UBIAD1 Plays an Essential Role in the Survival of Pancreatic Acinar Cells

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Tamoxifen-Induced Ubiad1-Deficient Mice Are Lethal
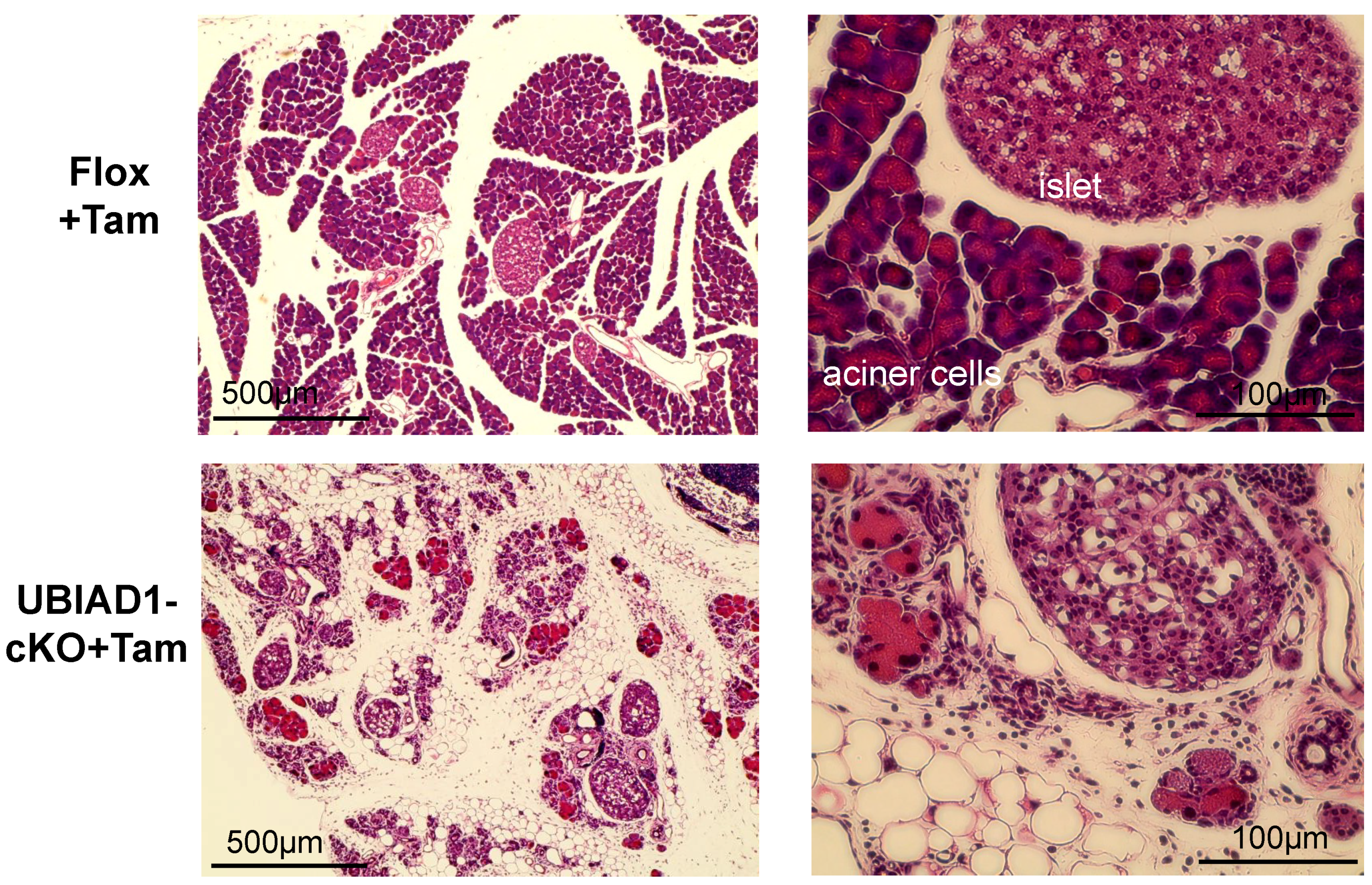
2.2. The Pancreas of Tamoxifen-Induced UBIAD1-Deficient Mice Was Markedly Atrophied
2.3. Disappearance of Pancreatic Acinar Cells and Formation of Vacuoles in the Pancreas of Tamoxifen-Induced UBIAD1-Deficient Mice
2.4. The Vacuoles Found in the Pancreas of Tamoxifen-Treated UBIAD1-cKO Mice Were Adipocytes
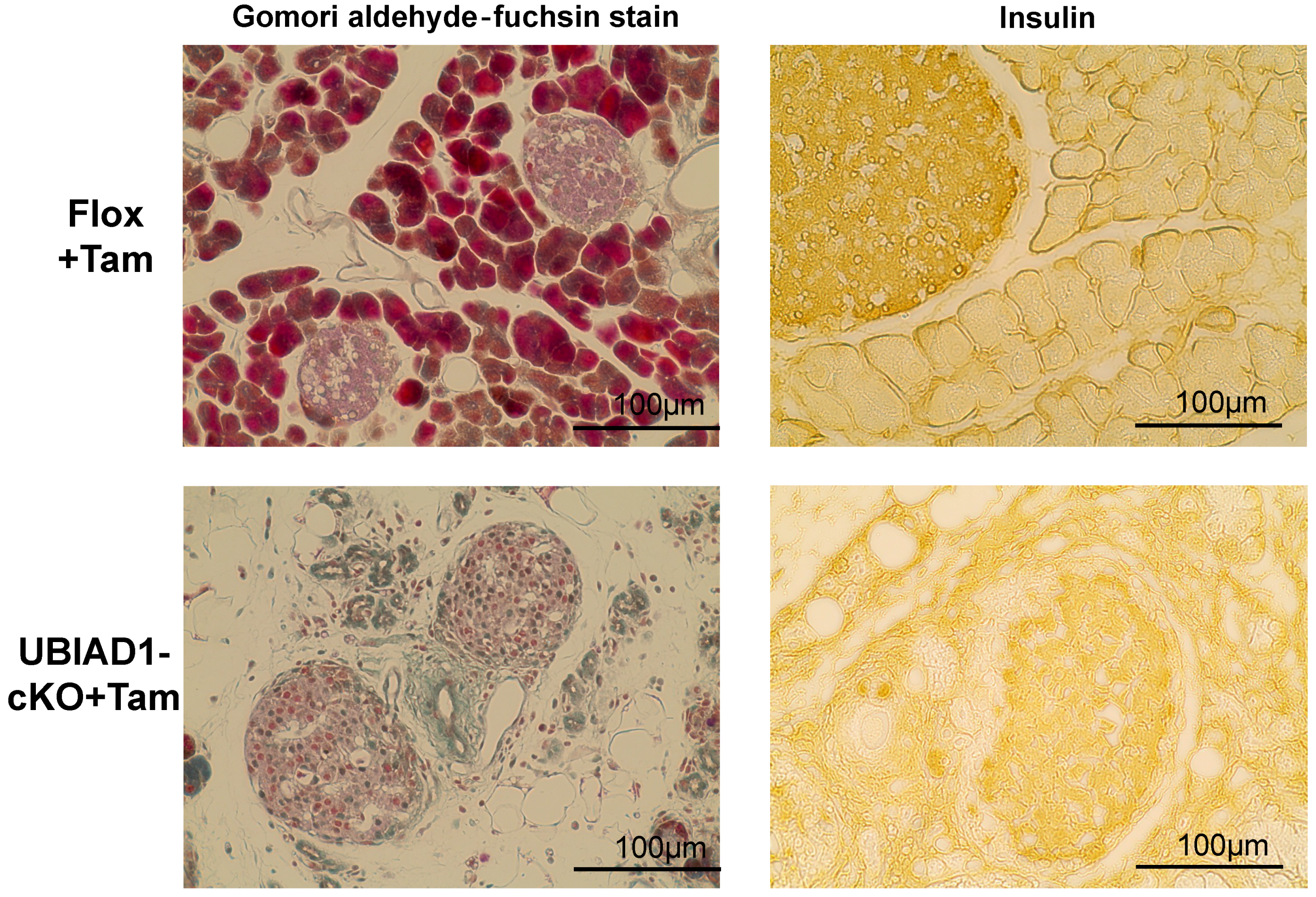
2.5. Pancreatic Acinar Cells in the Pancreas of Tamoxifen-Treated UBIAD1-cKO Mice Increased Oxidative Stress and Autophagy and Disappeared by Apoptosis
2.6. Abundance of Neutrophils and Mesenchymal Stem Cells in the Pancreas of Tamoxifen-Treated UBIAD1-cKO Mice
2.7. Serum Glucose and Insulin Concentrations and Glucose Tolerance in Tamoxifen-Induced UBIAD1-Deficient Mice
2.8. Abnormal Tissue Structure and Decreased Sodium-Dependent Glucose Transporter 1 (SGLT1) and Glucose Transporter 2 (GLUT2) Expressions in the Duodenum of Tamoxifen-Treated UBIAD1-cKO Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Ethics Statement
4.3. Mice
4.4. Genotyping
4.5. Real-Time PCR
4.6. Measurements of MK-4 and MK-4-d7 in Tissues of Mice Administered MD-d8
4.7. Histology and Immunohistochemistry
4.8. Blood Glucose and Insulin
4.9. OGTT
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 4-HNE | 4-hydroxynonenal |
| ES | embryonic stem |
| FRT | flippase |
| FRT | flippase recombinant target |
| GLUT2 | glucose transporter 2 |
| MK-4 | menaquinone-4 |
| MPO | myeloperoxidase |
| OGTT | oral glucose tolerance test |
| PKA | protein kinase A |
| SEM | standard error of mean |
| SGLT1 | sodium-dependent glucose transporter 1 |
| UBIAD1 | UbiA prenyltransferase domain-containing protein 1 |
References
- Furie, B.; Bouchard, B.A.; Furie, B.C. Vitamin K-dependent biosynthesis of gamma-carboxyglutamic acid. Blood 1999, 93, 1798–1808. [Google Scholar]
