Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing


 ,
,  , and
, and
Abstract
:
1. Introduction
2. Results
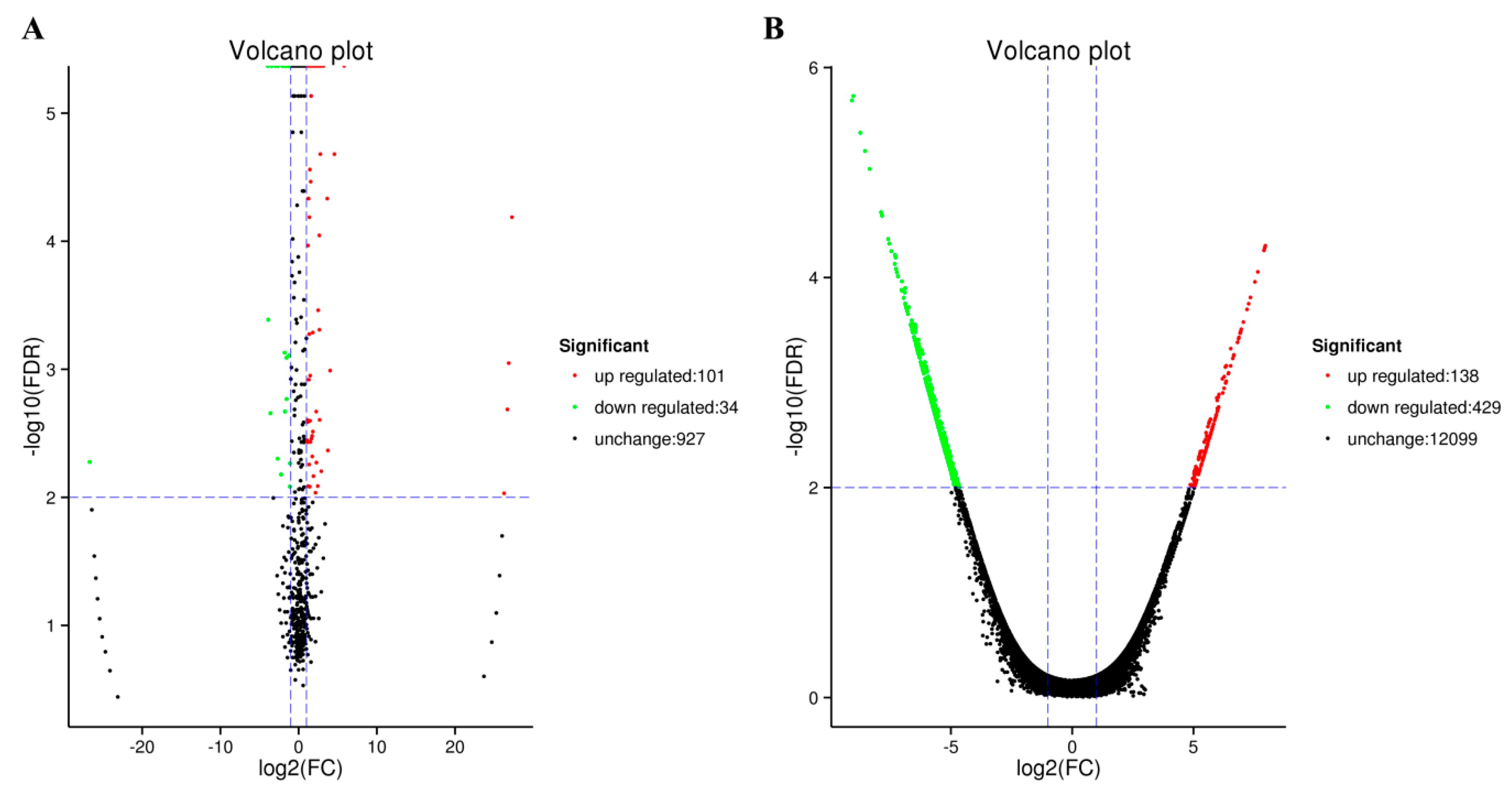
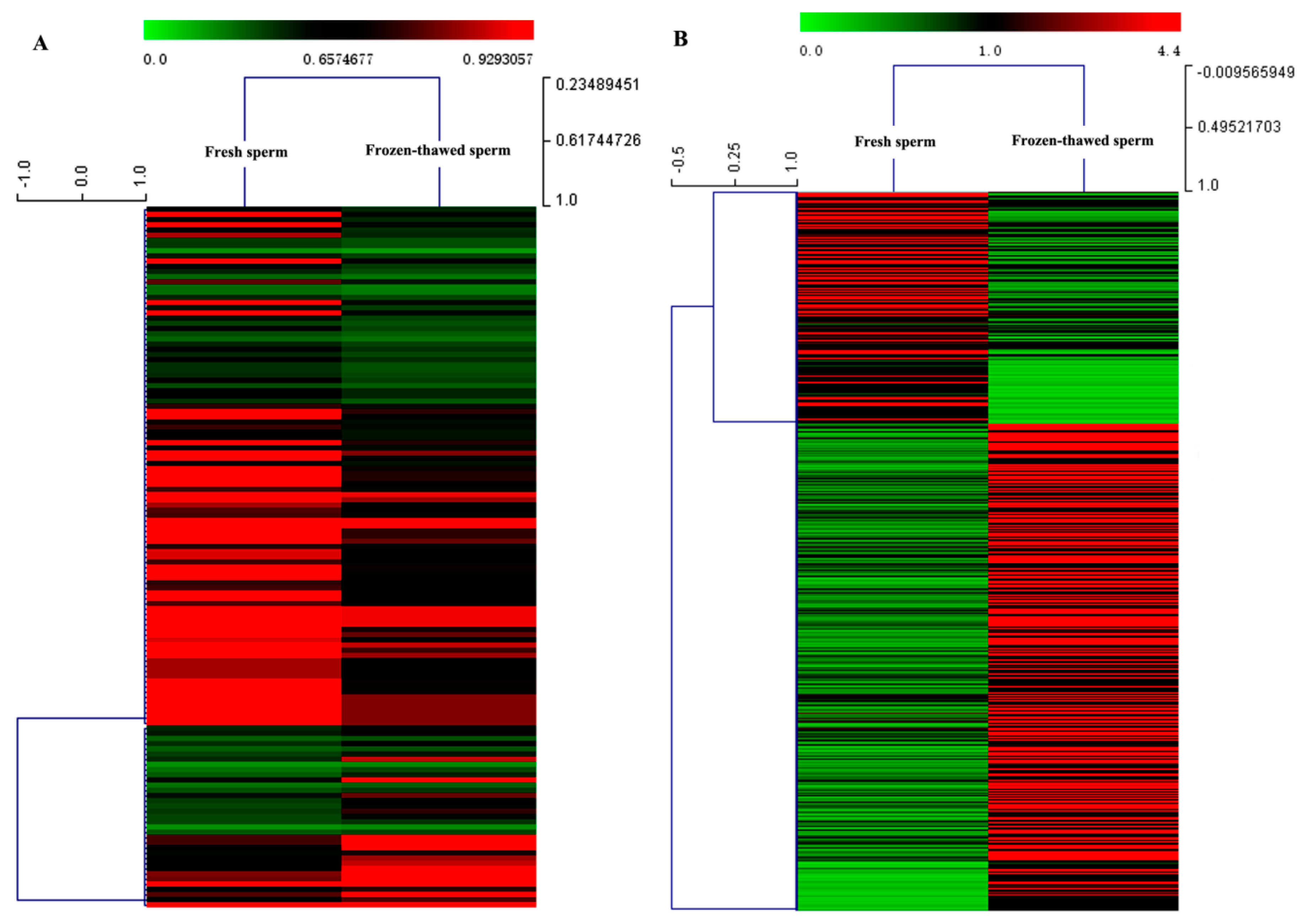
2.1. Analysis of Small RNA Sequences
2.2. Analysis of Transcriptome Sequences
2.3. Combined Analysis of Transcriptome and Small RNA Sequencing
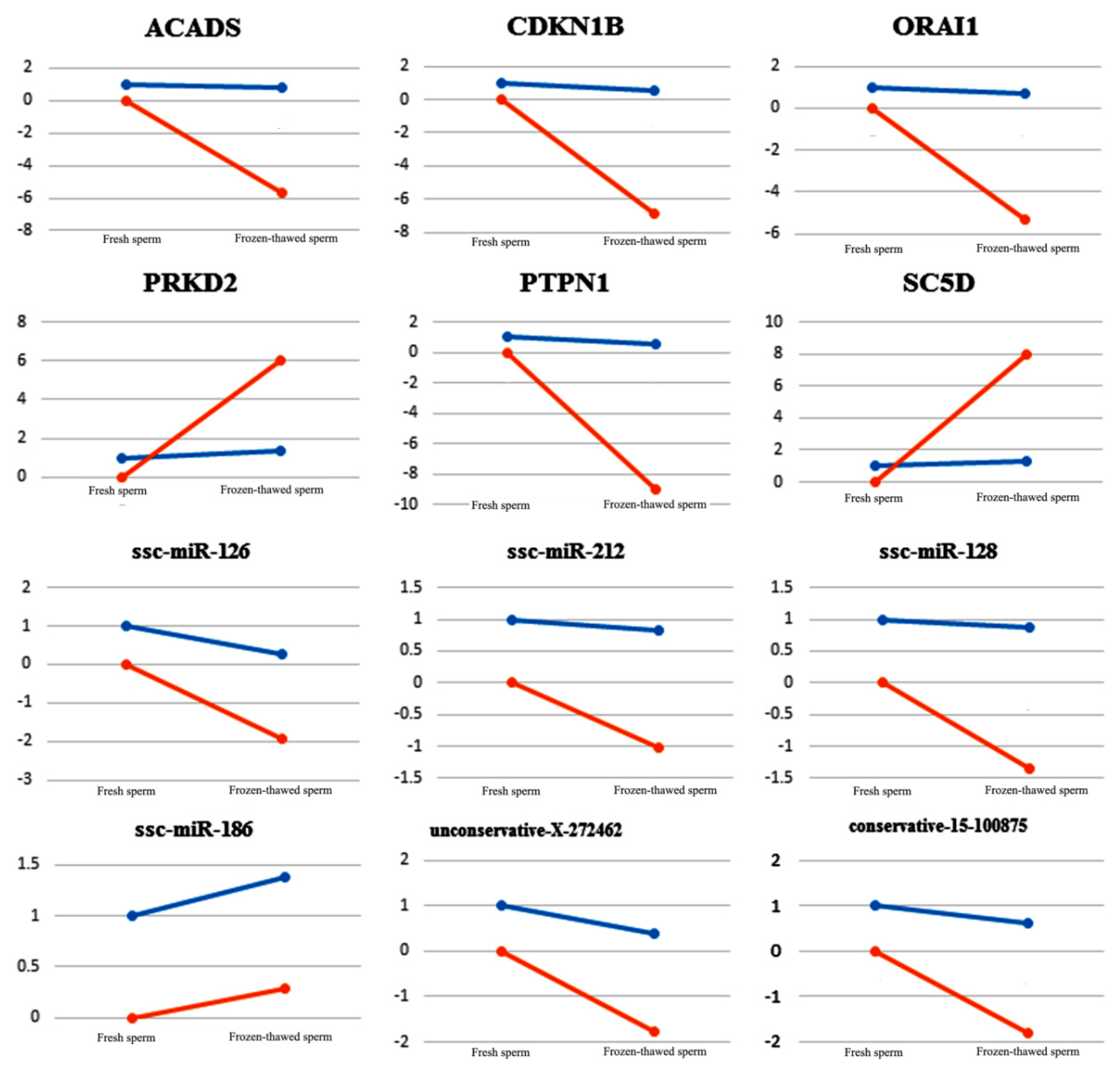
2.4. QRT-PCR Validation
3. Discussion
4. Materials and Methods
4.1. Semen Collection
4.2. Semen Cryopreservation
4.3. RNA Preparation and Small RNA Sequencing
4.4. Identification of Known/Novel miRNAs and Target Gene Prediction
4.5. Transcriptome Library Construction, Sequencing and Analysis
4.6. Quantitative Real-Time Reverse Transcription PCR (qRT-PCR) Validation of Differentially Expressed mRNAs and miRNAs
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bailey, J.L.; Lessard, C.; Jacques, J.; Brèque, C.; Dobrinski, I.; Zeng, W.; Galantino-Homer, H.L. Cryopreservation of boar semen and its future importance to the industry. Theriogenology 2008, 70, 1251–1259. [Google Scholar] [CrossRef]
- Yeste, M. Recent advances in boar sperm cryopreservation: State of the art and current perspectives. Reprod. Domest. Anim. 2015, 50, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, Y.-J.; Kim, Y.-J. Changes in sperm membrane and ROS following cryopreservation of liquid boar semen stored at 15 C. Anim. Reprod. Sci. 2011, 124, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2016, 85, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P.; Leibo, S.; Seidel, G.E., Jr. Cryopreservation of the germplasm of animals used in biological and medical research: Importance, impact, status, and future directions. Biol. Reprod. 2008, 78, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Kopeika, J.; Thornhill, A.; Khalaf, Y. The effect of cryopreservation on the genome of gametes and embryos: Principles of cryobiology and critical appraisal of the evidence. Hum. Reprod. Update 2015, 21, 209–227. [Google Scholar] [CrossRef] [PubMed]
- O’connell, M.; McClure, N.; Lewis, S. The effects of cryopreservation on sperm morphology, motility and mitochondrial function. Hum. Reprod. 2002, 17, 704–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tongdee, P.; Sukprasert, M.; Satirapod, C.; Wongkularb, A.; Choktanasiri, W. Comparison of cryopreserved human sperm between ultra rapid freezing and slow programmable freezing: Effect on motility, morphology and DNA integrity. J. Med. Assoc. Thail. 2015, 98, S33–S42. [Google Scholar]
- Bailey, J.L.; BLODEAU, J.F.; CORMIER, N. Semen cryopreservation in domestic animals: A damaging and capacitating phenomenon minireview. J. Androl. 2000, 21, 1–7. [Google Scholar] [PubMed]
