The Desaturase Gene Family is Crucially Required for Fatty Acid Metabolism and Survival of the Brown Planthopper, Nilaparvata lugens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
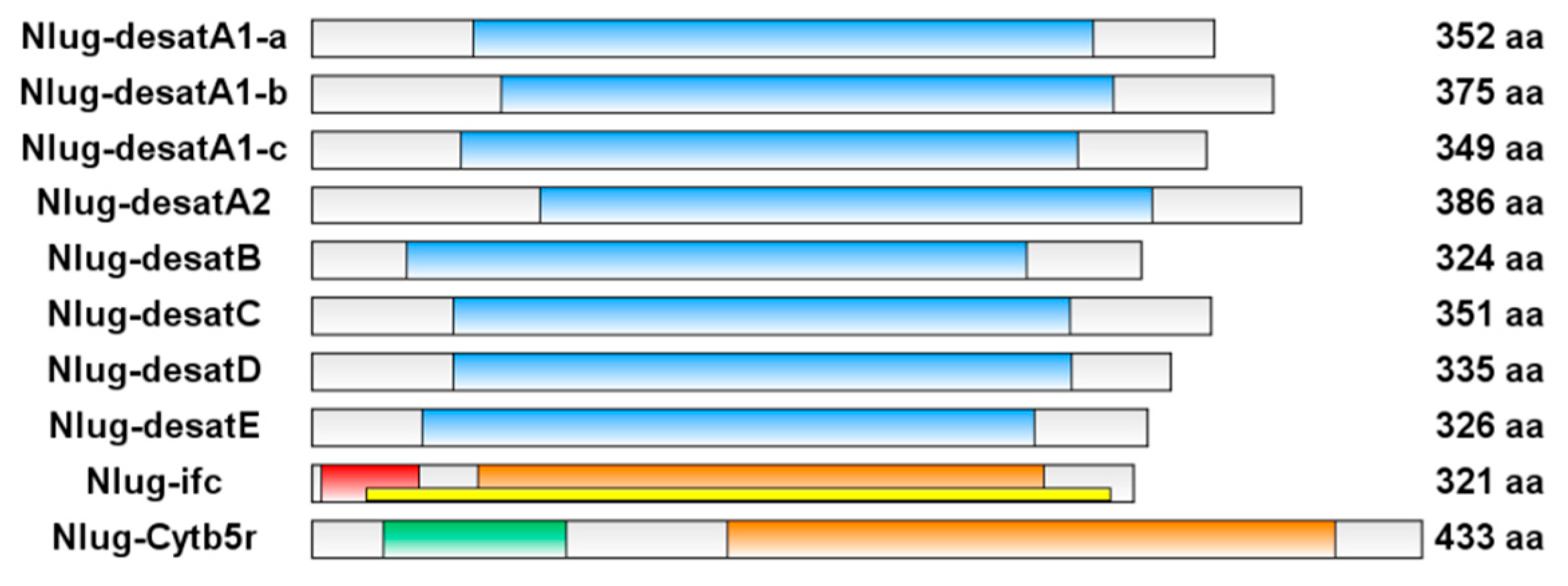
2.1. Brown Planthopper Genome Contains 10 Putative acyl-CoA Desaturases
2.2. Phylogenetic Analyses of Nlug-desat Genes
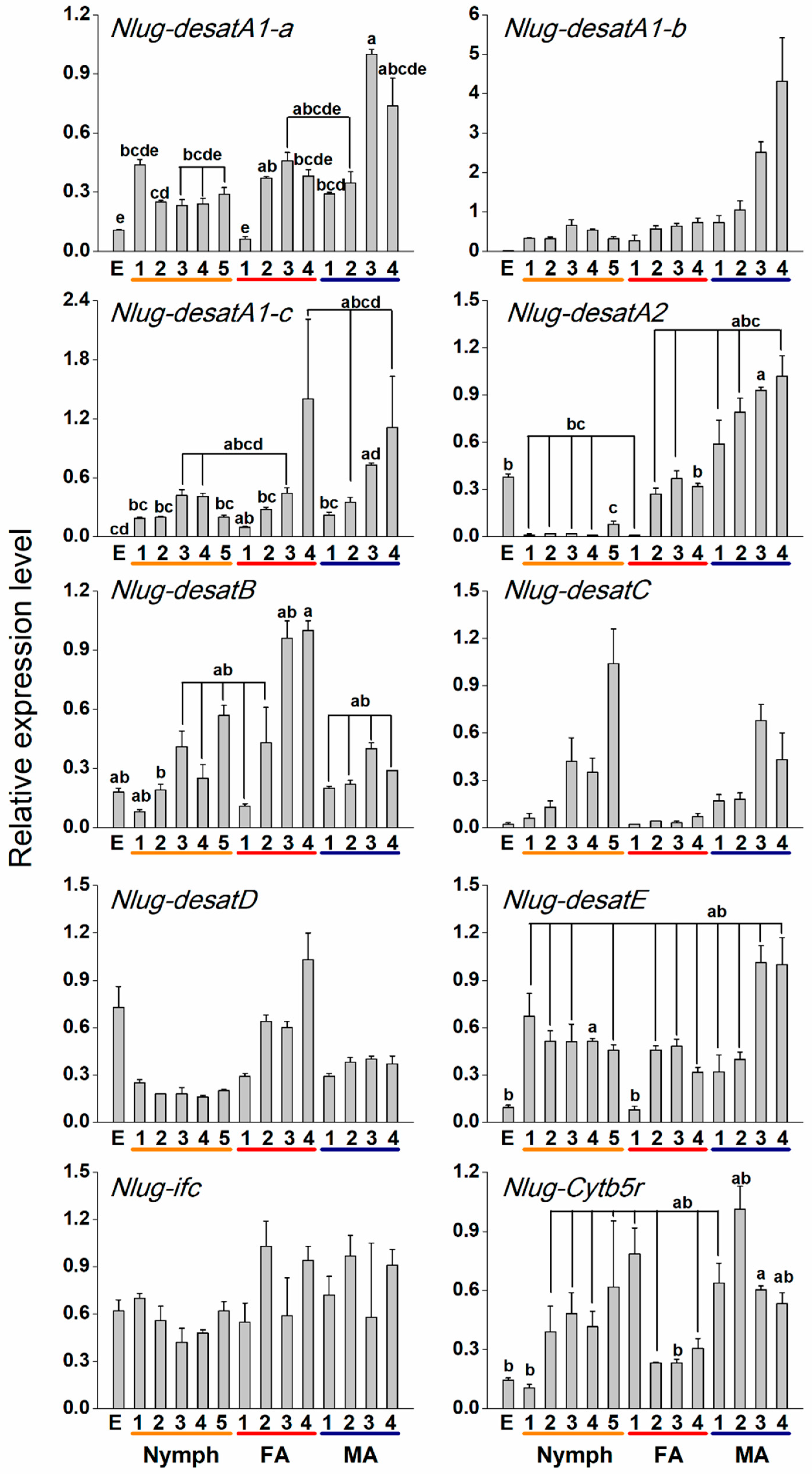
2.3. Developmental Expression Patterns of Nlug-desat Genes
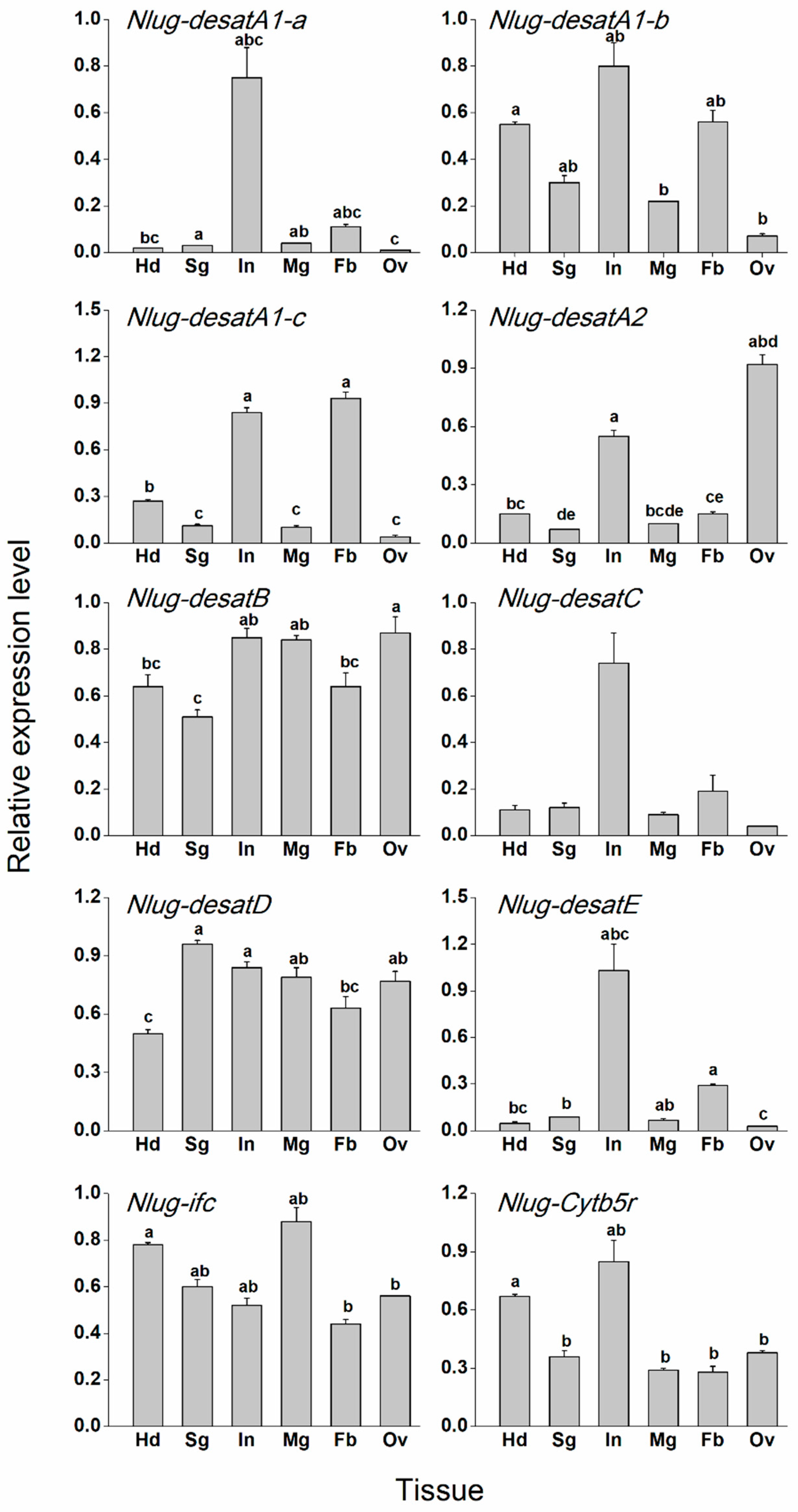
2.4. Tissue-Specific Expression Patterns of Nlug-desat Genes
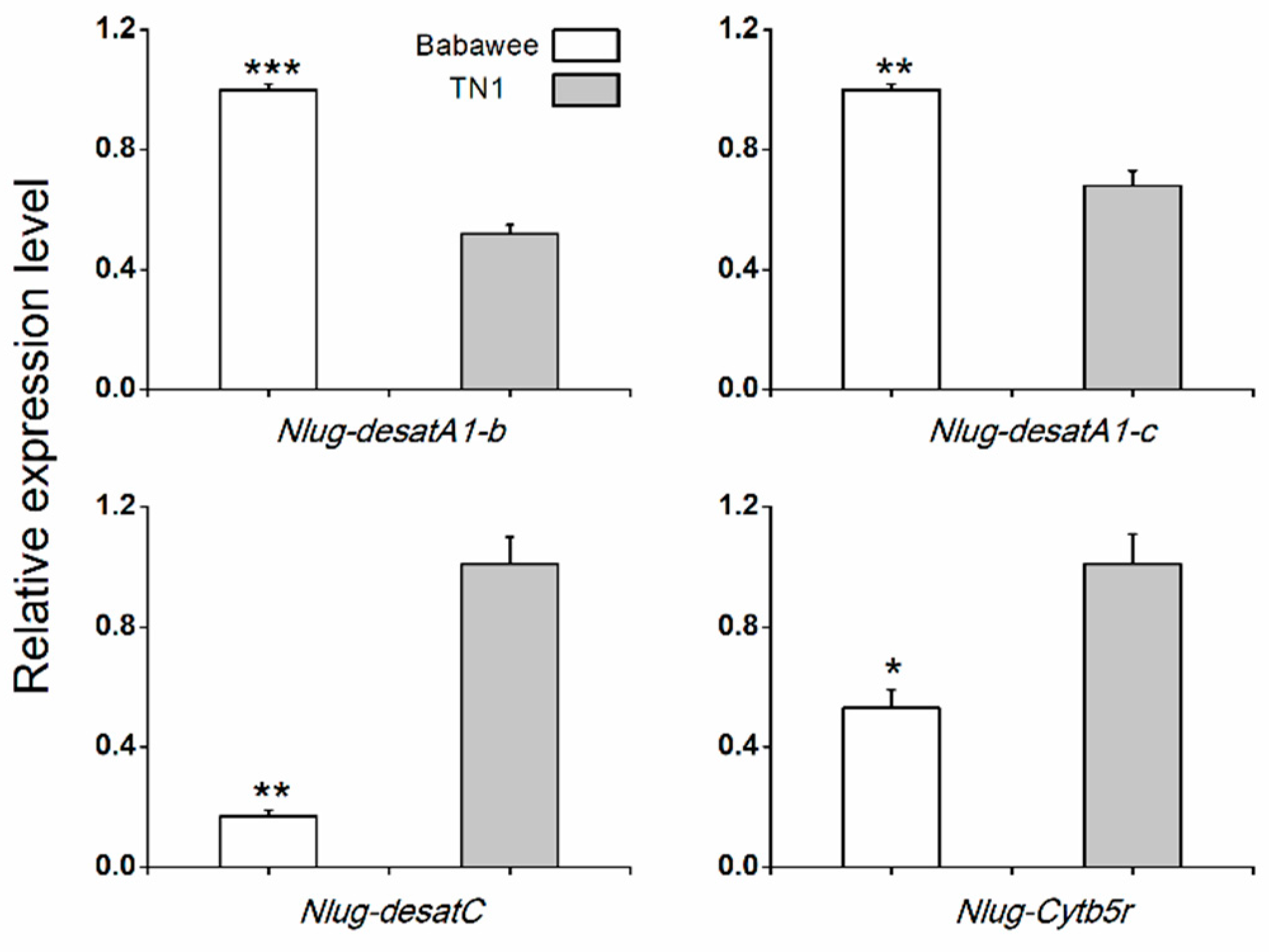
2.5. Differences in Nlug-desat Genes Expression between TN1 and Babawee Populations
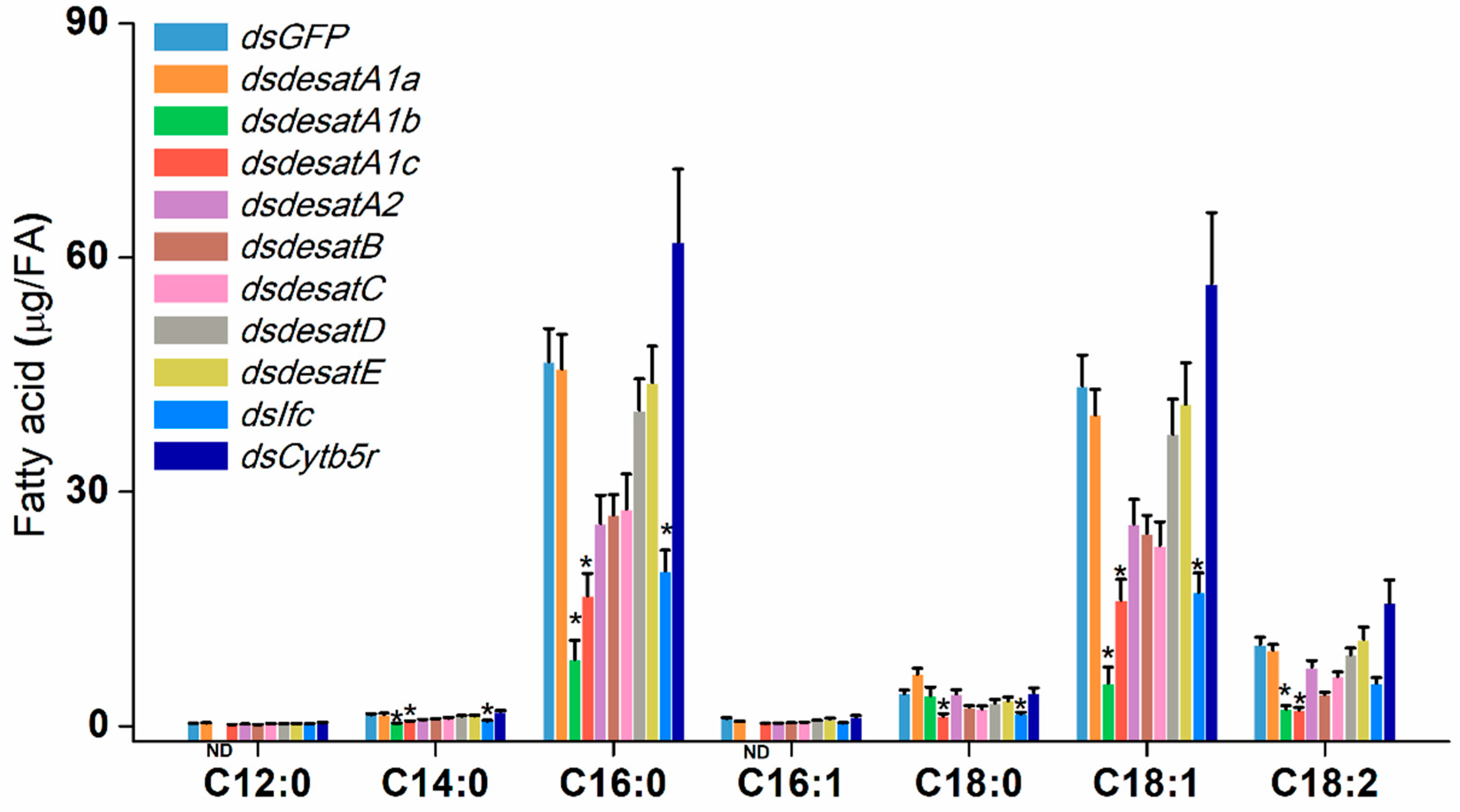
2.6. Influence of Nlug-desats Suppression on Fatty Acids of BPH Female Adults
2.7. Knockdown of Nlug-desats Decrease the Survival Rate of BPH Nymphs
3. Discussion
4. Materials and methods
4.1. Insect Rearing and Plant Growth
4.2. Identification and Amplification of Nlug-desat Genes
4.3. Sequence Analysis and Phylogenetic Construction
4.4. RNA Preparation and Real Time qPCR
4.5. RNAi Experiment
4.6. Fatty Acid Analysis
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hazel, J.R.; Eugene Williams, E. The role of alterations in membrane lipid composition in enabling physiological adaptation of organisms to their physical environment. Prog. Lipid Res. 1990, 29, 167–227. [Google Scholar] [CrossRef]
- Holthuis, J.C.M.; Menon, A.K. Lipid landscapes and pipelines in membrane homeostasis. Nature 2014, 510, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Tiku, P.E.; Gracey, A.Y.; Macartney, A.I.; Beynon, R.J.; Cossins, A.R. Cold-induced expression of delta 9-desaturase in carp by transcriptional and posttranslational mechanisms. Science 1996, 271, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Ntambi, J.M. Role of stearoyl-coenzyme A desaturase in lipid metabolism. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 113–121. [Google Scholar] [CrossRef]
