285. [Google Scholar] [CrossRef]
- Xiong, Y.; Sheen, J. Moving beyond translation: Glucose-TOR signaling in the transcriptional control of cell cycle. Cell Cycle 2013, 12, 1989–1990. [Google Scholar] [CrossRef] [Green Version]
- Dobrenel, T.; Caldana, C.; Hanson, J.; Robaglia, C.; Vincentz, M.; Veit, B.; Meyer, C. TOR Signaling and Nutrient Sensing. Annu. Rev. Plant Biol. 2016, 67, 261–285. [Google Scholar] [CrossRef] [PubMed]
- Heitman, J.; Movva, N.R.; Hiestand, P.C.; Hall, M.N. FK 506-binding protein proline rotamase is a target for the immunosuppressive agent FK 506 in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1991, 88, 1948–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menand, B.; Desnos, T.; Nussaume, L.; Berger, F.; Bouchez, D.; Meyer, C.; Robaglia, C. Expression and disruption of the Arabidopsis TOR (target of rapamycin) gene. Proc. Natl. Acad. Sci. USA 2002, 99, 6422–6427. [Google Scholar] [CrossRef] [Green Version]
- A Ryabova, L.; Robaglia, C.; Meyer, C. Target of Rapamycin kinase: Central regulatory hub for plant growth and metabolism. J. Exp. Bot. 2019, 70, 2211–2216. [Google Scholar] [CrossRef]
- Sabatini, D.M.; Erdjument-Bromage, H.; Lui, M.; Tempst, P.; Snyder, S.H. RAFT1: A mammalian protein that binds to FKBP12 in a rapamycin-dependent fashion and is homologous to yeast TORs. Cell 1994, 78, 35–43. [Google Scholar] [CrossRef]
- Xiong, Y.; Sheen, J. The Role of Target of Rapamycin Signaling Networks in Plant Growth and Metabolism. Plant Physiol. 2014, 164, 499–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Wang, P.; Xiong, Y. Target of Rapamycin Signaling in Plant Stress Responses. Plant Physiol. 2020, 182, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- John, F.; Roffler, S.; Wicker, T.; Ringli, C. Plant TOR signaling components. Plant Signal. Behav. 2011, 6, 1700–1705. [Google Scholar] [CrossRef] [Green Version]
- Rexin, D.; Meyer, C.; Robaglia, C.; Veit, B. TOR signalling in plants. Biochem. J. 2015, 470, 1–14. [Google Scholar] [CrossRef]
- Russell, R.; Fang, C.; Guan, K.-L. An emerging role for TOR signaling in mammalian tissue and stem cell physiology. Development 2011, 138, 3343–3356. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Um, J.-H.; Kim, S.; Kim, Y.-K.; Song, S.-B.; Lee, S.-H.; Verma, D.P.S.; Cheon, C.-I. RNA interference-mediated repression of S6 kinase 1 impairs root nodule development in soybean. Mol. Cells 2013, 35, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, L.; Yang, Z.; Zhao, G.; Zhang, X.; Wang, L.; Zheng, L.; Zhuo, F.; Yin, H.; Ge, X.; et al. Target of Rapamycin (TOR) Regulates the Expression of lncRNAs in Response to Abiotic Stresses in Cotton. Front. Genet. 2019, 9, 690. [Google Scholar] [CrossRef] [Green Version]
- Tatebe, H.; Shiozaki, K. Evolutionary Conservation of the Components in the TOR Signaling Pathways. Biomolecules 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Mahfouz, M.M.; Kim, S.; Delauney, A.J.; Verma, D.P.S. Arabidopsis TARGET OF RAPAMYCIN Interacts with RAPTOR, Which Regulates the Activity of S6 Kinase in Response to Osmotic Stress Signals. Plant Cell 2005, 18, 477–490. [Google Scholar] [CrossRef] [Green Version]
- Van Leene, J.; Han, C.; Gadeyne, A.; Eeckhout, D.; Matthijs, C.; Cannoot, B.; De Winne, N.; Persiau, G.; Van De Slijke, E.; Van De Cotte, B.; et al. Capturing the phosphorylation and protein interaction landscape of the plant TOR kinase. Nat. Plants 2019, 5, 316–327. [Google Scholar] [CrossRef]
- Xiong, Y.; Sheen, J. Rapamycin and Glucose-Target of Rapamycin (TOR) Protein Signaling in Plants. J. Biol. Chem. 2012, 287, 2836–2842. [Google Scholar] [CrossRef] [Green Version]