- Bilodeau, J.F.; Chatterjee, S.; Sirard, M.A.; Gagnon, C. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing and thawing. Mol. Reprod. Dev. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Kim, S.-H.; Yu, D.-H.; Kim, Y.-J. Effects of cryopreservation on phosphatidylserine translocation, intracellular hydrogen peroxide, and DNA integrity in canine sperm. Theriogenology 2010, 73, 282–292. [Google Scholar] [CrossRef]
- Gürler, H.; Malama, E.; Heppelmann, M.; Calisici, O.; Leiding, C.; Kastelic, J.; Bollwein, H. Effects of cryopreservation on sperm viability, synthesis of reactive oxygen species, and DNA damage of bovine sperm. Theriogenology 2016, 86, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Tuncer, P.B.; Sariozkan, S.; Baspinar, N.; Taspinar, M.; Coyan, K.; Bilgili, A.; Akalin, P.P.; Buyukleblebici, S.; Aydos, S.; et al. Effects of antioxidants on post-thawed bovine sperm and oxidative stress parameters: Antioxidants protect DNA integrity against cryodamage. Cryobiology 2010, 61, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.; Welch, G. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.; Welch, G. Effects of reactive oxygen species on sperm function. Theriogenology 2012, 78, 1700–1708. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Alborcia, M.J.; Valverde, A.; Parrilla, I.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Detrimental effects of non-functional spermatozoa on the freezability of functional spermatozoa from boar ejaculate. PLoS ONE 2012, 7, e36550. [Google Scholar] [CrossRef]
- Cross, N.L. Role of cholesterol in sperm capacitation. Biol. Reprod. 1998, 59, 7–11. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Althouse, G.C. Characterization of capacitation, cryoinjury, and the role of seminal plasma in porcine sperm. Theriogenology 2011, 76, 1508–1516. [Google Scholar] [CrossRef]
- Buhr, M.; Curtis, E.; Kakuda, N.S. Composition and behavior of head membrane lipids of fresh and cryopreserved boar sperm. Cryobiology 1994, 31, 224–238. [Google Scholar] [CrossRef]
- Cerolini, S.; Maldjian, A.; Surai, P.; Noble, R. Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage1. Anim. Reprod. Sci. 2000, 58, 99–111. [Google Scholar] [CrossRef]
- Brouwers, J.F.; Silva, P.F.; Gadella, B.M. New assays for detection and localization of endogenous lipid peroxidation products in living boar sperm after BTS dilution or after freeze-thawing. Theriogenology 2005, 63, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Di Santo, M.; Tarozzi, N.; Nadalini, M.; Borini, A. Human sperm cryopreservation: Update on techniques, effect on DNA integrity, and implications for ART. Adv. Urol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Flores, E.; Fernández-Novell, J.; Peña, A.; Rigau, T.; Rodríguez-Gil, J. Cryopreservation-induced