- Vigh, L.; Los, D.A.; Horvath, I.; Murata, N. The primary signal in the biological perception of temperature: Pd-catalyzed hydrogenation of membrane lipids stimulated the expression of the desA gene in Synechocystis PCC6803. Proc. Natl. Acad. Sci. USA 1993, 90, 9090–9094. [Google Scholar] [CrossRef]
- Li, T.; Cofer, T.M.; Engelberth, M.J.; Engelberth, J. Defense priming by non-jasmonate producing fatty acids in maize (Zea mays). Plant Signal. Behav. 2016, 11, e1243635. [Google Scholar] [CrossRef] [PubMed]
- Sperling, P.; Ternes, P.; Zank, T.K.; Heinz, E. The evolution of desaturases. Prostaglandins Leukot. Essent. Fat. Acids 2003, 68, 73–95. [Google Scholar] [CrossRef]
- Kachroo, A.; Shanklin, J.; Whittle, E.; Lapchyk, L.; Hildebrand, D.; Kachroo, P. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis. Plant Mol. Biol. 2007, 63, 257–271. [Google Scholar] [CrossRef]
- Shanklin, J.; Whittle, E.; Fox, B.G. Eight histidine residues are catalytically essential in a membrane-associated iron enzyme, stearoyl-CoA desaturase, and are conserved in alkane Hydroxylase and xylene Monooxygenase. Biochemistry 1994, 33, 12787–12794. [Google Scholar] [CrossRef]
- Los, D.A.; Murata, N. Structure and expression of fatty acid desaturases. Biochim. Biophys. Acta Lipids Lipid Metab. 1998, 1394, 3–15. [Google Scholar] [CrossRef]
- Spychalla, J.P.; Kinney, A.J.; Browse, J. Identification of an animal ω-3 fatty acid desaturase by heterologous expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 1142–1147. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Yoshizawa, A.C.; Okuda, S.; Kuma, K.; Goto, S.; Kanehisa, M. The repertoire of desaturases and elongases reveals fatty acid variations in 56 eukaryotic genomes. J. Lipid Res. 2008, 49, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Helmkampf, M.; Cash, E.; Gadau, J. Evolution of the insect desaturase gene family with an emphasis on social Hymenoptera. Mol. Biol. Evol. 2015, 32, 456–471. [Google Scholar] [CrossRef] [PubMed]
