From Ecology to Biotechnology, Study of the Defense Strategies of Algae and Halophytes (from Trapani Saltworks, NW Sicily) with a Focus on Antioxidants and Antimicrobial Properties

 , , and
, , and
Abstract
:
1. Introduction
2. Results
2.1. Polyphenol Content
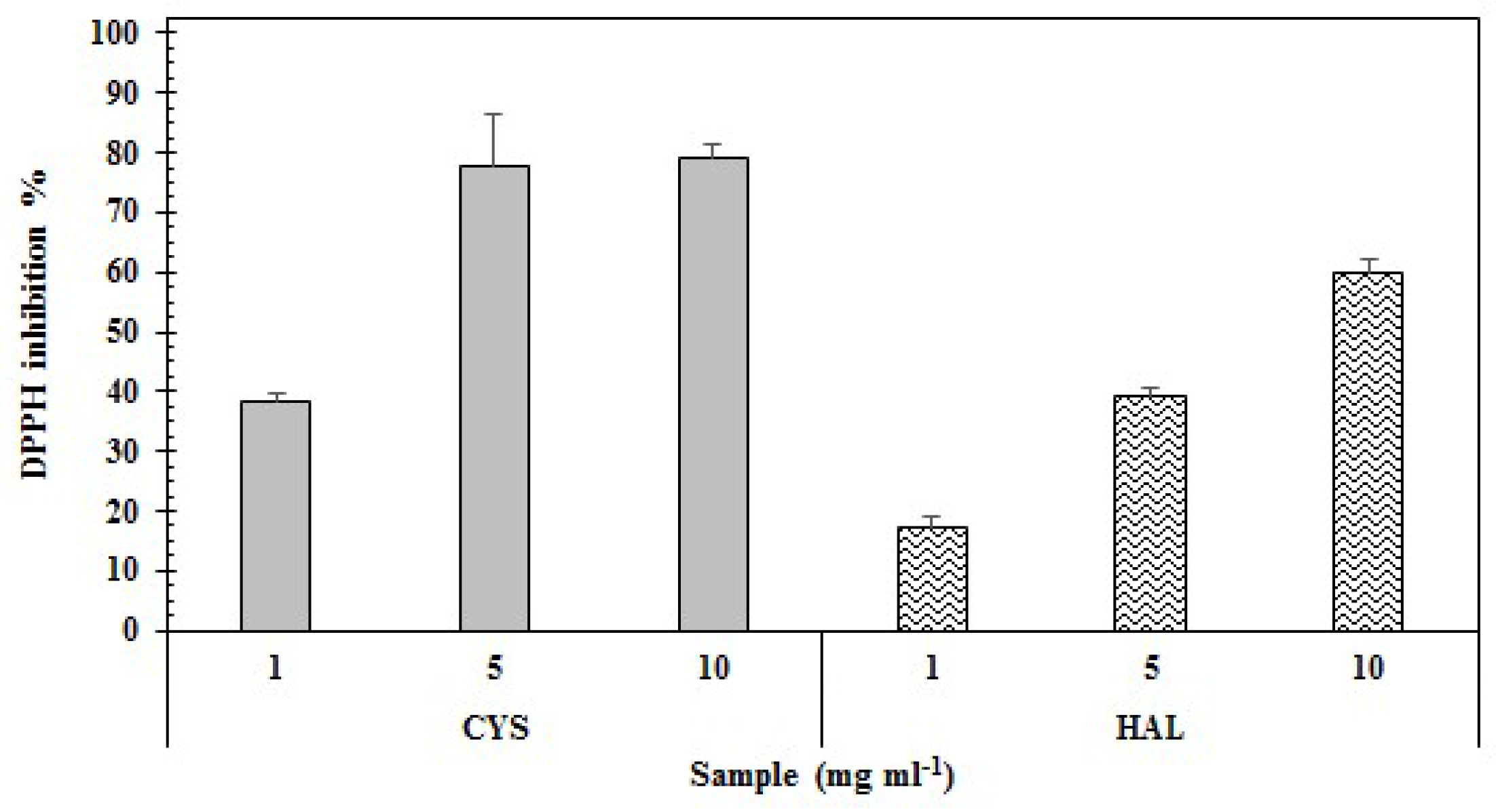
2.2. Antioxidant Properties
2.3. Antibacterial and Anti-Microalgal Properties
3. Discussion
4. Material and Methods
4.1. Sample Collections and Processing
4.2. Total Polyphenolic Content Evaluation
4.3. Antioxidant Evaluation
4.3.1. DPPH Radical-Scavenging Activity
4.3.2. Reducing Power
4.4. Antimicrobial Activity
4.4.1. Antibacterial Assays
4.4.2. Antimicroalgal Assays
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dias, D.A.; Urban, S.; Roessner, U.; Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Cobb, R.E.; Zhao, H. Recent advances in natural product discovery. Curr. Opin. Biotechnol. 2014, 30, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, S.; Abu-Ghannam, N.; Gupta, S. An assessment of the antioxidant and antimicrobial activity of six species of edible Irish seaweeds. Int. Food Res. J. 2010, 17, 205–220. [Google Scholar]
- Miri, A.; Sharifi Rad, J.; Mahsan Hoseini Alfatemi, S.; Sharifi Rad, M. A study of Antibacterial potentiality of some plants extracts against multi- drug resistant human pathogens. Ann. Biol. Res. 2013, 4, 35–41. [Google Scholar]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: some basic aspects. Mar. Ecol. Prog. Ser. 1989, 58, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Dumay, O.; Costa, J.; Desjobert, J.M.; Pergent, G. Variations in the concentration of phenolic compounds in the seagrass Posidonia oceanica under conditions of competition. Phytochemistry 2004, 65, 3211–3220. [Google Scholar] [PubMed]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gürel, A.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Jha, B.; Kavita, K.; Westphal, J.; Hartmann, A.; Schmitt-Kopplin, P. Quorum sensing inhibition by Asparagopsis taxiformis, a marine macro alga: Separation of the compound that interrupts bacterial communication. Mar. Drugs 2013, 11, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Nahas, R.; Abatis, D.; Anagnostopoulou, M.A.; Kefalas, P.; Vagias, C.; Roussis, V. Radical-scavenging activity of Aegean Sea marine algae. Food Chem. 2007, 102, 577–581. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P. Halophiles. In eLS; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1–13. ISBN 0470016175. [Google Scholar]
- Oren, A. Industrial and environmental applications of halophilic microorganisms. Environ. Technol. 2010, 31, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.M.; Bio, A.; Amat, F.; Vieira, N. Artisanal salt production in Aveiro/Portugal—An ecofriendly process. Saline Syst. 2011, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Lai, F. Nature and the city: The salt-works park in the urban area of Cagliari (Sardinia, Italy). J. Political Ecol. 2013, 20, 329–341. [Google Scholar] [CrossRef]
- Petanidou, T. The postmodern saline landscape in Greece and the European Mediterranean Salinas for salt or what? In Proceedings of the Saltworks: Preserving Saline Coastal Ecosystems—Proceedings of the Post-Conference Symposium of the 6th International Conference on Environmental Science and Technology, Samos, Greece, 30 August–2 September 1999; pp. 67–80. [Google Scholar]
- Crisman, T.L.; Takavakoglou, V.; Alexandridis, T.; Antonopoulos, V.; Zalidis, G. Rehabilitation of abandoned saltworks to maximize conservation, ecotourism and water treatment potential. Glob. NEST J. 2009, 11, 24–31. [Google Scholar]
- De Medeiros Rocha, R.; Costa, D.F.; Lucena-Filho, M.A.; Bezerra, R.M.; Medeiros, D.H.; Azevedo-Silva, A.M.; Araújo, C.N.; Xavier-Filho, L. Brazilian solar saltworks—Ancient uses and future possibilities. Aquat. Biosyst. 2012, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Thoughts on the “Missing Link” Between Saltworks Biology and Solar Salt Quality. Glob. Nest J. 2010, 12, 417–425. [Google Scholar]