alterations in boar spermatozoa mitochondrial function are related to changes in the expression and location of midpiece mitofusin-2 and actin network. Theriogenology 2010, 74, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Ferrusola, C.O.; Fernández, L.G.; Morrell, J.; Sandoval, C.S.; García, B.M.; Rodríguez-Martinez, H.; Tapia, J.; Peña, F. Lipid peroxidation, assessed with BODIPY-C11, increases after cryopreservation of stallion spermatozoa, is stallion-dependent and is related to apoptotic-like changes. Reproduction 2009, 138, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Pena, F.; Plaza Davila, M.; Ball, B.; Squires, E.; Martin Munoz, P.; Ortega Ferrusola, C.; Balao da Silva, C. The impact of reproductive technologies on stallion mitochondrial function. Reprod. Domest. Anim. 2015, 50, 529–537. [Google Scholar] [CrossRef]
- Yeste, M.; Estrada, E.; Rocha, L.; Marín, H.; Rodríguez-Gil, J.; Miró, J. Cryotolerance of stallion spermatozoa is related to ROS production and mitochondrial membrane potential rather than to the integrity of sperm nucleus. Andrology 2015, 3, 395–407. [Google Scholar] [CrossRef]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of apoptosis in sperm cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef]
- Castro, L.; Hamilton, T.; Mendes, C.; Nichi, M.; Barnabe, V.; Visintin, J.; Assumpção, M. Sperm cryodamage occurs after rapid freezing phase: Flow cytometry approach and antioxidant enzymes activity at different stages of cryopreservation. J. Anim. Sci. Biotechnol. 2016, 7, 17. [Google Scholar] [CrossRef]
- Sieme, H.; Oldenhof, H.; Wolkers, W.F. Mode of action of cryoprotectants for sperm preservation. Anim. Reprod. Sci. 2016, 169, 2–5. [Google Scholar] [CrossRef]
- Yang, C.-H.; Wu, T.-W.; Cheng, F.-P.; Wang, J.-H.; Wu, J.-T. Effects of different cryoprotectants and freezing methods on post-thaw boar semen quality. Reprod. Biol. 2016, 16, 41–46. [Google Scholar] [CrossRef]
- Zhang, W.; Yi, K.; Chen, C.; Hou, X.; Zhou, X. Application of antioxidants and centrifugation for cryopreservation of boar spermatozoa. Anim. Reprod. Sci. 2012, 132, 123–128. [Google Scholar] [CrossRef] [PubMed]
- De Vantéry Arrighi, C.; Lucas, H.; Chardonnens, D.; De Agostini, A. Removal of spermatozoa with externalized phosphatidylserine from sperm preparation in human assisted medical procreation: Effects on viability, motility and mitochondrial membrane potential. Reprod. Biol. Endocrinol. 