- Stanley, T.B.; Wu, S.M.; Houben, R.J.; Mutucumarana, V.P.; Stafford, D.W. Role of the propeptide and gamma-glutamic acid domain of factor IX for in vitro carboxylation by the vitamin K-dependent carboxylase. Biochemistry 1998, 37, 13262–13268. [Google Scholar] [CrossRef]
- Azuma, K.; Urano, T.; Ouchi, Y.; Inoue, S. Vitamin K2 suppresses proliferation and motility of hepatocellular carcinoma cells by activating steroid and xenobiotic receptor. Endocrine. J. 2009, 56, 843–849. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grün, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Vitamin K2 induces phosphorylation of protein kinase A and expression of novel target genes in osteoblastic cells. J. Mol. Endocrinol. 2007, 39, 239–247. [Google Scholar] [CrossRef]
- Vos, M.; Esposito, G.; Edirisinghe, J.N.; Vilain, S.; Haddad, D.M.; Slabbaert, J.R.; Van Meensel, S.; Schaap, O.; De Strooper, B.; Meganathan, R.; et al. Vitamin K2 is a mitochondrial electron carrier that rescues pink1 deficiency. Science 2012, 336, 1306–1310. [Google Scholar] [CrossRef]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (vitamin K3) is a catabolic product of oral phylloquinone (vitamin K1) in the intestine and a circulating precursor of tissue menaquinone-4 (vitamin K2) in rats. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef]
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of phylloquinone (Vitamin K1) into menaquinone-4 (Vitamin K2) in mice: two possible routes for menaquinone-4 accumulation in cerebra of mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar] [CrossRef]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Shearer, M.J.; Okano, T. Key pathways and regulators of vitamin K function and intermediary metabolism. Annu. Rev. Nutr. 2018, 38, 127–151. [Google Scholar] [CrossRef]
- Nickerson, M.L.; Bosley, A.D.; Weiss, J.S.; Kostiha, B.N.; Hirota, Y.; Brandt, W.; Esposito, D.; Kinoshita, S.; Wessjohann, L.; Morham, S.G.; et al. The UBIAD1 prenyltransferase links menaquinone-4 synthesis to cholesterol metabolic enzymes. Hum. Mutat. 2013, 34, 317–329. [Google Scholar] [CrossRef]