- Oren, A. Salts and brines. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Dordrecht, The Netherlands, 2012; Volume 9789400738, pp. 401–426. ISBN 9789400738553. [Google Scholar]
- Petanidou, T.; Dalaka, A. Mediterranean’s changing saltscapes: A study of the abandonment of salt-making business in Greece. Glob. NEST J. 2009, 11, 415–433. [Google Scholar]
- Zeno, C. The ecological importance of the Margherita di Savoia Saltworks. Glob. NEST J. 2009, 11, 1–9. [Google Scholar]
- Santulli, A. Aquacoltura in Salina: Promozione e Valorizzazione: Definizione ed Applicazione di una Metodologia a Impatto Ridotto per L’allevamento Semi Intensivo in Saline Ricadenti All’interno di Riserva Naturale; POR Sicilia 2000/06, N°1999, IT.16.1.PO.011/4.17B/8.3.7./0063; Consorzio Universitario della Provincia di Trapani: Trapani, Italy, 2007; p. 326. [Google Scholar]
- Messina, C.M.; Santulli, A. Quality of fish reared in extensive in the salt works of Trapani and Marsala (Western Sicily). Sea salt and fish as flag products to promote territory and Natural Reserves. In Proceedings of the The Organic World Congress, Modena, Italy, 18–20 June 2008; pp. 104–106. [Google Scholar]
- Santulli, A.; Modica, A. Aquaculture in Sicily: The state of the art. Ital. J. Anim. Sci. 2009, 8, 829–838. [Google Scholar] [CrossRef]
- Viver, T.; Cifuentes, A.; Díaz, S.; Rodríguez-Valdecantos, G.; González, B.; Antón, J.; Rosselló-Móra, R. Diversity of extremely halophilic cultivable prokaryotes in Mediterranean, Atlantic and Pacific solar salterns: Evidence that unexplored sites constitute sources of cultivable novelty. Syst. Appl. Microbiol. 2015, 38, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelly, C.; Krouma, A.; Drevon, J. NoNitrogen fixation and yield of chickpea in saline Mediterranean zones. Grain Legum. 2005, 42, 16–17. [Google Scholar]
- Glenn, E.P.; Brown, J.J.; O’Leary, J.W. Irrigating Crops with Seawater. Sci. Am. 1998, 279, 76–81. [Google Scholar] [CrossRef]
- Cabioc’h, J.; Floc’h, J.Y.; Le Toquin, A.; Boudouresque, C.-F.; Meinesz, A.; Verlaque, M. Guide des Algues des Mers d’Europe; Delachaux et Niestlé: Paris, France, 2006; p. 272. [Google Scholar]
- De Los Reyes, C.; Zbakh, H.; Motilva, V.; Zubía, E. Antioxidant and anti-inflammatory meroterpenoids from the brown alga Cystoseira usneoides. J. Nat. Prod. 2013, 76, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, G.; Brullo, C.; Brullo, S.; Guarino, R.; Sciandrello, S. Studi tassonomici sulle popolazioni Italiane di Halocnemum strobilaceum (Amaranthaceae). In Societa’ Botanica Italiana, Flora Vascolare D’Italia: Studi Biosistematici Taxa endemici e loci classici; Soc. Botanica Firenze: Florence, Italy, 2012; pp. 29–31. ISBN 9788885915060. [Google Scholar]
- Pérez-Lahiguera, J.A.; Pastor-López, A.; Ferrandez-Castello, D. Is Halocnemum strobilaceum (Pall.) M. Bieb. a good flagship specie for conservation of biological diversity in coastal wetlands ecosystems? A perspective from Wadi sites in Spain, Italy and Tunisia. In Sustainable Management of Mediterranean Coastal Fres; Firenze University Press: Firenze, Italy, 2009; pp. 91–110. [Google Scholar]
- El Shaer, H.M. Halophytes and salt-tolerant plants as potential forage for ruminants in the Near East region. Small Rumin. Res. 2010, 91, 3–12. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; García-Blairsy Reina, G.; Herrero, M.; Señoráns, F.J.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirel, Z.; Yildirim, Z.D.; Tuney, I.; Kesici, K.; Sukatar, A. Biochemical Analysis of Some Brown Seaweeds from the Aegean Sea. Bot. Serbica 2012, 36, 91–95. [Google Scholar]
- Wei, R.; Lee, M.-S.; Lee, B.; Oh, C.-W.; Choi, C.-G.; Kim, H.-R. Isolation and identification of anti-inflammatory compounds from ethyl acetate fraction of Ecklonia stolonifera and their anti-inflammatory action. J. Appl. Phycol. 2016, 28, 3535–3545. [Google Scholar] [CrossRef]