2009, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Valcarce, D.; Cartón-García, F.; Herráez, M.; Robles, V. Effect of cryopreservation on human sperm messenger RNAs crucial for fertilization and early embryo development. Cryobiology 2013, 67, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Card, C.J.; Anderson, E.J.; Zamberlan, S.; Krieger, K.E.; Kaproth, M.; Sartini, B.L. Cryopreserved bovine spermatozoal transcript profile as revealed by high-throughput ribonucleic acid sequencing. Biol. Reprod. 2013, 88, 49. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.Y.; Mulligan, B.P.; Kim, H.-M.; Yang, B.-C.; Lee, C.-K. Quantitative analysis of sperm mRNA in the pig: Relationship with early embryo development and capacitation. Reprod. Fertil. Dev. 2013, 25, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Peng, W.; Ding, L.; He, L.; Zhang, Y.; Fang, D.; Tang, K. A preliminary study on epigenetic changes during boar spermatozoa cryopreservation. Cryobiology 2014, 69, 119–127. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.I.; Miyazono, K. Emerging complexity of microRNA generation cascades. J. Biochem. 2010, 149, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curry, E.; Safranski, T.J.; Pratt, S.L. Differential expression of porcine sperm microRNAs and their association with sperm morphology and motility. Theriogenology 2011, 76, 1532–1539. [Google Scholar] [CrossRef]
- Capra, E.; Turri, F.; Lazzari, B.; Cremonesi, P.; Gliozzi, T.; Fojadelli, I.; Stella, A.; Pizzi, F. Small RNA sequencing of cryopreserved semen from single bull revealed altered miRNAs and piRNAs expression between High-and Low-motile sperm populations. BMC Genom. 2017, 18, 14. [Google Scholar] [CrossRef] [PubMed]
- Ran, M.-X.; Li, Y.; Zhang, Y.; Liang, K.; Ren, Y.-N.; Zhang, M.; Zhou, G.-B.; Zhou, Y.-M.; Wu, K.; Wang, C.-D. Transcriptome Sequencing Reveals the Differentially Expressed lncRNAs and mRNAs Involved in Cryoinjuries in Frozen-Thawed Giant Panda (Ailuropoda melanoleuca) Sperm. Int. J. Mol. Sci. 2018, 19, 3066. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, R.H.; Ran, M.X.; Zhang, Y.; Liang, K.; Ren, Y.N.; He, W.C.; Zhang, M.; Zhou, G.B.; Qazi, I.H.; et al. High throughput small RNA and transcriptome sequencing reveal capacitation-related microRNAs and mRNA in boar sperm. BMC Genom. 2018, 19, 736. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gordon, E.; Harkiss, D.; Twigg, J.P.; Milne, P.; Jennings, Z.; Irvine, D.S. Relative impact of oxidative stress on the functional competence and genomic integrity of human spermatozoa. Biol. Reprod. 1998, 59, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Cordova, A.; Perez, J.; Lleo, B.; Artiga, C.G.; Rillo, S.M. In vitro fertilizing capacity of deep frozen boar semen packaged in 0.5 and 5 ml straws. Reprod. Domest. Anim. 2001, 36, 199–202. [Google Scholar] [CrossRef]