- Ternes, P.; Franke, S.; Zähringer, U.; Sperling, P.; Heinz, E. Identification and characterization of a sphingolipid Δ4-desaturase family. J. Biol. Chem. 2002, 277, 25512–25518. [Google Scholar] [CrossRef] [PubMed]
- Aronstein, K.A.; Murray, K.D.; Saldivar, E. Transcriptional responses in Honey Bee larvae infected with chalkbrood fungus. BMC Genomics 2010, 11, 391. [Google Scholar] [CrossRef]
- Vesala, L.; Salminen, T.S.; Laiho, A.; Hoikkala, A.; Kankare, M. Cold tolerance and cold-induced modulation of gene expression in two Drosophila virilis group species with different distributions. Insect Mol. Biol. 2012, 21, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Dennis, A.B.; Dunning, L.T.; Sinclair, B.J.; Buckley, T.R. Parallel molecular routes to cold adaptation in eight genera of New Zealand stick insects. Sci. Rep. 2015, 5, 13965. [Google Scholar] [CrossRef] [Green Version]
- Phan, V.H.; Herr, D.R.; Panton, D.; Fyrst, H.; Saba, J.D.; Harris, G.L. Disruption of sphingolipid metabolism elicits apoptosis-associated reproductive defects in Drosophila. Dev. Biol. 2007, 309, 329–341. [Google Scholar] [CrossRef]
- Bousquet, F.; Nojima, T.; Houot, B.; Chauvel, I.; Chaudy, S.; Dupas, S.; Yamamoto, D.; Ferveur, J.-F. Expression of a desaturase gene, DESAT1, in neural and nonneural tissues separately affects perception and emission of sex pheromones in Drosophila. Proc. Natl. Acad. Sci. USA 2012, 109, 249–254. [Google Scholar] [CrossRef]
- Kayukawa, T.; Chen, B.; Hoshizaki, S.; Ishikawa, Y. Upregulation of a desaturase is associated with the enhancement of cold hardiness in the onion maggot, Delia antiqua. Insect Biochem. Mol. Biol. 2007, 37, 1160–1167. [Google Scholar] [CrossRef]