- Deng, K.; Dong, P.; Wang, W.; Feng, L.; Xiong, F.; Wang, K.; Zhang, S.; Feng, S.; Wang, B.; Zhang, J.; et al. The TOR Pathway Is Involved in Adventitious Root Formation in Arabidopsis and Potato. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Song, Y.; Wang, K.; Dong, P.; Zhang, X.; Li, F.; Li, Z.; Ren, M. TOR-inhibitor insensitive-1 (TRIN1) regulates cotyledons greening in Arabidopsis. Front. Plant Sci. 2015, 6, 861. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.; Venglat, P.; Qiu, S.; Feng, L.; Cao, Y.; Wang, E.; Xiang, D.; Wang, J.; Alexander, D.; Chalivendra, S.; et al. Target of Rapamycin Signaling Regulates Metabolism, Growth, and Life Span in Arabidopsis. Plant Cell 2012, 24, 4850–4874. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Wu, Y.; Sheen, J. TOR signaling in plants: Conservation and innovation. Development 2018, 145. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Zhao, G.; Zhang, X.; Li, L.; Xiong, F.; Zhuo, F.; Zhang, C.; Yang, Z.; Datla, R.; Ren, M.; et al. The crosstalk between Target of Rapamycin (TOR) and Jasmonic Acid (JA) signaling existing in Arabidopsis and cotton. Sci. Rep. 2017, 7, srep45830. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Zhang, R.; Meng, Z.; Deng, K.; Que, Y.; Zhuo, F.; Feng, L.; Guo, S.; Datla, R.; Ren, M. Brassinosteriod Insensitive 2 (BIN2) acts as a downstream effector of the Target of Rapamycin (TOR) signaling pathway to regulate photoautotrophic growth in Arabidopsis. New Phytol. 2016, 213, 233–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, G.H.; Veit, B.; Hanson, M.R. The Arabidopsis AtRaptor genes are essential for post-embryonic plant growth. BMC Biol. 2005, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreau, M.; Azzopardi, M.; Clément, G.; Dobrenel, T.; Marchive, C.; Renne, C.; Martin-Magniette, M.L.; Taconnat, L.; Renou, J.P.; Robaglia, C.; et al. Mutations in the Arabidopsis homolog of LST8/GβL, a partner of the target of Rapamycin kinase, impair plant growth, flowering, and metabolic adaptation to long days. Plant Cell 2012, 24, 463–481. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.; Qiu, S.; Venglat, P.; Xiang, D.; Feng, L.; Selvaraj, G.; Datla, R. Target of Rapamycin Regulates Development and Ribosomal RNA Expression through Kinase Domain in Arabidopsis. Plant Physiol. 2011, 155, 1367–1382. [Google Scholar] [CrossRef] [Green Version]
- Jamsheer, K.M.; Jindal, S.; Laxmi, A. Evolution of TOR-SnRK dynamics in green plants and its integration with phytohormone signaling networks. J. Exp. Bot. 2019, 70, 2239–2259. [Google Scholar] [CrossRef]
- Schepetilnikov, M.; Ryabova, L.A. Recent Discoveries on the Role of TOR (Target of Rapamycin) Signaling in Translation in Plants. Plant Physiol. 2017, 176, 1095–1105. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, L.; Li, L.; Fu, L.; Liu, Y.; Xiong, Y.; Sheen, J. Integration of nutrient, energy, light, and hormone signalling via TOR in plants. J. Exp. Bot. 2019, 70, 2227–2238. [Google Scholar] [CrossRef]
- Quilichini, T.D.; Gao, P.; Pandey, P.K.; Xiang, D.; Ren, M.; Datla, R. A role for TOR signaling at every stage of plant life. J. Exp. Bot. 2019, 70, 2285–2296. [Google Scholar] [CrossRef]
- Caldana, C.; Li, Y.; Leisse, A.; Zhang, Y.; Bartholomaeus, L.; Fernie, A.R.; Willmitzer, L.; Giavalisco, P. Systemic analysis of inducible target of rapamycin mutants reveals a general metabolic switch controlling growth in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2013, 73, 897–909. [Google Scholar] [CrossRef]