- Fraser, L. Markers for Sperm Freezability and Relevance of Transcriptome Studies in Semen Cryopreservation: A Review. In Theriogenology; Carreira, R.P., Ed.; IN TECH: Rijeka, Croatia, 2017; Chapter 3; pp. 47–62. [Google Scholar]
- Flores, E.; Cifuentes, D.; Fernandez-Novell, J.M.; Medrano, A.; Bonet, S.; Briz, M.D.; Pinart, E.; Pena, A.; Rigau, T.; Rodriguez-Gil, J.E. Freeze-thawing induces alterations in the protamine-1/DNA overall structure in boar sperm. Theriogenology 2008, 69, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Flores, E.; Ramió-Lluch, L.; Bucci, D.; Fernández-Novell, J.; Peña, A.; Rodríguez-Gil, J. Freezing-thawing induces alterations in histone H1-DNA binding and the breaking of protein-DNA disulfide bonds in boar sperm. Theriogenology 2011, 76, 1450–1464. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Wang, H.Y.; Greenawalt, D.M.; Azaro, M.A.; Luo, M.; Tereshchenko, I.V.; Cui, X.; Yang, Q.; Gao, R.; Shen, L. AccuTyping: New algorithms for automated analysis of data from high-throughput genotyping with oligonucleotide microarrays. Nucleic Acids Res. 2006, 34, e116. [Google Scholar] [CrossRef]
- Curry, E.; Ellis, S.; Pratt, S. Detection of porcine sperm microRNAs using a heterologous microRNA microarray and reverse transcriptase polymerase chain reaction. Mol. Reprod. Dev. 2009, 76, 218–219. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zeng, C.-J.; He, L.; Ding, L.; Tang, K.-Y.; Peng, W.-P. Selection of endogenous reference microRNA genes for quantitative reverse transcription polymerase chain reaction studies of boar spermatozoa cryopreservation. Theriogenology 2015, 83, 634–641. [Google Scholar] [CrossRef]
- Govindaraju, A.; Uzun, A.; Robertson, L.; Atli, M.O.; Kaya, A.; Topper, E.; Crate, E.A.; Padbury, J.; Perkins, A.; Memili, E. Dynamics of microRNAs in bull spermatozoa. Reprod. Biol. Endocrinol. 2012, 10, 82. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, D.; Chang, Y.; Li, Y.; Zhang, M.; Zhou, G.; Peng, Z.; Zeng, C. Cryopreservation of boar sperm induces differential microRNAs expression. Cryobiology 2017, 76, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Huse, J.T.; Brennan, C.; Hambardzumyan, D.; Wee, B.; Pena, J.; Rouhanifard, S.H.; Sohn-Lee, C.; Le Sage, C.; Agami, R.; Tuschl, T. The PTEN-regulating microRNA miR-26a is amplified in high-grade glioma and facilitates gliomagenesis in vivo. Genes Dev. 2009, 23, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoshan, E.; Mobley, A.K.; Braeuer, R.R.; Kamiya, T.; Huang, L.; Vasquez, M.E.; Salameh, A.; Lee, H.J.; Kim, S.J.; Ivan, C. Reduced adenosine-to-inosine miR-455-5p editing promotes melanoma growth and metastasis. Nat. Cell Biol. 2015, 17, 311. [Google Scholar] [CrossRef] [PubMed]