- Gul, B.; Abideen, Z.; Ansari, R.; Khan, M.A. Halophytic biofuels revisited. Biofuels 2013, 4, 575–577. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, W.M.; Medini, F.; Mkadmini, K.; Legault, J.; Magné, C.; Abdelly, C.; Ksouri, R. LC–ESI-TOF–MS identification of bioactive secondary metabolites involved in the antioxidant, anti-inflammatory and anticancer activities of the edible halophyte Zygophyllum album Desf. Food Chem. 2013, 139, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, S.; Mathew, K.T.; Gray, A.I. A caffeic acid ester from Halocnemum strobilaceum. Phytochemistry 1999, 51, 465–467. [Google Scholar] [CrossRef]
- Miftakhova, A.F.; Burasheva, G.S.; Abilov, Z.A. Flavonoids of Halocnemum strobilaceum. Chem. Nat. Compd. 1999, 35, 100–101. [Google Scholar] [CrossRef]
- Miftakhova, A.F.; Burasheva, G.S.; Abilov, Z.A.; Ahmad, V.U.; Zahid, M. Coumarins from the aerial part of Halocnemum strobilaceum. Fitoterapia 2001, 72, 319–321. [Google Scholar] [CrossRef]
- Radwan, H.; Shams, K. Phytochemical and biological investigations on Halocnemum strobilaceum (Chenopodiaceae). Planta Med. 2007, 73, 337. [Google Scholar] [CrossRef]
- Rodil, I.F.; Lastra, M. Environmental factors affecting benthic macrofauna along a gradient of intermediate sandy beaches in northern Spain. Estuar. Coast. Shelf Sci. 2004, 61, 37–44. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Coetser, S.E.; Cloete, T.E. Biofouling and biocorrosion in industrial water systems. Crit. Rev. Microbiol. 2005, 31, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.M.; Lodge, D.M. Hull fouling is a risk factor for intercontinental species exchange in aquatic ecosystems. Aquat. Invasions 2007, 2, 121–131. [Google Scholar] [CrossRef]
- Hellio, C.; Bremer, G.; Pons, A.M.; Le Gal, Y.; Bourgougnon, N. Inhibition of the development of microorganisms (bacteria and fungi) by extracts of marine algae from Brittany, France. Appl. Microbiol. Biotechnol. 2000, 54, 543–549. [Google Scholar] [CrossRef]
- Maréchal, J.P.; Hellio, C. Challenges for the development of new non-toxic antifouling solutions. Int. J. Mol. Sci. 2009, 10, 4623–4637. [Google Scholar] [CrossRef]
- Falaye, B.J.; Oyewumi, K.J. Solutions of the Dirac equation with spin and pseudospin symmetry for the trigonometric scarf potential in D-dimensions. Afr. Rev. Phys. 2011, 6, 211–220. [Google Scholar]
- Bettin, C.; Oehlmann, J.; Stroben, E. TBT-induced imposex in marine neogastropods is mediated by an increasing androgen level. Helgoländer Meeresuntersuchungen 1996, 50, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Oehlmann, U.; Oehlmann, J.; Fioroni, P.; Bauer, B. Imposex and reproductive failure in Hydrobia ulvae (Gastropoda: Prosobranchia). Mar. Biol. 1997, 128, 257–266. [Google Scholar] [CrossRef]
- Alzieu, C. Environmental impact of TBT: The French experience. Sci. Total Environ. 2000, 258, 99–102. [Google Scholar] [CrossRef]
- IMO. International Convention on the Control of Harmful Anti-Fouling Systems on Ships; IMO: London, UK, 2001. [Google Scholar]
- Quintã, R.; Santos, R.; Thomas, D.N.; Le Vay, L. Growth and nitrogen uptake by Salicornia europaea and Aster tripolium in nutrient conditions typical of aquaculture wastewater. Chemosphere 2015, 120, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, O.R.; de MB Borin, M.R. Medicinal Products: Regulation of Biosynthesis in Space and Time. Mem. Inst. Oswaldo Cruz 2000, 95, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Schoenwaelder, M.E.A. The Biology of Phenolic Containing Vesicles. ALGAE 2008, 23, 163–175. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Lagrimini, L.M. Wound-induced deposition of polyphenols in transgenic plants overexpressing peroxidase. Plant Physiol. 1991, 96, 577–583. [Google Scholar] [CrossRef]