- Haritos, V.S.; Horne, I.; Damcevski, K.; Glover, K.; Gibb, N.; Okada, S.; Hamberg, M. The convergent evolution of defensive polyacetylenic fatty acid biosynthesis genes in soldier beetles. Nat. Commun. 2012, 3, 1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; da Cruz, T.C.; Pulfemuller, A.; Grégoire, S.; Ferveur, J.F.; Moussian, B. Inhibition of fatty acid desaturases in Drosophila melanogaster larvae blocks feeding and developmental progression. Arch. Insect Biochem. Physiol. 2016, 92, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Köhler, K.; Brunner, E.; Xue, L.G.; Boucke, K.; Greber, U.F.; Mohanty, S.; Barth, J.M.I.; Wenk, M.R.; Hafen, E. A combined proteomic and genetic analysis identifies a role for the lipid desaturase Desat1 in starvation-induced autophagy in Drosophila. Autophagy 2009, 5, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Ntambi, J.M.; Miyazaki, M.; Stoehr, J.P.; Lan, H.; Kendziorski, C.M.; Yandell, B.S.; Song, Y.; Cohen, P.; Friedman, J.M.; Attie, A.D. Loss of stearoyl-CoA desaturase-1 function protects mice against adiposity. Proc. Natl. Acad. Sci. USA 2002, 99, 11482–11486. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.J.; Browse, J.; Watts, J.L. Fatty acid desaturation and the regulation of adiposity in Caenorhabditis elegans. Genetics 2007, 176, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, M.; Chertemps, T.; Labeur, C.; Wicker-Thomas, C. Mutations in the DESAT1 gene reduces the production of courtship stimulatory pheromones through a marked effect on fatty acids in Drosophila melanogaster. Insect Biochem. Mol. Biol. 2005, 35, 911–920. [Google Scholar] [CrossRef]
- Knipple, D.C.; Rosenfield, C.L.; Nielsen, R.; You, K.M.; Jeong, S.E. Evolution of the integral membrane desaturase gene family in moths and flies. Genetics 2002, 162, 1737–1752. [Google Scholar] [PubMed]
- Moto, K.; Suzuki, M.G.; Hull, J.J.; Kurata, R.; Takahashi, S.; Yamamoto, M.; Okano, K.; Imai, K.; Ando, T.; Matsumoto, S. Involvement of a bifunctional fatty-acyl desaturase in the biosynthesis of the silkmoth, Bombyx mori, sex pheromone. Proc. Natl. Acad. Sci. USA 2004, 101, 8631–8636. [Google Scholar] [CrossRef] [PubMed]