- Salem, M.A.; Li, Y.; Bajdzienko, K.; Fisahn, J.; Watanabe, M.; Hoefgen, R.; Schöttler, M.A.; Giavalisco, P. RAPTOR Controls Developmental Growth Transitions by Altering the Hormonal and Metabolic Balance. Plant Physiol. 2018, 177, 565–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepetilnikov, M.; Dimitrova, M.; Mancera-Martínez, E.; Geldreich, A.; Keller, M.; Ryabova, L.A. TOR and S6K1 promote translation reinitiation of uORF-containing mRNAs via phosphorylation of eIF3h. EMBO J. 2013, 32, 1087–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; McCormack, M.P.; Li, L.; Hall, Q.; Xiang, C.; Sheen, J. Glucose–TOR signalling reprograms the transcriptome and activates meristems. Nat. Cell Biol. 2013, 496, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, P.; Xiong, F.; Que, Y.; Wang, K.; Yu, L.; Li, Z.; Ren, M. Expression profiling and functional analysis reveals that TOR is a key player in regulating photosynthesis and phytohormone signaling pathways in Arabidopsis. Front. Plant Sci. 2015, 6, 677. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.; Kim, S.-J.; Xing, A.; A Schenck, C.; Liu, L.; Jiang, N.; Wang, J.; Last, R.L.; Brandizzi, F. Homeostasis of branched-chain amino acids is critical for the activity of TOR signaling in Arabidopsis. eLife 2019, 8. [Google Scholar] [CrossRef]
- Schaufelberger, M.; Galbier, F.; Herger, A.; Francisco, R.D.B.; Roffler, S.; Clement, G.; Diet, A.; Hörtensteiner, S.; Wicker, T.; Ringli, C. Mutations in the Arabidopsis ROL17/isopropylmalate synthase 1 locus alter amino acid content, modify the TOR network, and suppress the root hair cell development mutant lrx1. J. Exp. Bot. 2019, 70, 2313–2323. [Google Scholar] [CrossRef]
- Dobrenel, T.; Mancera-Martínez, E.; Forzani, C.; Azzopardi, M.; Davanture, M.; Moreau, M.; Schepetilnikov, M.; Chicher, J.; Langella, O.; Zivy, M.; et al. The Arabidopsis TOR Kinase Specifically Regulates the Expression of Nuclear Genes Coding for Plastidic Ribosomal Proteins and the Phosphorylation of the Cytosolic Ribosomal Protein S6. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Cai, W.; Liu, Y.; Li, H.; Fu, L.; Liu, Z.; Xu, L.; Liu, H.; Xu, T.; Xiong, Y. Differential TOR activation and cell proliferation in Arabidopsis root and shoot apexes. Proc. Natl. Acad. Sci. USA 2017, 114, 2765–2770. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, J.-Y.; Roh, J.; Marchive, C.; Kim, S.-K.; Meyer, C.; Sun, Y.; Wang, W.; Wang, Z.-Y. TOR Signaling Promotes Accumulation of BZR1 to Balance Growth with Carbon Availability in Arabidopsis. Curr. Biol. 2016, 26, 1854–1860. [Google Scholar] [CrossRef]
- Brunkard, J.O.; Xu, M.; Scarpin, M.R.; Chatterjee, S.; Shemyakina, E.A.; Goodman, H.M.; Zambryski, P. TOR dynamically regulates plant cell–cell transport. Proc. Natl. Acad. Sci. USA 2020, 117, 5049–5058. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, A.; Janocha, D.; Dong, Y.; Medzihradszky, A.; Schöne, S.; Daum, G.; Suzaki, T.; Forner, J.; Langenecker, T.; Rempel, E.; et al. Integration of light and metabolic signals for stem cell activation at the shoot apical meristem. eLife 2016, 5, e17023. [Google Scholar] [CrossRef]
- Chen, G.-H.; Liu, M.-J.; Xiong, Y.; Sheen, J.; Wu, S.-H. TOR and RPS6 transmit light signals to enhance protein translation in deetiolating Arabidopsis seedlings. Proc. Natl. Acad. Sci. USA 2018, 115, 12823–12828. [Google Scholar] [CrossRef] [Green Version]
- Enganti, R.; Cho, S.K.; Toperzer, J.D.; Camacho, R.U.; Cakir, O.S.; Ray, A.P.; Abraham, P.E.; Hettich, R.L.; Von Arnim, A.G. Phosphorylation of Ribosomal Protein RPS6 Integrates Light Signals and Circadian Clock Signals. Front. Plant Sci. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, Y.; McFarlane, H.E.; Obata, T.; Richter, A.S.; Lohse, M.; Grimm, B.; Persson, S.; Fernie, A.R.; Giavalisco, P. Inhibition of TOR Represses Nutrient Consumption, Which Improves Greening after Extended Periods of Etiolation. Plant Physiol. 2018, 178, 101–117. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yu, Y.; Hu, W.; Min, Q.; Kang, H.; Li, Y.; Hong, Y.; Wang, X.; Hong, Y. Ribosomal protein S6 kinase1 coordinates with TOR-Raptor2 to regulate thylakoid membrane biosynthesis in rice. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2016, 1861, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, B.; Bilooei, S.F.; Dóczi, R.; Grove, E.; Railo, S.; Palme, K.; Ditengou, F.A.; Bögre, L.; López-Juez, E. Converging Light, Energy and Hormonal Signaling Control Meristem Activity, Leaf Initiation, and Growth. Plant Physiol. 2017, 176, 1365–1381. [Google Scholar] [CrossRef] [Green Version]
