Increased Cardiovascular Risk Associated with Chemical Sensitivity to Perfluoro–Octanoic Acid: Role of Impaired Platelet Aggregation

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
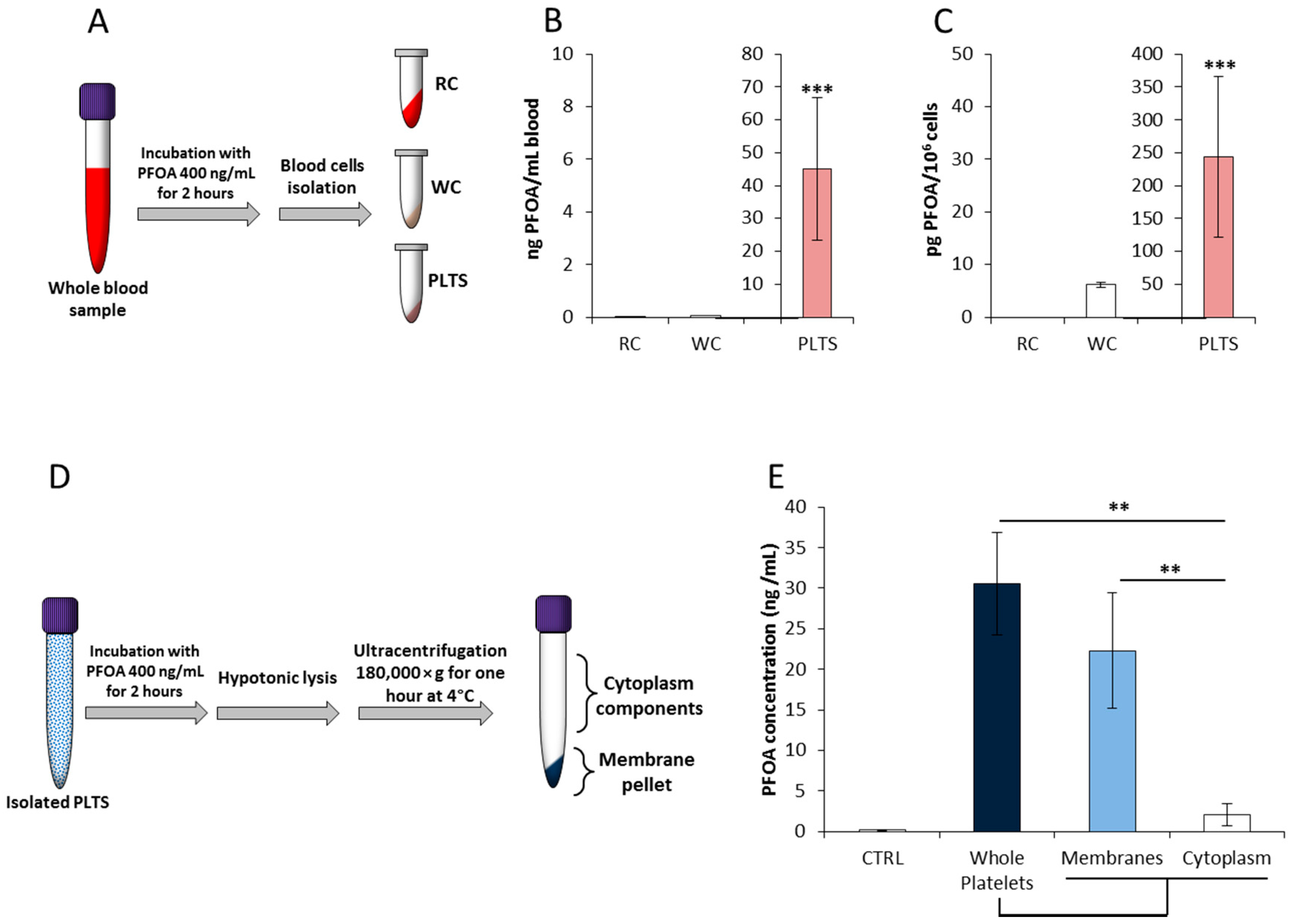
2.1. Platelets’ Membrane is the Major Cell-Accumulation Site of Blood PFOA
2.2. PFOA Accumulation Alters Platelets’ Membrane Stability at Docking Analysis and the Aggregation Process In Vitro
2.3. Environmental Exposure to PFOA Associates with Hypercoagulability Profile
3. Discussion
4. Materials and Methods
4.1. Platelets’ Isolation for Flow Cytometry and Immunofluorescence Studies
4.2. Liquid Chromatography-Mass Spectrometry
4.3. Docking and Membrane Modelling
4.4. Flow Cytometry Analysis
4.5. Analysis of Platelet Calcium Flux
4.6. Study Population
4.7. Multiplate® Platelet Aggregation Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASPI | Arachidonic acid |
| AUC | Area under the curve |
| CVD | Cardiovascular diseases |
| ED | endocrine disruptors |
| LC/MS/MS | liquid chromatography-mass spectrometry |
| LOQ | limit of quantification |
| MC540 | Merocyanine 540 |
| PC | Phosphatidylcholine |
| PFAS | Perfluoro–alkyl substances; |
| PFOA | Perfluoro–octanoic acid |
| PFOS | Perfluoro–octane–sulfonate |
| TRAP-6 | Thrombin receptor activator peptide 6 |
| ADP | Adenosine diphosphate |
References