- Hirota, Y.; Nakagawa, K.; Sawada, N.; Okuda, N.; Suhara, Y.; Uchino, Y.; Kimoto, T.; Funahashi, N.; Kamao, M.; Tsugawa, N.; et al. Functional characterization of the vitamin K2 biosynthetic enzyme UBIAD1. PLoS ONE 2015, 10, e0125737. [Google Scholar] [CrossRef]
- Nakagawa, K.; Sawada, N.; Hirota, Y.; Uchino, Y.; Suhara, Y.; Hasegawa, T.; Amizuka, N.; Okamoto, T.; Tsugawa, N.; Kamao, M.; et al. Vitamin K2 biosynthetic enzyme, UBIAD1 is essential for embryonic development of mice. PLoS ONE 2014, 9, e104078. [Google Scholar] [CrossRef]
- Thomas, D.D.; Krzykowski, K.J.; Engelke, J.A.; Groblewski, G.E. Exocrine pancreatic secretion of phospholipid, menaquinone-4, and caveolin-1 in vivo. Biochem. Biophys. Res. Commun. 2004, 319, 974–979. [Google Scholar] [CrossRef]
- Davidson, R.T.; Foley, A.L.; Engelke, J.A.; Suttie, J.W. Conversion of dietary phylloquinone to tissue menadione-4 in rats is not dependent on gut bacteria. J. Nutr. 1998, 128, 220–223. [Google Scholar] [CrossRef]
- Thijssen, H.H.W.; Drittij-Reijnders, M.J.; Fischer, M.A.J.G. Phylloquinone and menaquinone-4 distribution in rats: synthesis rather than uptake determines meaquinone-4 organ concentrations. J. Nutr. 1995, 126, 537–543. [Google Scholar] [CrossRef]
- Furuhashi, M.; Hotamisligil, G.S. Fatty acid-binding proteins: role in metabolic diseases and potential as drug targets. Nat. Rev. Drug Discov 2008, 7, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Corrales, P.; Vidal-Puig, A.; Medina-Gómez, G. PPARs and metabolic disorders associated with challenged adipose tissue plasticity. Int. J. Mol. Sci. 2018, 19, 2124. [Google Scholar] [CrossRef]
- Kimmel, A.R.; Sztalryd, C. The perilipins: major cytosolic lipid droplet–associated proteins and their roles in cellular lipid storage, mobilization, and systemic homeostasis. Annu. Rev. Nutr. 2016, 36, 471–509. [Google Scholar] [CrossRef]