- Roustan, V.; Weckwerth, W. Quantitative Phosphoproteomic and System-Level Analysis of TOR Inhibition Unravel Distinct Organellar Acclimation in Chlamydomonas reinhardtii. Front. Plant Sci. 2018, 9, 1590. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, S.; Rao, B.J. Reciprocal regulation of photosynthesis and mitochondrial respiration by TOR kinase in Chlamydomonas reinhardtii. Plant Direct 2019, 3, e00184. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, A.; Moin, M.; Kumar, M.U.; Reddy, A.B.M.; Ren, M.; Datla, R.; Siddiq, E.A.; Kirti, P.B. Ectopic expression of Arabidopsis Target of Rapamycin (AtTOR) improves water-use efficiency and yield potential in rice. Sci. Rep. 2017, 7, 42835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, C.S.; Ahn, H.-K.; Pai, H.-S. Overexpression of the PP2A regulatory subunit Tap46 leads to enhanced plant growth through stimulation of the TOR signalling pathway. J. Exp. Bot. 2014, 66, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, C.S.; Han, J.-A.; Lee, H.-S.; Lee, S.; Pai, H.-S. The PP2A Regulatory Subunit Tap46, a Component of the TOR Signaling Pathway, Modulates Growth and Metabolism in Plants. Plant Cell 2011, 23, 185–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kravchenko, A.; Citerne, S.; Jéhanno, I.; Bersimbaev, R.I.; Veit, B.; Meyer, C.; Leprince, A.-S. Mutations in the Arabidopsis Lst8 and Raptor genes encoding partners of the TOR complex, or inhibition of TOR activity decrease abscisic acid (ABA) synthesis. Biochem. Biophys. Res. Commun. 2015, 467, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, Y.; Li, Z.; Hsu, C.-C.; Liu, X.; Fu, L.; Hou, Y.-J.; Du, Y.; Xie, S.; Zhang, C.; et al. Reciprocal Regulation of the TOR Kinase and ABA Receptor Balances Plant Growth and Stress Response. Mol. Cell 2017, 69, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, S.; Nomura, Y.; Takemura, T.; Pancha, I.; Taki, K.; Toguchi, K.; Tozawa, Y.; Tanaka, K. The checkpoint kinase TOR (target of rapamycin) regulates expression of a nuclear-encoded chloroplast RelA-SpoT homolog ( RSH ) and modulates chloroplast ribosomal RNA synthesis in a unicellular red alga. Plant J. 2018, 94, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Ford, M.M.; Smythers, A.L.; McConnell, E.W.; Lowery, S.C.; Kolling, D.R.J.; Hicks, L.M. Inhibition of TOR in Chlamydomonas reinhardtii Leads to Rapid Cysteine Oxidation Reflecting Sustained Physiological Changes. Cells 2019, 8, 1171. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Structure of TOR protein and strategies to dissect TOR function. The TOR protein contains five conserved domains including HEAT repeats, FAT domain, FRB domain, kinase domain and FATC domain. The multifaceted functions of TOR kinase are dissected by integrated analysis of chemical inhibitor treatments, TOR RNAi seedlings and TOR signaling-related mutants. Abbreviations: TOR, Target of Rapamycin; HEAT repeats, Huntingtin, Elongation factor 3, subunit of protein phosphatase 2A and TOR1; FAT, FRAP, ATM and TRRAP domain; FRB, FKP12-rapamycin binding domain; FATC, Carboxy-terminal FAT domain; RAPTOR, regulatory-associated protein of mTOR; LST8, lethal with Sec13 protein 8; FKBP12, FK506-binding protein 12; RNAi, RNA interference; amiRNA, artificial microRNA.