- Dallerac, R.; Labeur, C.; Jallon, J.-M.; Knipple, D.C.; Roelofs, W.L.; Wicker-Thomas, C. A delta 9 desaturase gene with a different substrate specificity is responsible for the cuticular diene hydrocarbon polymorphism in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2000, 97, 9449–9454. [Google Scholar] [CrossRef]
- Labeur, C.; Dallerac, R.; Wicker-Thomas, C. Involvement of DESAT1 gene in the control of Drosophila melanogaster pheromone biosynthesis. Genetica 2002, 114, 269–274. [Google Scholar] [CrossRef]
- Buček, A.; Vogel, H.; Matoušková, P.; Prchalová, D.; Žáček, P.; Vrkoslav, V.; Šebesta, P.; Svatoš, A.; Jahn, U.; Valterová, I. The role of desaturases in the biosynthesis of marking pheromones in bumblebee males. Insect Biochem. Mol. Biol. 2013, 43, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Horne, I.; Gibb, N.; Damcevski, K.; Glover, K.; Haritos, V.S. Two conserved Z9-octadecanoic acid desaturases in the red flour beetle. Tribolium castaneum. Gene 2010, 468, 41–47. [Google Scholar]
- Haritos, V.S.; Horne, I.; Damcevski, K.; Glover, K.; Gibb, N. Unexpected functional diversity in the fatty acid desaturases of the flour beetle Tribolium castaneum and identification of key residues determining activity. Insect Biochem. Mol. Biol. 2014, 51, 62–70. [Google Scholar] [CrossRef]
- Riddervold, M.H.; Tittiger, C.; Blomquist, G.J.; Borgeson, C.E. Biochemical and molecular characterizaton of house cricket (Acheta domesticus, Orthoptera: Gryllidae) Δ9 desaturase. Insect Biochem. Mol. Biol. 2002, 32, 1731–1740. [Google Scholar] [CrossRef]