- Conder, J.M.; Hoke, R.A.; De Wolf, W.; Russell, M.H.; Buck, R.C. Are PFCAs bioaccumulative? A critical review and comparison with regulatory criteria and persistent lipophilic compounds. Environ. Sci. Technol. 2008, 42, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Anitole, K.; Hodes, C.; Lai, D.; Pfahles-Hutchens, A.; Seed, J. Perfluoroalkyl acids: A review of monitoring and toxicological findings. Toxicol. Sci. 2007, 99, 366–394. [Google Scholar] [CrossRef] [Green Version]
- Steenland, K.; Fletcher, T.; Savitz, D.A. Epidemiologic evidence on the health effects of perfluorooctanoic acid (PFOA). Environ. Health Perspect. 2010, 118, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, A.; Sabovic, I.; Valente, U.; Tescari, S.; Rocca, M.S.; Guidolin, D.; Dall’Acqua, S.; Acquasaliente, L.; Pozzi, N.; Plebani, M.; et al. Endocrine Disruption of Androgenic Activity by Perfluoroalkyl Substances: Clinical and Experimental Evidence. J. Clin. Endocrinol. Metab. 2019, 104, 1259–1271. [Google Scholar] [CrossRef] [Green Version]
- Olsen, G.W.; Burris, J.M.; Ehresman, D.J.; Froehlich, J.W.; Seacat, A.M.; Butenhoff, J.L.; Zobel, L.R. Half-life of serum elimination of perfluorooctanesulfonate, perfluorohexanesulfonate, and perfluorooctanoate in retired fluorochemical production workers. Env. Health Perspect. 2007, 115, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- IARC Monographs Volume 110: Perfluorooctanoic Acid, Tetrafluoroethylene, Dichloromethane, 1,2-Dichloropropane, and 1,3-Propane Sultone. IARC Publications 20 July 2016. Available online: https://publications.iarc.fr/547 (accessed on 8 January 2020).