Figure 1.
Structure of TOR protein and strategies to dissect TOR function. The TOR protein contains five conserved domains including HEAT repeats, FAT domain, FRB domain, kinase domain and FATC domain. The multifaceted functions of TOR kinase are dissected by integrated analysis of chemical inhibitor treatments, TOR RNAi seedlings and TOR signaling-related mutants. Abbreviations: TOR, Target of Rapamycin; HEAT repeats, Huntingtin, Elongation factor 3, subunit of protein phosphatase 2A and TOR1; FAT, FRAP, ATM and TRRAP domain; FRB, FKP12-rapamycin binding domain; FATC, Carboxy-terminal FAT domain; RAPTOR, regulatory-associated protein of mTOR; LST8, lethal with Sec13 protein 8; FKBP12, FK506-binding protein 12; RNAi, RNA interference; amiRNA, artificial microRNA.

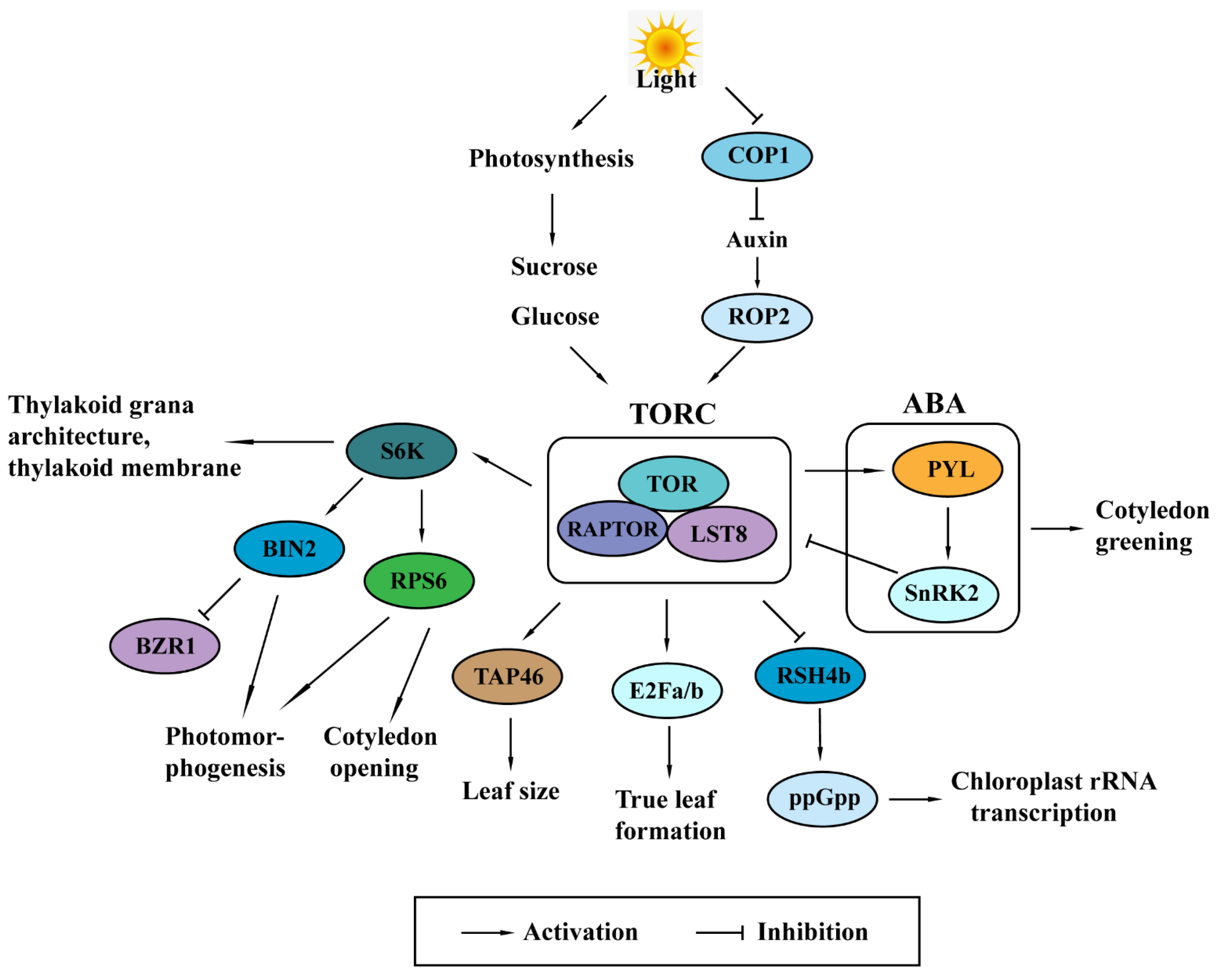
Figure 2.