- Luo, J.; Li, Z.; Ma, C.; Zhang, Z.; Hull, J.J.; Lei, C.; Jin, S.; Chen, L. Knockdown of a metathoracic scent gland desaturase enhances the production of (E)-4-oxo-2-hexenal and suppresses female sexual attractiveness in the plant bug Adelphocoris suturalis. Insect Mol. Biol. 2017, 26, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Brar, D.S.; Virk, P.S.; Jena, K.K.; Khush, G.S. Breeding for resistance to planthoppers in rice. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 401–427. [Google Scholar]
- Ling, K.C.; Tiongco, E.R.; Aguiero, V.M. Rice ragged stunt, a new virus disease. Plant Dis. Rep. 1978, 62, 701–705. [Google Scholar]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 1996, 34, 249–274. [Google Scholar] [CrossRef]
- Sōgawa, K. The rice brown planthopper: Feeding physiology and host plant interactions. Annu. Rev. Entomol. 1982, 27, 49–73. [Google Scholar] [CrossRef]
- Watanabe, T.; Kitagawa, H. Photosynthesis and translocation of assimilates in rice plants following phloem feeding by the planthopper Nilaparvata lugens (Homoptera: Delphacidae). J. Econ. Entomol. 2000, 93, 1192–1198. [Google Scholar] [CrossRef]
- Yu, H.; Ji, R.; Ye, W.; Chen, H.; Lai, W.; Fu, Q.; Lou, Y. Transcriptome analysis of fat bodies from two brown planthopper (Nilaparvata lugens) populations with different virulence levels in rice. PLoS ONE 2014, 9, e88528. [Google Scholar] [CrossRef]
- Jagadeesan, R.; Collins, P.J.; Daglish, G.J.; Ebert, P.R.; Schlipalius, D.I. Phosphine resistance in the rust red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae): Inheritance, gene interactions and fitness costs. PLoS ONE 2012, 7, e31582. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Schlipalius, D.I.; Collins, P.J.; Swain, A.J.; Ebert, P.R. Inheritance and relative dominance, expressed as toxicity response and delayed development, of phosphine resistance in immature stages of Rhyzopertha dominica (F.) (Coleoptera: Bostrichidae). J. Stored Prod. Res. 2012, 51, 74–80. [Google Scholar] [CrossRef]