- Aquila, S.; Middea, E.; Catalano, S.; Marsico, S.; Lanzino, M.; Casaburi, I.; Barone, I.; Bruno, R.; Zupo, S.; Ando, S. Human sperm express a functional androgen receptor: Effects on PI3K/AKT pathway. Hum. Reprod. 2007, 22, 2594–2605. [Google Scholar] [CrossRef] [PubMed]
- Kurmasheva, R.T.; Harwood, F.C.; Houghton, P.J. Differential regulation of vascular endothelial growth factor by Akt and mammalian target of rapamycin inhibitors in cell lines derived from childhood solid tumors. Mol. Cancer Therap. 2007, 6, 1620–1628. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Nakada, M.T.; Rafferty, P.; Laraio, J.; McCabe, F.L.; Millar, H.; Cunningham, M.; Snyder, L.A.; Bugelski, P.; Yan, L. Regulation of vascular endothelial growth factor expression by EMMPRIN via the PI3K-Akt signaling pathway. Mol. Cancer Res. 2006, 4, 371–377. [Google Scholar] [CrossRef]
- Iyibozkurt, A.C.; Balcik, P.; Bulgurcuoglu, S.; Arslan, B.K.; Attar, R.; Attar, E. Effect of vascular endothelial growth factor on sperm motility and survival. Reprod. Biomed. Online 2009, 19, 784–788. [Google Scholar] [CrossRef]
- Lee, T.-C.; Ho, H.-C. Effects of prostaglandin E2 and vascular endothelial growth factor on sperm might lead to endometriosis-associated infertility. Fertil. Steril. 2011, 95, 360–362. [Google Scholar] [CrossRef]
- Sargent, K.M.; Clopton, D.T.; Lu, N.; Pohlmeier, W.E.; Cupp, A.S. VEGFA splicing: Divergent isoforms regulate spermatogonial stem cell maintenance. Cell Tissue Res. 2016, 363, 31–45. [Google Scholar] [CrossRef]
- Min, Y.H.; Cheong, J.-W.; Kim, J.Y.; Eom, J.I.; Lee, S.T.; Hahn, J.S.; Ko, Y.W.; Lee, M.H. Cytoplasmic mislocalization of p27Kip1 protein is associated with constitutive phosphorylation of Akt or protein kinase B and poor prognosis in acute myelogenous leukemia. Cancer Res. 2004, 64, 5225–5231. [Google Scholar] [CrossRef]
- Prasad, S.B.; Yadav, S.S.; Das, M.; Modi, A.; Kumari, S.; Pandey, L.K.; Singh, S.; Pradhan, S.; Narayan, G. PI3K/AKT pathway-mediated regulation of p27 Kip1 is associated with cell cycle arrest and apoptosis in cervical cancer. Cell. Oncol. 2015, 38, 215–225. [Google Scholar] [CrossRef]
- Krakowiak, P.A.; Wassif, C.A.; Kratz, L.; Cozma, D.; Kovářová, M.; Harris, G.; Grinberg, A.; Yang, Y.; Hunter, A.G.; Tsokos, M. Lathosterolosis: An inborn error of human and murine cholesterol synthesis due to lathosterol 5-desaturase deficiency. Hum. Mol. Genet. 2003, 12, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Lang-Ouellette, D.; Richard, T.; Morin, P. Mammalian hibernation and regulation of lipid metabolism: A focus on non-coding RNAs. Biochemistry 2014, 79, 1161–1171. [Google Scholar] [CrossRef]