- Roth, G.A.; Huffman, M.D.; Moran, A.E.; Feigin, V.; Mensah, G.A.; Naghavi, M.; Murray, C.J. Global and regional patterns in cardiovascular mortality from 1990 to 2013. Circulation 2015, 132, 1667–1678. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, A. Environmental cardiology: Studying mechanistic links between pollution and heart disease. Circ. Res. 2006, 99, 692–705. [Google Scholar] [CrossRef] [Green Version]
- Mastin, J.P. Environmental cardiovascular disease. Cardiovasc. Toxicol. 2005, 5, 91–94. [Google Scholar] [CrossRef]
- Spiezia, L.; Campello, E.; Maggiolo, S.; Tormene, D.; Simioni, P. Short-term exposure to high levels of air pollution (nickel) and the risk of acute unprovoked proximal deep vein thrombosis in the legs. Intern. Emerg. Med. 2016, 11, 159–162. [Google Scholar] [CrossRef]
- Geiger, S.D.; Xiao, J.; Ducatman, A.; Frisbee, S.; Innes, K.; Shankar, A. The association between PFOA, PFOS and serum lipid levels in adolescents. Chemosphere 2014, 98, 78–83. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chen, P.C.; Lin, Y.C.; Lin, L.Y. Association among serum perfluoroalkyl chemicals, glucose homeostasis, and metabolic syndrome in adolescents and adults. Diabetes. Care 2009, 32, 702–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, J.W.; Hatch, E.E.; Webster, T.F. Exposure to polyfluoroalkyl chemicals and cholesterol, body weight, and insulin resistance in the general U.S. population. Environ. Health Perspect. 2010, 118, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, N.M.; Fenton, S.E.; Strynar, M.; Hines, E.P.; Pritchard, D.A.; Steiner, A.Z. Effects of perfluorinated chemicals on thyroid function, markers of ovarian reserve, and natural fertility. Reprod. Toxicol. 2017, 69, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.S.; Wen, L.L.; Chu, P.L.; Lin, C.Y. Association among total serum isomers of perfluorinated chemicals, glucose homeostasis, lipid profiles, serum protein and metabolic syndrome in adults: NHANES, 2013–2014. Environ. Pollut. 2018, 232, 73–79. [Google Scholar] [CrossRef]
- Shankar, A.; Xiao, J.; Ducatman, A. Perfluorooctanoic Acid and Cardiovascular Disease in US Adults. Arch. Intern. Med. 2012, 172, 1397. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Jiao, J.; Zhuang, P.; Chen, X.; Wang, J.; Zhang, Y. Serum polyfluoroalkyl chemicals are associated with risk of cardiovascular diseases in national US population. Environ. Int. 2018, 119, 37–46. [Google Scholar] [CrossRef]
- Anderson-Mahoney, P.; Kotlerman, J.; Takhar, H.; Gray, D.; Dahlgren, J. Self-reported health effects among community residents exposed to perfluorooctanoate. New Solut. 2008, 18, 129–143. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lin, L.Y.; Wen, T.W.; Lien, G.W.; Chien, K.L.; Hsu, S.H.; Liao, C.C.; Sung, F.C.; Chen, P.C.; Su, T.C. Association between levels of serum perfluorooctane sulfate and carotid artery intima-media thickness in adolescents and young adults. Int. J. Cardiol. 2013, 168, 3309–3316. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chen, P.C.; Lo, S.C.; Torng, P.L.; Sung, F.C.; Su, T.C. The association of carotid intima-media thickness with serum Level of perfluorinated chemicals and endothelium-platelet microparticles in adolescents and young adults. Environ. Int. 2016, 94, 292–299. [Google Scholar] [CrossRef]
- Lebas, H.; Yahiaoui, K.; Martos, R.; Boulaftali, Y. Platelets Are at the Nexus of Vascular Diseases. Front. Cardiovasc. Med. 2019, 6, 132. [Google Scholar] [CrossRef]
- Willoughby, S.; Holmes, A.; Loscalzo, J. Platelets and cardiovascular disease. Eur. J. Cardiovasc. Nurs. 2002, 1, 273–288. [Google Scholar] [CrossRef]