- Mathiesen, L.; Malterud, K.E.; Sund, R.B. Hydrogen bond formation as basis for radical scavenging activity: A structure-activity study of C-methylated dihydrochalcones from Myrica gale and structurally related acetophenones. Free Radic. Biol. Med. 1997, 22, 307–311. [Google Scholar] [CrossRef]
- Blainski, A.; Lopes, G.C.; De Mello, J.C.P. Application and analysis of the Folin ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Gharbi, S.; Renda, G.; La Barbera, L.; Amri, M.; Messina, C.M.; Santulli, A. Tunisian tomato by-products, as a potential source of natural bioactive compounds. Nat. Prod. Res. 2017, 31, 626–631. [Google Scholar] [CrossRef]
- Manuguerra, S.; Caccamo, L.; Mancuso, M.; Arena, R.; Rappazzo, A.C.; Genovese, L.; Santulli, A.; Messina, C.M.; Maricchiolo, G. The antioxidant power of horseradish, Armoracia rusticana, underlies antimicrobial and antiradical effects, exerted in vitro. Nat. Prod. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Council of Europe. Determination of tannins in herbal drugs. In European Pharmacopoeia, 6th ed.; European Directorate for the Quality of Medicines: Strasbourg, France, 2007; p. A286. [Google Scholar]
- Farmacopeia Brasileira, 5th ed.; Agência Nacional de Vigilância: Brasília, Brazil, 2010.
- Schofield, P.; Mbugua, D.M.; Pell, A.N. Analysis of condensed tannins: A review. Anim. Feed Sci. Technol. 2001, 91, 21–40. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On Tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, LXXIII, 627–650. [Google Scholar]
- Glasl, H. Zur Photometrie in der Drogenstandardisierung. 3. Gehaltsbestimmung von Gerbstoffdrogen. Dtsch Apoth Ztg 1983, 123, 1979–1983. [Google Scholar]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef]
- Messina, C.; Renda, G.; Randazzo, M.; Laudicella, A.; Gharbi, S.; Pizzo, F.; Morghese, M.; Santulli, A. Extraction of bioactive compounds from shrimp waste. Bull. Inst. Natl. Sci. Technol. Mer Salammbô 2015, 42, 2014–2016. [Google Scholar]
- Tajbakhsh, S.; Ilkhani, M.; Rustaiyan, A.; Larijani, K.; Sartavi, K.; Tahmasebi, R.; Asayesh, G. Antibacterial effect of the brown alga Cystoseira trinodis. J. Med. Plants Res. 2011, 5, 4654–4657. [Google Scholar]
- Mhadhebi, L.; Chaieb, K.; Bouraoui, A. Evaluation of antimicrobial activity of organic fractions of six marine algae from tunisian mediterranean coasts. Int. J. Pharm. Pharm. Sci. 2012, 4, 534–537. [Google Scholar]
- Bennamara, A.; Abourriche, A.; Berrada, M.; Charrouf, M.; Chaib, N.; Boudouma, M.; Garneau, F.X. Methoxybifurcarenone: An antifungal and antibacterial meroditerpenoid from the brown alga Cystoseira tamariscifolia. Phytochemistry 1999, 52, 37–40. [Google Scholar] [CrossRef]
- Zineb, S.; Mohamed, L.; Mohamed, F.; Khadija, F. Inhibition of growth and mycotoxins formation in moulds by marine algae Cystoseira tamariscifolia. J Biotechnol 2004, 3, 71–75. [Google Scholar]
- Ruberto, G.; Baratta, M.T.; Biondi, D.M.; Amico, V. Antioxidant activity of extracts of the marine algal genus Cystoseira in a micellar model system. J. Appl. Phycol. 2001, 13, 403–407. [Google Scholar] [CrossRef]
- Chkhikvishvili, I.D.; Ramazanov, Z.M. Phenolic substances of brown algae and their antioxidant activity. Appl. Biochem. Microbiol. 2000, 36, 289–291. [Google Scholar] [CrossRef]
- Mhadhebi, L.; Mhadhebi, A.; Robert, J.; Bouraoui, A. Antioxidant, anti-inflammatory and antiproliferative effects of aqueous extracts of three mediterranean brown seaweeds of the Genus Cystoseira. Iran. J. Pharm. Res. 2014, 13, 207–220. [Google Scholar] [PubMed]
- Turan, F.; Ozgun, S.; Sayın, S.; Ozyılmaz, G. Biochemical composition of some red and green seaweeds from Iskenderun Bay, the northeastern Mediterranean coast of Turkey. J. Black Sea/Mediterranean Environ. 2015, 21, 239–249. [Google Scholar]
- Abdala-Díaz, R.T.; Cabello-Pasini, A.; Pérez-Rodríguez, E.; Conde Álvarez, R.M.; Figueroa, F.L. Daily and seasonal variations of optimum quantum yield and phenolic compounds in Cystoseira tamariscifolia (Phaeophyta). Mar. Biol. 2006, 148, 459–465. [Google Scholar] [CrossRef]
