Cloning and Partial Characterization of an Endo-α-(1→6)-d-Mannanase Gene from Bacillus circulans

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of the Full-Size Endo-α-(1→6)-d-Mannanase Gene from Bacillus circulans TN-31
2.2. Expression and Purification of the Glycosyl Hydrolase Domain of Emn
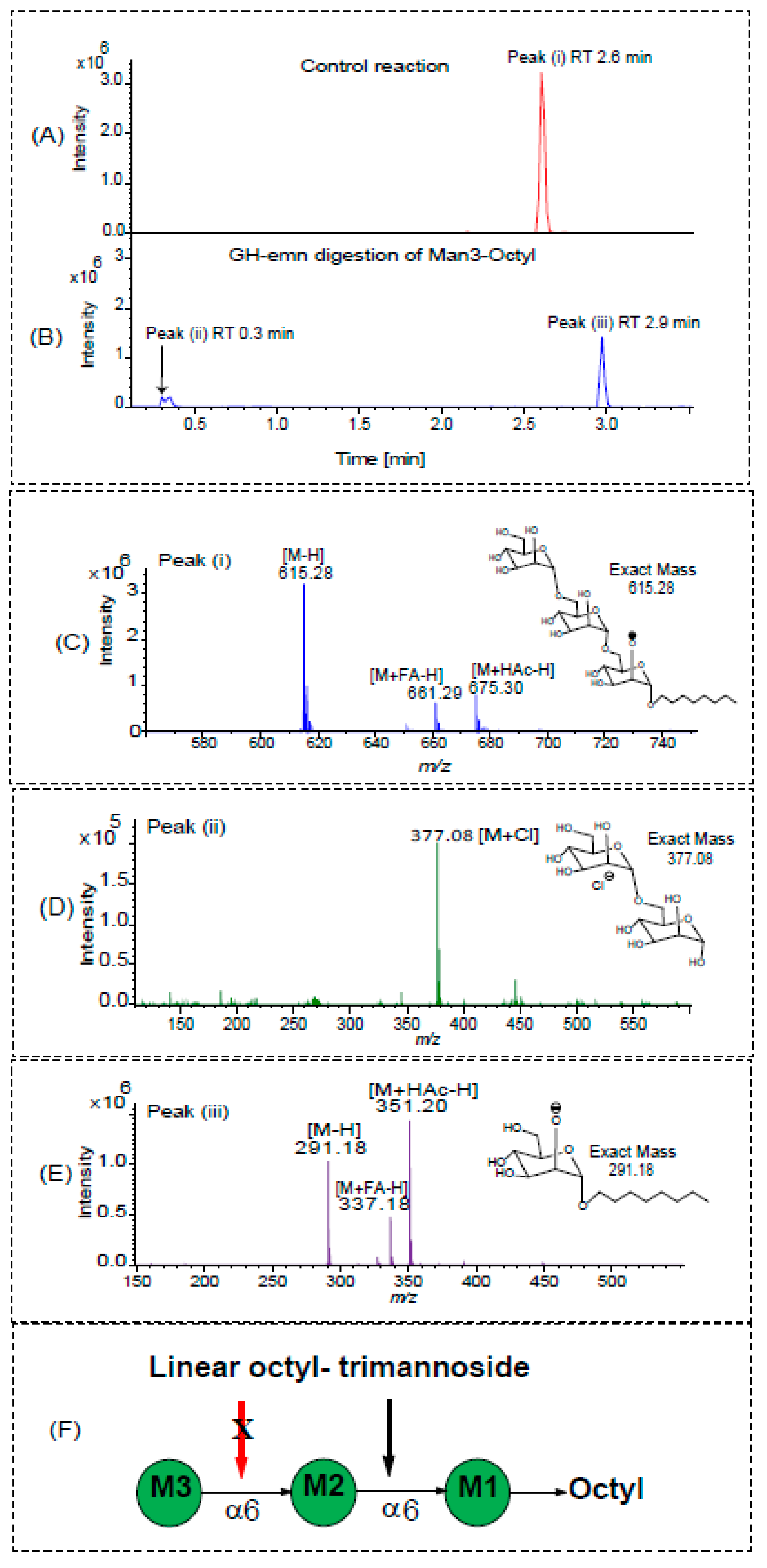
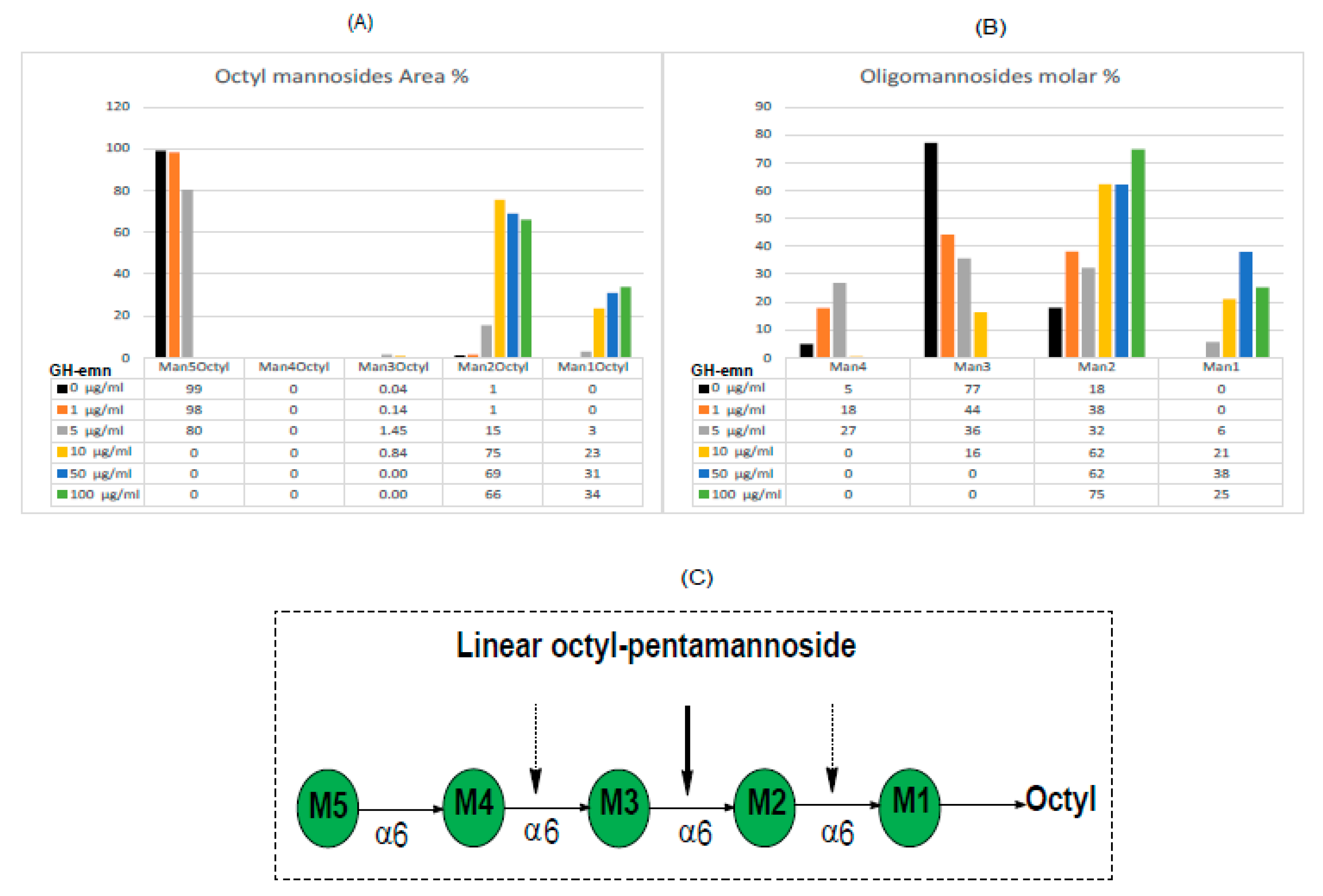
2.3. Digestion of Synthetic Mannoside Substrates
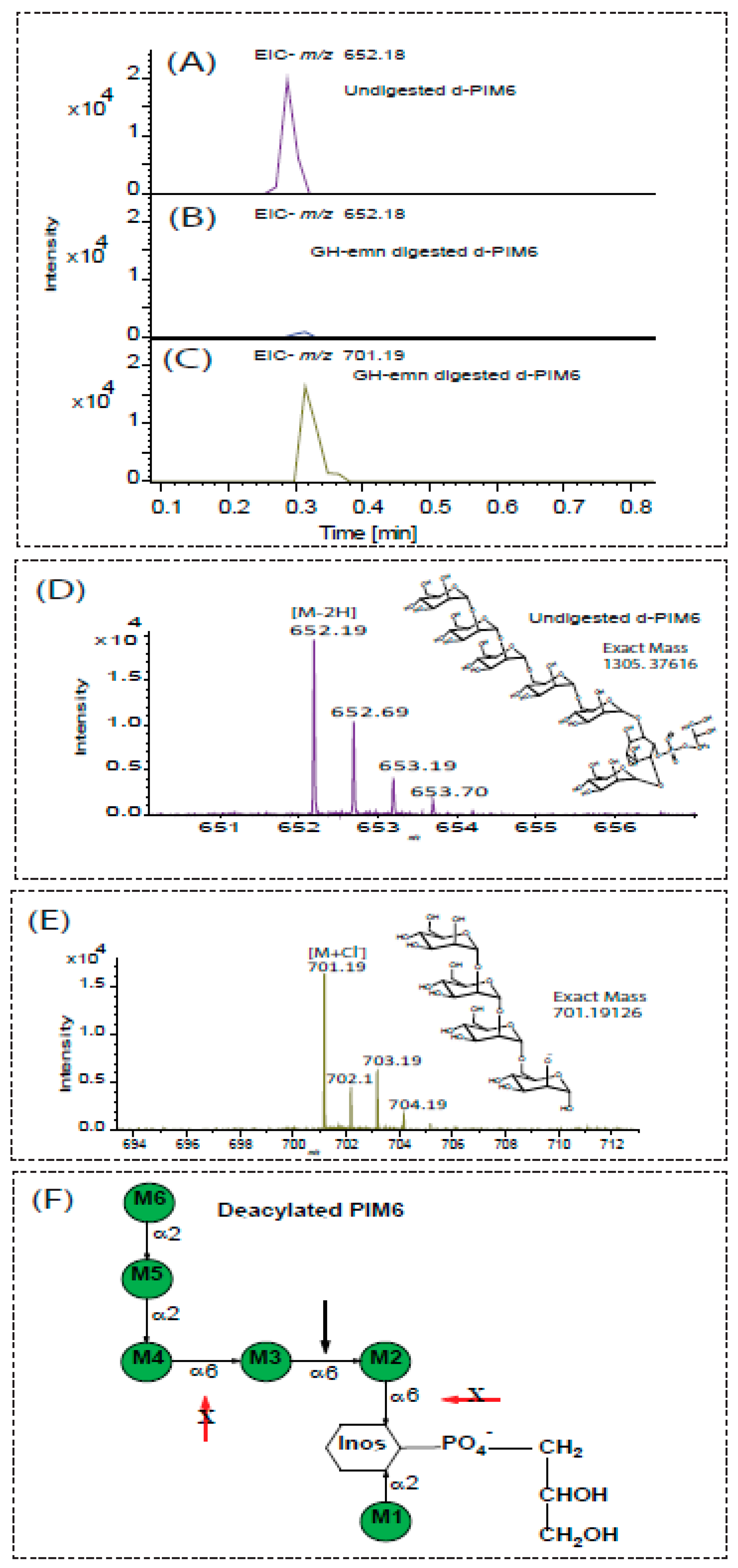
2.4. Digestion of Phosphatidyl-Myo-Inositol-Mannosides
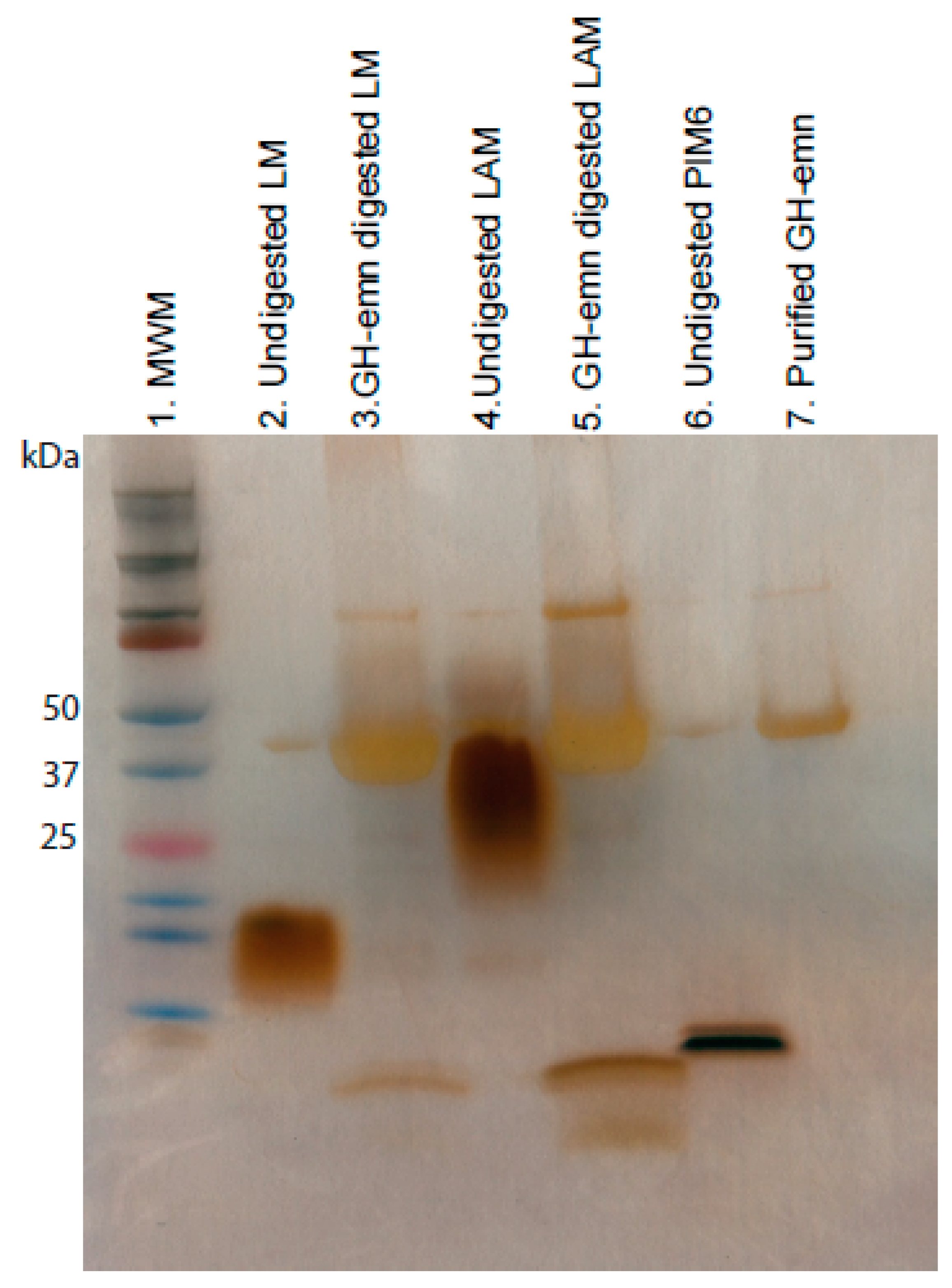