- Schmidt, S.; Corydon, T.J.; Pedersen, C.B.; Bross, P.; Gregersen, N. Misfolding of short-chain acyl-CoA dehydrogenase leads to mitochondrial fission and oxidative stress. Mol. Genet. Metabol. 2010, 100, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, L.; Schmidt, R.E.; Su, C.; Huang, X.; Gould, K.; Cao, G. Characterization of HSCD5, a novel human stearoyl-CoA desaturase unique to primates. Biochem. Biophys. Res. Commun. 2005, 332, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yanzhu, Y.; Yuguang, S. Characterization of human SCD2, an oligomeric desaturase with improved stability and enzyme activity by cross-linking in intact cells. Biochem. J. 2005, 388, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.Q.; Ruden, D.M.; Lu, X.Y. PKD2 cation channel is required for directional sperm movement and male fertility. Curr. Biol. 2003, 13, 2175–2178. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.C.; Phadnis, N. Parallel evolution of sperm hyper-activation ca2+ channels. Genome Bio. Evol. 2017, 9, 1938–1949. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lu, X.Y. Drosophila sperm motility in the reproductive tract. Bio. Reprod. 2011, 84, 1005–1015. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Zhu, H.; Hao, H.; Zhao, X.; Qin, T.; Wang, D. Comparative transcript profiling of gene expression of fresh and frozen-thawed bull sperm. Theriogenology. 2015, 4, 504–511. [Google Scholar] [CrossRef]
- Card, C.J.; Kreiger, K.E.; Kaproth, M.; Sartini, B.L. Oligo-dT selected spermatozoal transcript profiles differ among higher and lower fertility dairy sires. Anim. Reprod. Sci. 2017, 177, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Chen, X.; Wang, Z.; Wang, D. Is transcription in sperm stationary or dynamic? J. Reprod. Dev. 2017, 63, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Dzeja, C.; Hagen, V.; Kaupp, U.B.; Frings, S. Ca2+ permeation in cyclic nucleotide-gated channels. EMBO J. 1999, 18, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhao, H.; Xu, B.; Jiang, Y. Odorant receptor might be related to sperm DNA integrity in Apis cerana cerana. Anim. Reprod. Sci. 2018, 193, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Moreno, J.; Esteso, M.C.; Pradiee, J.; Castano, C.; Toledano-Diaz, A.; O’Brien, E.; Lopez-Sebastian, A.; Martinez-Nevado, E.; Delclaux, M.; Fernandez-Moran, J.; et al. Giant panda (Ailuropoda melanoleuca) sperm morphometry and function after repeated freezing and thawing. Andrologia 2016, 48, 470–474. [Google Scholar] [CrossRef] [PubMed]
- King, G.J.; Macpherson, J.W. A comparison of two methods for boar semen collection. J. Anim. Sci. 1973, 36, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; He, L.; Peng, W.; Ding, L.; Tang, K.; Fang, D.; Zhang, Y. Selection of optimal reference genes for quantitative RT-PCR studies of boar spermatozoa cryopreservation. Cryobiology 2014, 68, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Raw Reads | Clean Reads | Q30 (%) | Mapped Reads | Total miRNAs | Known miRNAs | Novel miRNAs |