- Puurunen, M.K.; Hwang, S.J.; Larson, M.G.; Vasan, R.S.; O’Donnell, C.J.; Tofler, G.; Johnson, A.D. ADP Platelet Hyperreactivity Predicts Cardiovascular Disease in the FHS (Framingham Heart Study). J. Am. Heart Assoc. 2018. [Google Scholar] [CrossRef] [PubMed]
- Argacha, J.F.; Mizukami, T.; Bourdrel, T.; Bind, M.A. Ecology of the cardiovascular system: Part II—A focus on non-air related pollutants. Trends Cardiovasc. Med. 2019, 29, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Whiting, D.; Di Nardo, J.A. TEG and ROTEM: Technology and clinical applications. Am. J. Hematol. 2014, 89, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Ehresman, D.J.; Froehlich, J.W.; Olsen, G.W.; Chang, S.-C.; Butenhoff, J.L. Comparison of human whole blood, plasma, and serum matrices for the determination of perfluorooctanesulfonate (PFOA), and other fluorochemicals. Environ. Res. 2007, 103, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Butenhoff, J.L.; Pieterman, E.; Ehresman, D.J.; Gorman, G.S.; Olsen, G.W.; Chang, S.C.; Princen, H.M. Distribution of perfluorooctanesulfonate and perfluorooctanoate into human plasma lipoprotein fractions. Toxicol. Lett. 2012, 210, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Nouhi, S.; Ahrens, L.; Campos Pereira, H.; Hughes, A.V.; Campana, M.; Gutfreund, P. Interactions of perfluoroalkyl substances with a phospholipid bilayer studied by neutron reflectometry. J. Colloid Interface Sci. 2018, 511, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, N.J.M.; Wargenau, A.; Sorenson, C.; Pedersen, J.; Tufenkji, N.; Novak, P.J.; Simcik, M.F. Partitioning and Accumulation of Perfluoroalkyl Substances in Model Lipid Bilayers and Bacteria. Environ. Sci. Technol. 2018, 52, 10433–10440. [Google Scholar] [CrossRef]
- Šabović, I.; Cosci, I.; De Toni, L.; Ferramosca, A.; Stornaiuolo, M.; Di Nisio, A.; Dall’Acqua, S.; Garolla, A.; Foresta, A. Perfluoro-octanoic acid impairs sperm motility through the alteration of plasma membrane. J. Endocrinol. Investig. 2019. [Google Scholar] [CrossRef]
- Birò, E.; Akkerman, J.W.N.; Hoek, F.J.; Gorter, G.; Pronk, L.M.; Sturk, A.; Nieuwland, R. The phospholipid composition and the cholesterol content of platelet-derived microparticles: A comparison with platelet membrane fractions. J. Thromb. Haemost. 2005, 3, 2754–2763. [Google Scholar] [CrossRef]
- Buffone, M.G.; Brugo-Olmedo, S.; Calamera, J.C.; Verstraeten, S.V.; Urrutia, F.; Grippo, L.; Corbetta, J.P.; Doncel, G.F. Decreased protein tyrosine phosphorylation and membrane fluidity in spermatozoa from infertile men with varicocele. Mol. Reprod. Dev. 2006, 73, 1591–1599. [Google Scholar] [CrossRef] [PubMed]
- Williamson, P.; Mattocks, K.; Schlegal, R.A. Merocyanine 540, a fluorescent probe sensitive to lipid packaging. Biochim. Biophys. Acta 1983, 732, 387–393. [Google Scholar] [CrossRef]
- Langner, M.; Hui, S.W. Merocyanine interaction with phosphatidylcholine bilayers. Biochim. Biophys. Acta 1993, 1149, 175–179. [Google Scholar] [CrossRef]
- Rathi, R.; Colenbrander, B.; Bevers, M.M.; Gadella, B.M. Evaluation of in vitro capacitation of stallion spermatozoa. Biol. Reprod. 2001, 65, 462–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavodnik, I.B.; Lapshina, E.A.; Palecz, D.; Bryszewska, M. The effects of palmitate on human erythrocyte membrane potential and osmotic stability. Scand. J. Clin. Lab. Investig. 1996, 56, 401–407. [Google Scholar] [CrossRef]
- Tillman, T.S.; Cascio, M. Effects of membrane lipids on ion channel structure and function. Cell Biochem. Biophys. 2003, 38, 161–190. [Google Scholar] [CrossRef]
- Stalker, T.J.; Newman, D.R.; Ma, P.; Wannemacher, K.M.; Brass, L.F. Platelet signaling. Handb. Exp. Pharm. 2012, 210, 59–85. [Google Scholar]