- Daglish, G.J.; Nayak, M.K.; Pavic, H. Phosphine resistance in Sitophilus oryzae (L.) from eastern Australia: Inheritance, fitness and prevalence. J. Stored Prod. Res. 2014, 59, 237–244. [Google Scholar] [CrossRef]
- Schlipalius, D.I.; Tuck, A.G.; Jagadeesan, R.; Nguyen, T.; Kaur, R.; Subramanian, S.; Barrero, R.; Nayak, M.; Ebert, P.R. Variant linkage analysis using de novo transcriptome sequencing identifies a conserved phosphine resistance gene in insects. Genetics 2018, 209, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Basu, J.; Li, Z. The Des-1 protein, required for central spindle assembly and cytokinesis, is associated with mitochondria along the meiotic spindle apparatus and with the contractile ring during male meiosis in Drosophila melanogaster. Mol. Gen. Genet. 1998, 259, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.W.; De Bie, T.; Stajich, J.E.; Nguyen, C.; Cristianini, N. Estimating the tempo and mode of gene family evolution from comparative genomic data. Genome Res. 2005, 15, 1153–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodson, L.; Fielding, B.A. Stearoyl-CoA desaturase: Rogue or innocent bystander? Prog. Lipid Res. 2013, 52, 15–42. [Google Scholar] [CrossRef]
- Bousquet, F.; Chauvel, I.; Flaven-Pouchon, J.; Farine, J.-P.; Ferveur, J.-F. Dietary rescue of altered metabolism gene reveals unexpected Drosophila mating cues. J. Lipid Res. 2016, 57, 443–450. [Google Scholar] [CrossRef]
- Roy, A.; Walker, W.B.; Vogel, H.; Chattington, S.; Larsson, M.C.; Anderson, P.; Heckel, D.G.; Schlyter, F. Diet dependent metabolic responses in three generalist insect herbivores Spodoptera spp. Insect Biochem. Mol. Biol. 2016, 71, 91–105. [Google Scholar] [CrossRef]
- Keays, M.C.; Barker, D.; Wicker-Thomas, C.; Ritchie, M.G. Signatures of selection and sex-specific expression variation of a novel duplicate during the evolution of the Drosophila desaturase gene family. Mol. Ecol. 2011, 20, 3617–3630. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevenet, F.; Brun, C.; Bañuls, A.L.; Jacq, B.; Christen, R. TreeDyn: Towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics 2006, 7, 439. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Lu, Y.; Zhu, X.; Wan, H.; Shakeel, M.; Zhan, S.; Jin, B.-R.; Li, J. Selection and evaluation of potential reference genes for gene expression analysis in the brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae) using reverse-transcription quantitative PCR. PLoS ONE 2014, 9, e86503. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ding, Z.; Zhang, C.; Yang, B.; Liu, Z. Gene knockdown by intro-thoracic injection of double-stranded RNA in the brown planthopper, Nilaparvata lugens. Insect Biochem. Mol. Biol. 2010, 40, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, Y.; Liu, Z.; You, L.; Wu, Y.; Xu, B.; Ge, L.; Stanley, D.; Song, Q.; Wu, J. Jinggangmycin increases fecundity of the brown planthopper, Nilaparvata lugens (Stal) via fatty acid synthase gene expression. J. Proteomics 2016, 130, 140–149. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, J.-m.; Ye, W.-f.; Noman, A.; Machado, R.A.R.; Lou, Y.-g. The Desaturase Gene Family is Crucially Required for Fatty Acid Metabolism and Survival of the Brown Planthopper, Nilaparvata lugens. Int. J. Mol. Sci. 2019, 20, 1369. https://doi.org/10.3390/ijms20061369
Zeng J-m, Ye W-f, Noman A, Machado RAR, Lou Y-g. The Desaturase Gene Family is Crucially Required for Fatty Acid Metabolism and Survival of the Brown Planthopper, Nilaparvata lugens. International Journal of Molecular Sciences. 2019; 20(6):1369. https://doi.org/10.3390/ijms20061369
Chicago/Turabian StyleZeng, Jia-mei, Wen-feng Ye, Ali Noman, Ricardo A.R. Machado, and Yong-gen Lou. 2019. "The Desaturase Gene Family is Crucially Required for Fatty Acid Metabolism and Survival of the Brown Planthopper, Nilaparvata lugens" International Journal of Molecular Sciences 20, no. 6: 1369. https://doi.org/10.3390/ijms20061369