- Betancor, S.; Domínguez, B.; Tuya, F.; Figueroa, F.L.; Haroun, R. Photosynthetic performance and photoprotection of Cystoseira humilis (Phaeophyceae) and Digenea simplex (Rhodophyceae) in an intertidal rock pool. Aquat. Bot. 2015, 121, 16–25. [Google Scholar] [CrossRef]
- Connan, S.; Stengel, D.B. Impacts of ambient salinity and copper on brown algae: 2. Interactive effects on phenolic pool and assessment of metal binding capacity of phlorotannin. Aquat. Toxicol. 2011, 104, 1–13. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative Stress in Marine Environments: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Ksouri, R.; Smaoui, A.; Isoda, H.; Abdelly, C. Utilization of halophyte species as new sources of bioactive substances. J. Arid Land Stud. 2012, 22, 41–44. [Google Scholar]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Messina, C.M.; Bono, G.; Renda, G.; La Barbera, L.; Santulli, A. Effect of natural antioxidants and modified atmosphere packaging in preventing lipid oxidation and increasing the shelf-life of common dolphinfish (Coryphaena hippurus) fillets. LWT Food Sci. Technol. 2015, 62, 271–277. [Google Scholar] [CrossRef]
- Trabelsi, N.; Megdiche, W.; Ksouri, R.; Falleh, H.; Oueslati, S.; Soumaya, B.; Hajlaoui, H.; Abdelly, C. Solvent effects on phenolic contents and biological activities of the halophyte Limoniastrum monopetalum leaves. LWT Food Sci. Technol. 2010, 43, 632–639. [Google Scholar] [CrossRef]
- Banerjee, D.; Chakrabarti, S.; Hazra, A.K.; Banerjee, S.; Ray, J.; Mukherjee, B. Antioxidant activity and total phenolics of some mangroves in Sundarbans. Afr. J. Biotechnol. 2008, 7, 805–810. [Google Scholar]
- Hanen, F.; Riadh, K.; Samia, O.; Sylvain, G.; Christian, M.; Chedly, A. Interspecific variability of antioxidant activities and phenolic composition in Mesembryanthemum genus. Food Chem. Toxicol. 2009, 47, 2308–2313. [Google Scholar] [CrossRef]
- Siracusa, L.; Kulisic-Bilusic, T.; Politeo, O.; Krause, I.; Dejanovic, B.; Ruberto, G. Phenolic composition and antioxidant activity of aqueous infusions from Capparis spinosa L. and Crithmum maritimum L. before and after submission to a two-step in vitro digestion model. J. Agric. Food Chem. 2011, 59, 12453–12459. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Le Floch, G.; Magné, C. Radical scavenging, antioxidant and antimicrobial activities of halophytic species. J. Ethnopharmacol. 2008, 116, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Aoun, Z.B.; Said, R.B.; Farhat, F. Anti-inflammatory, antioxidant and antimicrobial activities of aqueous and organic extracts from Dictyopteris membranacea. Bot. Mar. 2010, 53, 259–264. [Google Scholar] [CrossRef]
- Alger, M.S.M. Polymer Science Dictionary; Springer: Berlin, Germany, 1997. [Google Scholar]
- Duan, X.J.; Zhang, W.W.; Li, X.M.; Wang, B.G. Evaluation of antioxidant property of extract and fractions obtained from a red alga, Polysiphonia urceolata. Food Chem. 2006, 95, 37–43. [Google Scholar] [CrossRef]
- Yen, G.-C.; Chen, H.-Y. Antioxidant Activity of Various Tea Extract in Their Antimutagenicity. J. Agric. Food Chem. 1995, 43, 27–32. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Amić, D.; Davidović-Amić, D.; Drago, B.; Trinajstić, N. Structure-Radical Scavenging Activity Relationships of Flavonoids Dragan. Croat. Chem. Acta 2003, 76, 55–61. [Google Scholar]
- Sellimi, S.; Younes, I.; Ayed, H.B.; Maalej, H.; Montero, V.; Rinaudo, M.; Dahia, M.; Mechichi, T.; Hajji, M.; Nasri, M. Structural, physicochemical and antioxidant properties of sodium alginate isolated from a Tunisian brown seaweed. Int. J. Biol. Macromol. 2015, 72, 1358–1367. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nagayama, K.; Uchida, K.; Tanaka, R. Antioxidant Activity of Phlorotannins Isolated from the Brown Alga Eisenia bicyclis. Fish. Sci. 1996, 62, 923–926. [Google Scholar] [CrossRef]