- Fong, K.P.; Zhu, H.; Span, L.M.; Moore, D.T.; Yoon, K.; Tamura, R.; Yin, H.; De Grado, W.T.; Bennett, J.S. Directly activating the integrin αIIbβ3 initiates outside-in signaling by causing αIIbβ3 clustering. J. Biol. Chem. 2016, 291, 11706–11716. [Google Scholar] [CrossRef] [Green Version]
- Shattil, S.J. Signaling through platelet integrin alpha IIb beta 3: Inside-out, outsidein, and sideways. Thromb. Haemost. 1999, 82, 318–325. [Google Scholar]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arter. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.F.; Van Den Bosch, M.T.; Hunter, R.W.; Sakamoto, K.; Poole, A.W.; Hers, I. Dual regulation of glycogen synthase kinase 3 (gsk3) alpha/beta by protein kinase c (pkc)alpha and AKT promotes thrombin-mediated integrin alphaiibbeta3 activation and granule secretion in platelets. J. Biol. Chem. 2013, 288, 3918–3928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, B.I.; Rocco, M.V.; Bates, J.T.; Chonchol, M.; Hawfield, A.T.; Lash, J.P.; Papademetriou, V.; Sedor, J.R.; Servilla, K.; Kimmel, P.L.; et al. APOL1 renal-risk variants do not associate with incident cardiovascular disease or mortality in the Systolic Blood Pressure Intervention Trial. Kidney Int. Rep. 2017, 2, 713–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendelsohn, M.E.; Loscalzo, J. Role of platelets in cholesteryl ester formation by U-937 cells. J. Clin. Investig. 1988, 81, 62–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, C.J.; Larson, M.G.; Feng, D.; Sutherland, P.A.; Lindpaintner, K.; Myers, R.H.; D’Agostino, R.A.; Levy, D.; Tofler, G.H. Genetic and environmental contributions to platelet aggregation: The Framingham heart study. Circulation 2001, 103, 3051–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lhermusier, T.; Chap, H.; Payrastre, B. Platelet membrane phospholipid asymmetry: From the characterization of a scramblase activity to the identification of an essential protein mutated in Scott syndrome. J. Thromb. Haemost. 2011, 9, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Shattil, S.J.; Anaya-Galindo, R.; Bennett, J.; Colman, R.W.; Cooper, R.A. Platelet hypersensitivity induced by cholesterol incorporation. J. Clin. Investig. 1975, 55, 636–643. [Google Scholar] [CrossRef] [Green Version]
- Shattil, S.J.; Bennett, J.S.; Colman, R.W.; Cooper, R.A. Abnormalities of cholesterol-phospholipid composition in platelets and low-density lipoproteins of human hyperbetalipoproteinemia. J. Lab. Clin. Med. 1977, 89, 341–353. [Google Scholar]
- Lacoste, L.; Lam, J.Y.; Hung, J.; Letchacovski, G.; Solymoss, C.B.; Waters, D. Hyperlipidemia and coronary disease. Correction of the increased thrombogenic potential with cholesterol reduction. Circulation 1995, 92, 3172–3177. [Google Scholar] [CrossRef]
- Panes, O.; González, C.; Hidalgo, P.; Valderas, J.P.; Acevedo, M.; Contreras, S.; Sánchez, X.; Pereira, J.; Rigotti, A.; Mezzano, D. Platelet tissue factor activity and membrane cholesterol are increased in hypercholesterolemia and normalized by rosuvastatin, but not by atorvastatin. Atherosclerosis 2017, 257, 164–171. [Google Scholar] [CrossRef]
- Van Lier, M.; Verhoef, S.; Cauwenberghs, S.; Heemskerk, J.W.; Akkerman, J.W.; Heijnen, H.F. Role of membrane cholesterol in platelet calcium signalling in response to VWF and collagen under stasis and flow. Thromb. Haemost. 2008, 99, 1068–1078. [Google Scholar] [CrossRef]
- Argacha, J.F.; Bourdrel, T.; van de Borne, P. Ecology of the cardiovascular system: A focus on air-related environmental factors. Trends Cardiovasc. Med. 2018, 28, 112–126. [Google Scholar] [CrossRef]