Summary of Target of Rapamycin (TOR) signaling in plant photosynthesis, including inputs for the activation of TOR signaling and output processes regulated by the TOR signaling. Light signals and sugars derived from photosynthesis activate TOR signaling and the conserved downstream outputs. The activated TOR downstream outputs regulate photomorphogenesis and leaf development, which eventually influences the efficiency of oxygenic photosynthesis. Abbreviations: COP1, constitutive photomorphogenesis 1; ROP2, Rho-like small GTPase 2; TORC, Target of Rapamycin complex; TOR, Target of Rapamycin; RAPTOR, regulatory-associated protein of mTOR; LST8, lethal with Sec13 protein 8; S6K, S6 kinase; BIN2, brassinosteroid-insensitive 2; BZR1, brassinazole resistance 1; RPS6, ribosomal protein S6; TAP46, type 2A-phosphatase-associated protein 46 kDa; E2Fa/b, E2 promoter-binding factor a/b; RSH4b, RelA-SpoT homolog gene 4b; ppGpp, guanosine 3′-diphosphate 5′-diphosphate; PYL, pyrabactin resistance 1-like; SnRK2, SNF1-related protein kinase 2.
Figure 2.
Summary of Target of Rapamycin (TOR) signaling in plant photosynthesis, including inputs for the activation of TOR signaling and output processes regulated by the TOR signaling. Light signals and sugars derived from photosynthesis activate TOR signaling and the conserved downstream outputs. The activated TOR downstream outputs regulate photomorphogenesis and leaf development, which eventually influences the efficiency of oxygenic photosynthesis. Abbreviations: COP1, constitutive photomorphogenesis 1; ROP2, Rho-like small GTPase 2; TORC, Target of Rapamycin complex; TOR, Target of Rapamycin; RAPTOR, regulatory-associated protein of mTOR; LST8, lethal with Sec13 protein 8; S6K, S6 kinase; BIN2, brassinosteroid-insensitive 2; BZR1, brassinazole resistance 1; RPS6, ribosomal protein S6; TAP46, type 2A-phosphatase-associated protein 46 kDa; E2Fa/b, E2 promoter-binding factor a/b; RSH4b, RelA-SpoT homolog gene 4b; ppGpp, guanosine 3′-diphosphate 5′-diphosphate; PYL, pyrabactin resistance 1-like; SnRK2, SNF1-related protein kinase 2.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Song, Y.; Alyafei, M.S.; Masmoudi, K.; Jaleel, A.; Ren, M. Contributions of TOR Signaling on Photosynthesis. Int. J. Mol. Sci. 2021, 22, 8959. https://doi.org/10.3390/ijms22168959
AMA Style
Song Y, Alyafei MS, Masmoudi K, Jaleel A, Ren M. Contributions of TOR Signaling on Photosynthesis. International Journal of Molecular Sciences. 2021; 22(16):8959. https://doi.org/10.3390/ijms22168959
Chicago/Turabian StyleSong, Yun, Mohammed Salem Alyafei, Khaled Masmoudi, Abdul Jaleel, and Maozhi Ren. 2021. "Contributions of TOR Signaling on Photosynthesis" International Journal of Molecular Sciences 22, no. 16: 8959. https://doi.org/10.3390/ijms22168959
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.