- Da Gama, B.A.P.; Plouguerné, E.; Pereira, R.C. The antifouling defence mechanisms of marine macroalgae. Adv. Bot. Res. 2014, 71, 413–440. [Google Scholar]
- Jacobson, A.H.; Willingham, G.L. Sea-nine antifoulant: An environmentally acceptable alternative to organotin antifoulants. Sci. Total Environ. 2000, 258, 103–110. [Google Scholar] [CrossRef]
- García-Bueno, N.; Decottignies, P.; Turpin, V.; Dumay, J.; Paillard, C.; Stiger-Pouvreau, V.; Kervarec, N.; Pouchus, Y.-F.; Marín-Atucha, A.A.; Fleurence, J. Seasonal antibacterial activity of two red seaweeds, Palmaria palmata and Grateloupia turuturu, on European abalone pathogen Vibrio harveyi. Aquat. Living Resour. 2014, 27, 83–89. [Google Scholar] [CrossRef]
- Lee, K.K.; Liu, P.C.; Chuang, W.H. Pathogenesis of gastroenteritis caused by Vibrio carchariae in cultured marine fish. Marine Biotechnol. 2002, 4, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Vandenberghe, J.; Thompson, F.L.; Gomez-Gil, B.; Swings, J. Phenotypic diversity amongst Vibrio isolates from marine aquaculture systems. Aquaculture 2003, 219, 9–20. [Google Scholar] [CrossRef]
- Benbouzid-Rollet, N.D.; Conte, M.; Guezennec, J.; Prieur, D. Monitoring of a Vibrio natriegens and Desulfovibrio vulgaris marine aerobic biofilm on a stainless steel surface in a laboratory tubular flow system. J. Appl. Bacteriol. 1991, 71, 244–251. [Google Scholar] [CrossRef]
- Cheng, G.; Li, G.; Xue, H.; Chen, S.; Bryers, J.D.; Jiang, S. Zwitterionic carboxybetaine polymer surfaces and their resistance to long-term biofilm formation. Biomaterials 2009, 30, 5234–5240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosink, J.J.; Woese, C.R.; Staley, J.T. Polaribacter gen. nov., with three new species, P. irgensii sp. nov., P. franzmannii sp. nov. and P. filamentus sp. nov., gas vacuolate polar marine bacteria of the Cytophaga-Flavobacterium-Bacteroides group and reclassification of Flectobacillus glomera. Int. J. Syst. Bacteriol. 1998, 48, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Jennings, J.G.; Steinberg, P.D. Phlorotannins versus other factors affecting epiphyte abundance on the kelp Ecklonia radiata. Oecologia 1997, 109, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Hellio, C.; Marechal, J.P.; Véron, B.; Bremer, G.; Clare, A.S.; Le Gal, Y. Seasonal variation of antifouling activities of marine algae from the Brittany Coast (France). Mar. Biotechnol. 2004, 6, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Husain, F.M.; Ahmad, I. Marine organisms as source of quorum sensing inhibitors. In Quorum Sensing vs Quorum Quenching: A Battle with No End in Sight; Springer: New Delhi, India, 2015; pp. 259–268. ISBN 9788132219828. [Google Scholar]
- Hellio, C.; De La Broise, D.; Dufossé, L.; Le Gal, Y.; Bourgougnon, N. Inhibition of marine bacteria by extracts of macroalgae: Potential use for environmentally friendly antifouling paints. Mar. Environ. Res. 2001, 52, 231–247. [Google Scholar] [CrossRef]
- Maréchal, J.P.; Culioli, G.; Hellio, C.; Thomas-Guyon, H.; Callow, M.E.; Clare, A.S.; Ortalo-Magné, A. Seasonal variation in antifouling activity of crude extracts of the brown alga Bifurcaria bifurcata (Cystoseiraceae) against cyprids of Balanus amphitrite and the marine bacteria Cobetia marina and Pseudoalteromonas haloplanktis. J. Exp. Mar. Biol. Ecol. 2004, 313, 47–62. [Google Scholar] [CrossRef]
- Thabard, M.; Gros, O.; Hellio, C.; Maréchal, J.P. Sargassum polyceratium (Phaeophyceae, Fucaceae) surface molecule activity towards fouling organisms and embryonic development of benthic species. Bot. Mar. 2011, 54, 147–157. [Google Scholar] [CrossRef]
- Plouguerné, E.; Hellio, C.; Cesconetto, C.; Thabard, M.; Mason, K.; Véron, B.; Pereira, R.C.; da Gama, B.A.P. Antifouling activity as a function of population variation in Sargassum vulgare from the littoral of Rio de Janeiro (Brazil). J. Appl. Phycol. 2010, 22, 717–724. [Google Scholar] [CrossRef]