- Melzer, D.; Rice, N.; Depledge, M.H.; Henley, W.E.; Galloway, T.S. Association between serum perfluorooctanoic acid (PFOA) and thyroid disease in the U.S. National Health and Nutrition Examination Survey. Environ. Health Perspect. 2010, 118, 686–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, L.L.; Lin, L.Y.; Su, T.C.; Chen, P.C.; Lin, C.Y. Association between serum perfluorinated chemicals and thyroid function in U.S. adults: The National Health and Nutrition Examination Survey 2007-2010. J. Clin. Endocrinol. Metab. 2013, 98, E1456–E1464. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.C.; Johns, L.E.; Meeker, J.D. Serum biomarkers of exposure to perfluoroalkyl substances in relation to serum testosterone and measures of thyroid function among adults and adolescents from NHANES 2011-2012. Int. J. Environ. Res. Public Health 2015, 12, 6098–6114. [Google Scholar] [CrossRef] [PubMed]
- Webster, G.M.; Rauch, S.A.; Marie, N.S.; Mattman, A.; Lanphear, B.P.; Venners, S.A. Cross-sectional associations of serum perfluoroalkyl acids and thyroid hormones in U.S. adults: Variation according to TPOAb and iodine status (NHANES 2007-2008). Environ. Health Perspect. 2016, 124, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Krassas, G.E.; Poppe, K.; Glinoer, D. Thyroid function and human reproductive health. Endocr. Rev. 2010, 31, 702–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrantonio, M.; Bai, E.; Uccelli, R.; Cordiano, V.; Screpanti, A.; Crosignani, P. Drinking water contamination from perfluoroalkyl substances (PFAS): An ecological mortality study in the Veneto Region, Italy. Eur. J. Public Health 2018, 28, 180–185. [Google Scholar] [CrossRef]
- Frisbee, S.J.; Shankar, A.; Knox, S.S.; Steenland, K.; Savitz, D.A.; Fletcher, T.; Ducatman, A.M. Perfluorooctanoic acid, perfluorooctanesulfonate, and serum lipids in children and adolescents: Results from the C8 Health Project. Arch. Pediatr. Adolesc. Med. 2010, 164, 860–869. [Google Scholar] [CrossRef]
- Steenland, K.; Tinker, S.; Frisbee, S.; Ducatman, A.; Vaccarino, V. Association of perfluorooctanoic acid and perfluorooctane sulfonate with serum lipids among adults living near a chemical plant. Am. J. Epidemiol. 2009, 170, 1268–1278. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Jones, P.D.; De Coen, W.; King, L.; Fraker, P.; Newsted, J.; Giesy, J.P. Alterations in cell membrane properties caused by perfluorinated compounds. Comp. Biochem. Physiol. C Toxicol. Pharm. 2003, 135, 77–88. [Google Scholar] [CrossRef]
- Harada, K.; Xu, F.; Ono, K.; Iijima, T.; Koizumi, A. Effects of PFOS and PFOA on L-type Ca2+ currents in guinea-pig ventricular myocytes. Biochem. Biophys. Res. Commun. 2005, 329, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.; Butenhoff, J.L.; Rogers, J.M. The developmental toxicity of perfluoroalkyl acids and their derivatives. Toxicol. Appl. Pharm. 2004, 198, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matyszewska, D.; Tappura, K.; Orädd, G.; Bilewicz, R. Influence of perfluorinated compounds on the properties of model lipid membranes. J. Phys. Chem. B 2007, 111, 9908–9918. [Google Scholar] [CrossRef] [PubMed]
- Matyszewska, D.; Leitch, J.; Bilewicz, R.; Lipkowski, J. Polarization modulation infrared reflection-absorption spectroscopy studies of the influence of perfluorinated compounds on the properties of a model biological membrane. Langmuir 2008, 24, 7408–7412. [Google Scholar] [CrossRef] [PubMed]