- Itabe, H.; Yamaguchi, T.; Nimura, S.; Sasabe, N. Perilipins: a diversity of intracellular lipid droplet proteins. Lipids Health Dis. 2017, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Coutu, D.L.; Wu, J.H.; Monette, A.; Rivard, G.E.; Blostein, M.D.; Galipeau, J. Periostin, a member of a novel family of vitamin K-dependent proteins, is expressed by mesenchymal stromal cells. J. Biol. Chem. 2008, 283, 17991–18001. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, S.; Regel, I.; Steiger, K.; Wagner, N.; Thorwirth, M.; Schlitter, A.M.; Esposito, I.; Michalski, C.W.; Friess, H.; Kleeff, J.; et al. Loss of periostin results in impaired regeneration and pancreatic atrophy after cerulein-induced pancreatitis. Am. J. Pathol. 2016, 186, 24–31. [Google Scholar] [CrossRef]
- Lankisch, P.G.; Apte, M.; Banks, P.A. Acute pancreatitis. Lancet 2015, 386, 85–96. [Google Scholar] [CrossRef]
- Leung, P.S.; Ip, S.P. Pancreatic acinar cell: its role in acute pancreatitis. Int. J. Biochem. Cell Biol. 2006, 38, 1024–1030. [Google Scholar] [CrossRef] [PubMed]
- Robles, L.; Vaziri, N.D.; Ichii, H. Role of oxidative stress in the pathogenesis of pancreatitis: effect of antioxidant therapy. Pancreat. Disord. Ther. 2014, 3, 112–119. [Google Scholar] [CrossRef]
- Logsdon, C.D.; Ji, B. The role of protein synthesis and digestive enzymes in acinar cell injury. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 362–370. [Google Scholar] [CrossRef] [Green Version]
- Sah, R.P.; Dawra, R.K.; Saluja, A.K. New insights into the pathogenesis of pancreatitis. Curr. Opin. Gastroenterol. 2013, 29, 523–530. [Google Scholar] [CrossRef]
- Meher, S.; Rath, S.; Sharma, R.; Rout, B.; Mishra, T.S.; Sasmal, P.K.; Sinha, M.K. Pathophysiology of oxidative stress and antioxidant therapy in acute pancreatitis. J. Mol. Biomark. Diagn. 2015, 6, 251. [Google Scholar] [CrossRef]
- Hashimoto, D.; Ohmuraya, M.; Hirota, M.; Yamamoto, A.; Suyama, K.; Ida, S.; Okumura, Y.; Takahashi, E.; Kido, H.; Araki, K.; et al. Involvement of autophagy in trypsinogen activation within the pancreatic acinar cells. J. Cell Biol. 2008, 181, 1065–1072. [Google Scholar] [CrossRef] [Green Version]