- Águila-Ramírez, R.N.; Arenas-gonzález, A.; Hernández-guerrero, C.J.; González-acosta, B.; Borges-souza, J.M.; Véron, B.; Pope, J.; Hellio, C. Antimicrobial and antifouling activities achieved by extracts of seaweeds from Gulf of California, Mexico Actividades antimicrobiana y anti-incrustante obtenidas de los extractos de algas marinas del Golfo de California, México. Hidrobiologica 2012, 22, 8–15. [Google Scholar]
- Silkina, A.; Bazes, A.; Vouvé, F.; Le Tilly, V.; Douzenel, P.; Mouget, J.L.; Bourgougnon, N. Antifouling activity of macroalgal extracts on Fragilaria pinnata (Bacillariophyceae): A comparison with Diuron. Aquat. Toxicol. 2009, 94, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Bazes, A.; Silkina, A.; Defer, D.; Bernède-Bauduin, C.; Quéméner, E.; Braud, J.P.; Bourgougnon, N. Active substances from Ceramium botryocarpum used as antifouling products in aquaculture. Aquaculture 2006, 258, 664–674. [Google Scholar] [CrossRef]
- Piazza, V.; Roussis, V.; Garaventa, F.; Greco, G.; Smyrniotopoulos, V.; Vagias, C.; Faimali, M. Terpenes from the Red Alga Sphaerococcus coronopifolius Inhibit the Settlement of Barnacles. Mar. Biotechnol. 2011, 13, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Sawabe, T.; Gacesa, P.; Ezura, Y. Rapid PCR Detection of Pseudoalteromonas Elyakovii, the Causative Bacterium of Laminaria Spot Wound Disease in Japan. In Proceedings of the Seventeenth International Seaweed Symposium, Cape Town, South Africa, 28 January–2 February 2001; Chapman, A.R.O., Anderson, R.J., Vreeland, V.J., Davison, I.R., Eds.; Oxford University Press: London, UK, 2001; pp. 389–394. [Google Scholar]
- Qurashi, A.W.; Sabri, A.N. Alleviation of salt stress by Halomonas sp. and osmolytes in Zea mays. AFRICAN J. Biotechnol. 2011, 10, 17778–17788. [Google Scholar]
- Hartley, M.; Bennett, B. Heterologous expression and purification of Vibrio proteolyticus (Aeromonas proteolytica) aminopeptidase: A rapid protocol. Protein Expr. Purif. 2009, 66, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Buchan, A.; González, J.M.; Moran, M.A. Overview of the Marine Roseobacter Lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef] [Green Version]
- Sharifah, E.N.; Eguchi, M. The Phytoplankton Nannochloropsis oculata Enhances the Ability of Roseobacter Clade Bacteria to Inhibit the Growth of Fish Pathogen Vibrio anguillarum. PLoS ONE 2011, 6, e26756. [Google Scholar] [CrossRef]
- Ksouri, R.; Falleh, H.; Megdiche, W.; Trabelsi, N.; Mhamdi, B.; Chaieb, K.; Bakrouf, A.; Magné, C.; Abdelly, C. Antioxidant and antimicrobial activities of the edible medicinal halophyte Tamarix gallica L. and related polyphenolic constituents. Food Chem. Toxicol. 2009, 47, 2083–2091. [Google Scholar] [CrossRef]
- Meot-Duros, L.; Cérantola, S.; Talarmin, H.; Le Meur, C.; Le Floch, G.; Magné, C. New antibacterial and cytotoxic activities of falcarindiol isolated in Crithmum maritimum L. leaf extract. Food Chem. Toxicol. 2010, 48, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Mahsan Hoseini Alfatemi, S.; Sharifi-Rad, M.; Sharifi Rad, J.; Sharifi Rad, M. In vitro assessment of antibacterial activity of Salicornia herbacea L. seed extracts against multidrug resistant gram-positive and gram-negative bacteria. Int. J. Biosci 2014, 4, 217–222. [Google Scholar]
- Deepa, S.; Srikumar, M.; Padmakumar, K. Antifouling potential of selected mangroves from Indian Coast. Nat. Prod. Res. 2014, 4, 54–57. [Google Scholar]
- Manilal, A.; Sujith, S.; Kiran, G.S.; Selvin, J.; Shakir, C. Biopotentials of Mangroves Collected from the Southwest Coast of India. Global J. Biotechnol. Biochem. 2009, 4, 59–65. [Google Scholar]
- Kong, N.N.; Fang, S.T.; Liu, Y.; Wang, J.H.; Yang, C.Y.; Xia, C.H. Flavonoids from the halophyte Apocynum venetum and their antifouling activities against marine biofilm-derived bacteria. Nat. Prod. Res. 2014, 28, 928–931. [Google Scholar] [CrossRef]