- Merten, M.; Thiagarajan, P. P-selectin expression on platelets determines size and stability of platelet aggregates. Circulation 2000, 102, 1931–1936. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Maeno, T.; Fukumoto, S.; Smith, D.F.; Hyman, M.C.; Jung, S.; Littman, D.R.; Weber, C.; Ley, K. Platelet P-selectin expression is associated with atherosclerotic wall thickness in carotid artery in humans. Circulation 2003, 108, 524–529. [Google Scholar] [CrossRef]
- Campello, E.; Spiezia, L.; Zabeo, E.; Maggiolo, S.; Vettor, R.; Simioni, P. Hypercoagulability detected by whole blood thromboelastometry (ROTEM®) and impedance aggregometry (MULTIPLATE®) in obese patients. Thromb. Res. 2015, 135, 548–553. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Knight, C.J.; Hub, J.S. MemGen: A general web server for the setup of lipid membrane simulation systems. Bioinformatics 2015, 31, 2897–2899. [Google Scholar] [CrossRef]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving physical realism, stereochemistry, and side-chain accuracy in homology modeling: Four approaches that performed well in CASP8. Proteins 2009, 77, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Assinger, A.; Volf, I.; Schmid, D. A novel, rapid method to quantify intraplatelet calcium dynamics by ratiometric flow cytometry. PLoS ONE 2015, 10, e0122527. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Clinical Characteristics | Exposed (48) | Controls (30) | p-Value |
|---|---|---|---|---|
| Demographic/blood parameters | ||||
| Age (years) | // | 18.7 ± 0.6 | 22.1 ± 1.3 | 0.102 |
| BMI (kg/m2) | // | 23.4 ± 3.3 | 24.2 ± 2.6 | 0.432 |
| Platelets (109/L) | // | 227.8 ± 39.9 | 230.1 ± 40.4 | 0.811 |
| Leukocytes (109/L) | // | 6.9 ± 1.6 | 7.2 ± 2.0 | 0.729 |
| Hemoglobin (g/L) | // | 154.1 ± 7.6 | 148.7 ± 6.3 | 0.612 |
| MCV (fL) | // | 86.0 ± 5.7 | 83.8 ± 7.1 | 0.197 |
| PFOA serum levels (ng/mL) | // | 128.0 ± 48.5 | 4.7 ± 2.1 | <0.001 |
| PFOA platelets level (ng/mL) | // | 37.2 ± 15.8 | <LOD | <0.001 |
| Family history | ||||
| Hypertension, diabetes, | Yes | 21 (43.8%) | 11 (36.7%) | 0.638 |
| dyslipidemia | No | 27 (56.2%) | 19 (63.3%) | |
| Early familiar CVD | Yes | 2 (4.2%) | 1 (3.3%) | 1.000 |
| No | 46 (958%) | 29 (96.7%) | ||
| Cigarette smoking | Yes | 10 (208%) | 5 (16.7%) | 0.772 |
| No | 38 (79.2%) | 25 (83.3%) | ||
| Alcohol Consumption | Yes | 2 (4.2%) | 1 (3.3%) | 1.000 |
| No | 46 (95.8%) | 29 (96.7%) | ||
| Aggregometry | ||||
| ADP test [range] (r.r.= 38–85 AUC) | // | 52 [44–62] | 48 [38.5–62.5] | 0.42 |
| ASPI test [range] (r.r.= 39–79 AUC) | // | 67 [57.5–77.0] | 55.5 [50.75–58.8] | 0.002 |
| TRAP-6 test [range] (r.r 69–117 AUC) | // | 100 [91.0–115.5] | 87 [82.8–96.5] | 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Toni, L.; Radu, C.M.; Sabovic, I.; Di Nisio, A.; Dall’Acqua, S.; Guidolin, D.; Spampinato, S.; Campello, E.; Simioni, P.; Foresta, C. Increased Cardiovascular Risk Associated with Chemical Sensitivity to Perfluoro–Octanoic Acid: Role of Impaired Platelet Aggregation. Int. J. Mol. Sci. 2020, 21, 399. https://doi.org/10.3390/ijms21020399
De Toni L, Radu CM, Sabovic I, Di Nisio A, Dall’Acqua S, Guidolin D, Spampinato S, Campello E, Simioni P, Foresta C. Increased Cardiovascular Risk Associated with Chemical Sensitivity to Perfluoro–Octanoic Acid: Role of Impaired Platelet Aggregation. International Journal of Molecular Sciences. 2020; 21(2):399. https://doi.org/10.3390/ijms21020399
Chicago/Turabian StyleDe Toni, Luca, Claudia Maria Radu, Iva Sabovic, Andrea Di Nisio, Stefano Dall’Acqua, Diego Guidolin, Salvatore Spampinato, Elena Campello, Paolo Simioni, and Carlo Foresta. 2020. "Increased Cardiovascular Risk Associated with Chemical Sensitivity to Perfluoro–Octanoic Acid: Role of Impaired Platelet Aggregation" International Journal of Molecular Sciences 21, no. 2: 399. https://doi.org/10.3390/ijms21020399