- Breitzig, M.; Bhimineni, C.; Lockey, R.; Kolliputi, N. 4-Hydroxy-2-nonenal: a critical target in oxidative stress? Am. J. Physiol. Cell Physiol. 2016, 311, C537–C543. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lin, J.C.; Wang, H.; Peterson, J.W.; Furie, B.C.; Furie, B.; Booth, S.L.; Volpe, J.J.; Rosenberg, P.A. Novel role of vitamin k in preventing oxidative injury to developing oligodendrocytes and neurons. J. Neurosci. 2003, 23, 5816–5826. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, H.; Rosenberg, P.A. Vitamin K prevents oxidative cell death by inhibiting activation of 12-lipoxygenase in developing oligodendrocytes. J. Neurosci. Res. 2009, 87, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Mugoni, V.; Postel, R.; Catanzaro, V.; De Luca, E.; Turco, E.; Digilio, G.; Silengo, L.; Murphy, M.P.; Medana, C.; Stainier, D.Y.; et al. Ubiad1 is an antioxidant enzyme that regulates eNOS activity by CoQ10 synthesis. Cell 2013, 152, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Pfützner, A.; Schipper, D.; Pansky, A.; Kleinfeld, C.; Roitzheim, B.; Tobiasch, E. Mesenchymal stem cell differentiation into adipocytes is equally induced by insulin and proinsulin in vitro. Int. J. Stem Cells. 2017, 10, 154–159. [Google Scholar] [CrossRef]
- Wright, E.M.; Loo, D.D.; Hirayama, B.A. Biology of human sodium glucose transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Inoue, R.; Matsumoto, M.; Yajima, T.; Ushida, K.; Iwanaga, T. Comparative expression of hexose transporters (SGLT1, GLUT1, GLUT2 and GLUT5) throughout the mouse gastrointestinal tract. Histochem. Cell Biol. 2011, 135, 183–194. [Google Scholar] [CrossRef]
- Gorboulev, V.; Schurmann, A.; Vallon, V.; Kipp, H.; Jaschke, A.; Klessen, D.; Friedrich, A.; Scherneck, S.; Rieg, T.; Cunard, R.; et al. Na(+)-d-glucose cotransporter SGLT1 is pivotal for intestinal glucose absorption and glucose-dependent incretin secretion. Diabetes 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Röder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef]
- Manna, P.; Kalita, J. Beneficial role of vitamin K supplementation on insulin sensitivity, glucose metabolism, and the reduced risk of type 2 diabetes: A review. Nutrition 2016, 32, 732–739. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flox Mice | Flox Mice + Tam | UBIAD1-cKO | UBIAD1-Cko + Tam | |