- Akarpat, A.; Turhan, S.; Ustun, N.S. Effects of hot-water extracts from Myrtle, Rosemary, Nettle and Lemon balm leaves on lipid oxidation and color of beef patties during frozen storage. J. Food Process. Preserv. 2008, 32, 117–132. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Kuda, T.; Hishi, T.; Maekawa, S. Antioxidant properties of dried product of “haba-nori”, an edible brown alga, Petalonia binghamiae (J. Agaradh) Vinogradova. Food Chem. 2006, 98, 545–550. [Google Scholar] [CrossRef]
- Trepos, R.; Cervin, G.; Pile, C.; Pavia, H.; Hellio, C.; Svenson, J. Evaluation of cationic micropeptides derived from the innate immune system as inhibitors of marine biofouling. Biofouling 2015, 31, 393–403. [Google Scholar] [CrossRef]
- Chapman, J.; Hellio, C.; Sullivan, T.; Brown, R.; Russell, S.; Kiterringham, E.; Le Nor, L.; Regan, F. Bioinspired synthetic macroalgae: Examples from nature for antifouling applications. Int. Biodeterior. Biodegrad. 2014, 86, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Kesarcodi-Watson, A.; Kaspar, H.; Lategan, M.J.; Gibson, L. Probiotics in aquaculture: The need, principles and mechanisms of action and screening processes. Aquaculture 2008, 274, 1–14. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antioxidant Properties | CYS | HAL | |
|---|---|---|---|
| DPPH | Inhibition (%) | 79.30 ± 1.27 | 59.90 ± 6.74 |
| EC50 (mg·mL−1) | 5.27 | 6.9 | |
| Reducing power | Reduction (%) | 93.91 ± 4.34 | 90.03 ± 6.19 |
| EC50 (mg·mL−1) | 3.59 | 4.07 | |
| MIC (µg·mL−1) | |||||||
|---|---|---|---|---|---|---|---|
| Species | Code | Growth | Adhesion | ||||
| Marine Bacteria | ATCC 1 | CYS | HAL | SEA-NINE™ 211N’ | CYS | HAL | SEA-NINE™ 211N’ |
| Polaribacter irgensii | 700398 | 0.005 | >10 | 1 | >10 | >10 | 0.1 |
| Halomonas aquamarina | 14400 | >10 | 0.0001 | 0.1 | >10 | >10 | <0.01 |
| Pseudoalteromonas elyakovii | 700519 | >10 | 0.00001 | 0.1 | >10 | >10 | <0.01 |
| Roseobacter litoralis | 49566 | >10 | 0.0001 | 1 | >10 | >10 | <0.01 |
| Shewanella putrefaciens | 8071 | >10 | >10 | 1 | >10 | >10 | 1 |
| Vibrio aesturianus | 35048 | 0.005 | 0.01 | <0.01 | >10 | >10 | 1 |
| Vibrio carchariae | 35084 | 0.005 | >10 | <0.01 | >10 | >10 | <0.01 |
| Vibrio harveyi | 700106 | 0.005 | >10 | 1 | >10 | >10 | 1 |
| Vibrio natriegens | 14058 | 0.01 | >10 | 1 | >10 | >10 | <0.01 |
| Vibrio proteolyticus | 53559 | >10 | 0.01 | 0.01 | >10 | >10 | 0.01 |
| Microalgae strains | Algobank Code | ||||||
| Exanthemachrysis gayraliae | AC 15 | 10 | >10 | <0.01 | 0.001 | >10 | <0.01 |
| Cylindrotheca closterium | AC 170 | 10 | >10 | <0.01 | 1 | >10 | <0.01 |
| Porphyridium purpureum | AC 122 | >10 | >10 | <0.01 | 10 | >10 | <0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messina, C.M.; Renda, G.; Laudicella, V.A.; Trepos, R.; Fauchon, M.; Hellio, C.; Santulli, A. From Ecology to Biotechnology, Study of the Defense Strategies of Algae and Halophytes (from Trapani Saltworks, NW Sicily) with a Focus on Antioxidants and Antimicrobial Properties. Int. J. Mol. Sci. 2019, 20, 881. https://doi.org/10.3390/ijms20040881
Messina CM, Renda G, Laudicella VA, Trepos R, Fauchon M, Hellio C, Santulli A. From Ecology to Biotechnology, Study of the Defense Strategies of Algae and Halophytes (from Trapani Saltworks, NW Sicily) with a Focus on Antioxidants and Antimicrobial Properties. International Journal of Molecular Sciences. 2019; 20(4):881. https://doi.org/10.3390/ijms20040881
Chicago/Turabian StyleMessina, Concetta Maria, Giuseppe Renda, Vincenzo Alessandro Laudicella, Rozenn Trepos, Marilyne Fauchon, Claire Hellio, and Andrea Santulli. 2019. "From Ecology to Biotechnology, Study of the Defense Strategies of Algae and Halophytes (from Trapani Saltworks, NW Sicily) with a Focus on Antioxidants and Antimicrobial Properties" International Journal of Molecular Sciences 20, no. 4: 881. https://doi.org/10.3390/ijms20040881