|---|---|---|---|---|---|---|---|
| Fresh sperm [42] | 18,956,444 | 12,561,033 | 86 | 3,027,230 | 1028 | 259 | 769 |
| Frozen-thawed sperm | 16,507,275 | 11,100,601 | 85 | 2,377,337 | 984 | 246 | 738 |
| Group | Clean Reads (Pair-End) | Clean Bases | GC Content | Q30 (%) | Mapped Reads (Single-End) | Unique Mapped Reads |
|---|---|---|---|---|---|---|
| Fresh sperm [42] | 26,843,452 | 6,642,110,360 | 48.62% | 86.90 | 30,016,749 (55.91%) | 28,565,403 (53.21%) |
| Frozen-thawed sperm | 24,611,191 | 6,084,424,468 | 45.49% | 86.05 | 24,785,693 (50.35%) | 23,864,342 (48.48%) |
| Gene Name | Primer Sequence | Amplicon (bp) | GenBank/miRBase Accession Number |
|---|---|---|---|
| PPIA | F: CACAAACGGTTCCCAGTTTT | 171 | NM_214353 |
| R: TGTCCACAGTCAGCAATGGT | |||
| ACADS | F: CCAGGGCATCCAGTTCAAGT | 102 | NM_213898 |
| R: TTGCCGGCTCCTTGATGAAT | |||
| CDKN1B | F: TGGAGGGCAAATACGAGTGG | 150 | NM_214316 |
| R: CAATTAAAGGCACCGCCTGG | |||
| ORAI1 | F: TGCATCTGTTTGCGCTGATG | 168 | NM_001173519 |
| R: CCAGGAAGAGCAGTGTACCG | |||
| PRKD2 | F: GGAAAACGTGTTGTTGGCGT | 157 | XM_021094608 |
| R: GTTGTAGCCCTGGTTGAGCA | |||
| PTPN1 | F: TACACCGTCCGACAGCTAGA | 149 | DQ239903 |
| R: CCCGACTCACGGACTTTGAA | |||
| SC5D | F: CGGCTGGTTTCGACTCCTT | 175 | AY609684.1 |
| R: AGCCATCCAGAGGGTGAAAAG | |||
| U6 | F: TTATGGGTCCTAGCCTGAC | - | EU520423 |
| R: CACTATTGCGGGTCTGC | |||
| ssc-miR-212 | ACCTTGGCTCTAGACTGCTTACT | - | MI0022140 |
| ssc-miR-186 | CAAAGAATTCTCCTTTTGGGCTT | - | MI0002456 |
| ssc-miR-128 | TCACAGTGAACCGGTCTCTTT | - | MIMAT0002157 |
| ssc-miR-126 | TCGTACCGTGAGTAATAATGCG | - | MI0016619 |
| unconservative-X-272462 | TGAACGGTGCCTGTGTGGCTAGA | - | / |
| conservative-15-100875 | TCTCTGCTGCGCTCTTTCCTGA | - | / |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, D.-H.; Qazi, I.H.; Ran, M.-X.; Liang, K.; Zhang, Y.; Zhang, M.; Zhou, G.-B.; Angel, C.; Zeng, C.-J. Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing. Int. J. Mol. Sci. 2019, 20, 802. https://doi.org/10.3390/ijms20040802
Dai D-H, Qazi IH, Ran M-X, Liang K, Zhang Y, Zhang M, Zhou G-B, Angel C, Zeng C-J. Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing. International Journal of Molecular Sciences. 2019; 20(4):802. https://doi.org/10.3390/ijms20040802
Chicago/Turabian StyleDai, Ding-Hui, Izhar Hyder Qazi, Ming-Xia Ran, Kai Liang, Yan Zhang, Ming Zhang, Guang-Bin Zhou, Christiana Angel, and Chang-Jun Zeng. 2019. "Exploration of miRNA and mRNA Profiles in Fresh and Frozen-Thawed Boar Sperm by Transcriptome and Small RNA Sequencing" International Journal of Molecular Sciences 20, no. 4: 802. https://doi.org/10.3390/ijms20040802