|---|---|---|---|---|
| Body weight (g) | 23.25 ± 0.39 | 23.45 ± 0.65 | 22.54 ± 0.39 | 22.03 ± 0.46 |
| Cerebrum (g) | 0.305 ± 0.023 | 0.287 ± 0.003 | 0.311 ± 0.004 | 0.294 ± 0.003 |
| Cerebellum (g) | 0.056 ± 0.005 | 0.059 ± 0.001 | 0.057 ± 0.004 | 0.055 ± 0.002 |
| Liver (g) | 0.887 ± 0.041 | 0.883 ± 0.014 | 0.872 ± 0.045 | 0.867 ± 0.020 |
| Kidney (g) | 0.257 ± 0.010 | 0.245 ± 0.006 | 0.254 ± 0.007 | 0.246 ± 0.010 |
| Pancreas (g) | 0.196 ± 0.011 | 0.192 ± 0.007 | 0.192 ± 0.005 | 0.054 ± 0.011 *** |
| MK-4 concentration (pmol/g) | MK-4-d7 concentration (pmol/g) | |||
|---|---|---|---|---|
| Flox + Tam | UBIAD1-cKO + Tam | Flox + Tam | UBIAD1-cKO + Tam | |
| Cerebrum | 429.67 ± 16.55 | 135.92 ± 14.67*** | 20.48 ± 1.15 | 2.38 ± 0.49*** |
| Cerebellum | 801.12 ± 76.35 | 223.46 ± 26.38** | 31.82 ± 5.95 | 2.67 ± 1.29*** |
| Kidney | 232.00 ± 12.03 | 24.45 ± 2.34*** | 108.18 ± 14.11 | 7.18 ± 2.46** |
| Pancreas | 1140.91 ± 108.15 | 96.24 ± 24.15** | 61.69 ± 5.21 | N.D.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakagawa, K.; Fujiwara, K.; Nishimura, A.; Murakami, C.; Kawamoto, K.; Ichinose, C.; Kunitou, Y.; Suhara, Y.; Okano, T.; Hasegawa, H. UBIAD1 Plays an Essential Role in the Survival of Pancreatic Acinar Cells. Int. J. Mol. Sci. 2019, 20, 1971. https://doi.org/10.3390/ijms20081971
Nakagawa K, Fujiwara K, Nishimura A, Murakami C, Kawamoto K, Ichinose C, Kunitou Y, Suhara Y, Okano T, Hasegawa H. UBIAD1 Plays an Essential Role in the Survival of Pancreatic Acinar Cells. International Journal of Molecular Sciences. 2019; 20(8):1971. https://doi.org/10.3390/ijms20081971
Chicago/Turabian StyleNakagawa, Kimie, Kiyomi Fujiwara, Akihiro Nishimura, Chinami Murakami, Kanaha Kawamoto, Chihiro Ichinose, Yumi Kunitou, Yoshitomo Suhara, Toshio Okano, and Hiroshi Hasegawa. 2019. "UBIAD1 Plays an Essential Role in the Survival of Pancreatic Acinar Cells" International Journal of Molecular Sciences 20, no. 8: 1971. https://doi.org/10.3390/ijms20081971