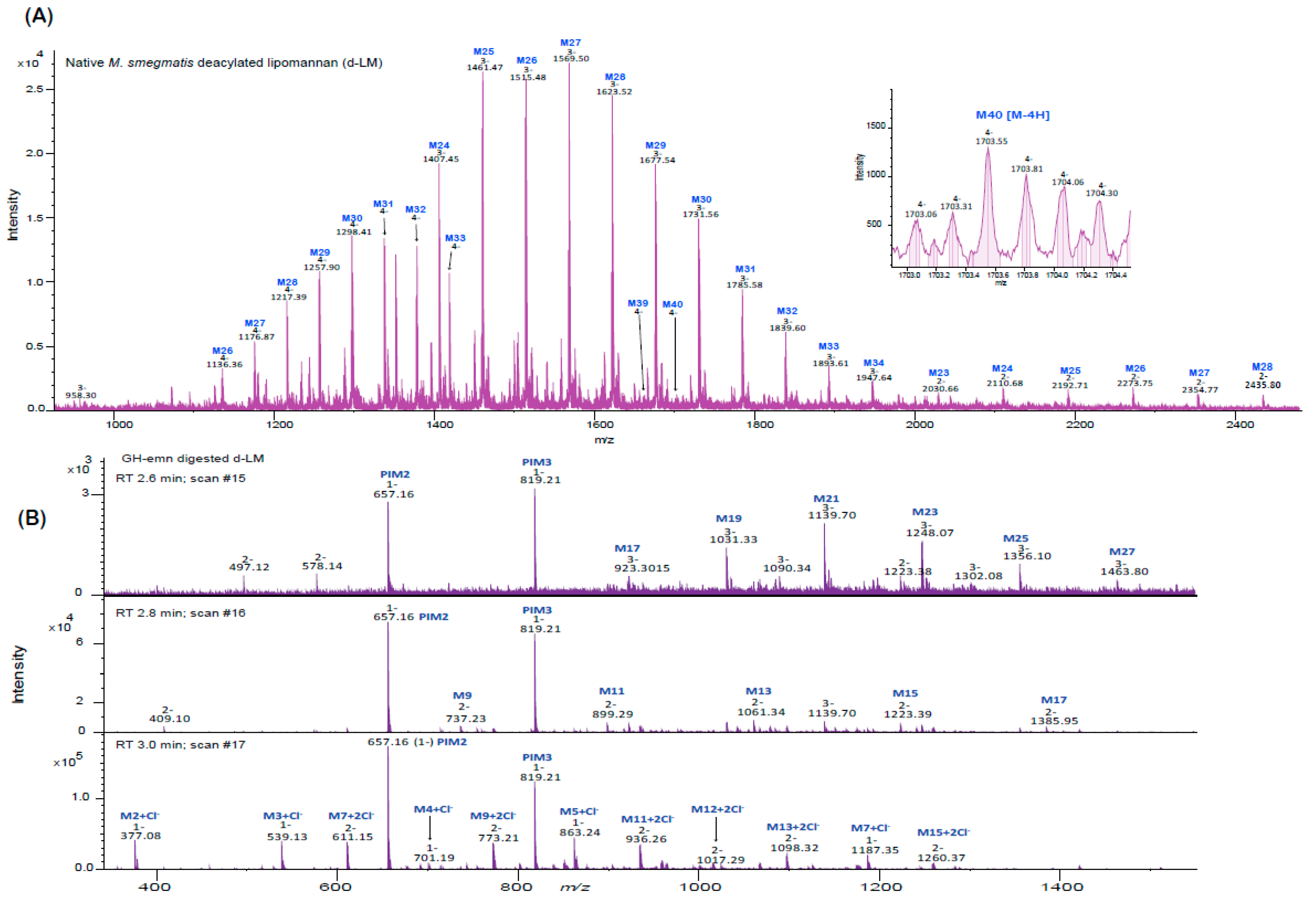
2.5. Analysis of Endomannanase-Digested LAM and LM
3. Materials and Methods
3.1. Cloning, Expression and Purification the Glycosyl Hydrolase Domain of Emn
3.2. Assays Using Synthetic Mannosides Substrates
3.3. Assays Using Purified Mycobacterial PIMs and Lipoglycans
3.4. Analysis of Substrates and Reaction Products by Liquid Chromatography—Mass Spectrometry
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Angala, S.K.; Belardinelli, J.M.; Huc-Claustre, E.; Wheat, W.H.; Jackson, M. The Cell Envelope Glycoconjugates of Mycobacterium Tuberculosis. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 361–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Matsumura, T.; Ato, M.; Hamasaki, M.; Nishiuchi, Y.; Murakami, Y.; Maeda, Y.; Yoshimori, T.; Matsumoto, S.; Kobayashi, K.; et al. Critical Roles for Lipomannan and Lipoarabinomannan in Cell Wall Integrity of Mycobacteria and Pathogenesis of Tuberculosis. MBio 2013, 4, e00472-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergne, I.; Gilleron, M.; Nigou, J. Manipulation of the Endocytic Pathway and Phagocyte Functions by Mycobacterium Tuberculosis Lipoarabinomannan. Front. Cell. Infect. Microbiol. 2014, 4, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, E.; Mori, D.; Yamasaki, S. Recognition of Mycobacterial Lipids by Immune Receptors. Trends Immunol. 2017, 38, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T. Presence of Alpha-D-Mannosidic Linkage in Glycoproteins. Liberation of D-Mannose from Various Glycoproteins by Alpha-Mannosidase Isolated from Jack Bean Meal. J. Biol. Chem. 1966, 241, 1010–1012. [Google Scholar] [PubMed]
- Chatterjee, D.; Hunter, S.W.; McNeil, M.; Brennan, P.J. Lipoarabinomannan. Multiglycosylated Form of the Mycobacterial Mannosylphosphatidylinositols. J. Biol. Chem. 1992, 267, 6228–6233. [Google Scholar] [PubMed]
- Nakajima, T.; Ballou, C.E. Structure of the Linkage Region between the Polysaccharide and Protein Parts of Saccharomyces Cerevisiae Mannan. J. Biol. Chem. 1974, 249, 7685–7694. [Google Scholar] [PubMed]
- Nakajima, T.; Maitra, S.K.; Ballou, C.E. An Endo-alpha1 leads to 6-D-Mannanase from a Soil Bacterium. Purification, Properties, and Mode of Action. J. Biol. Chem. 1976, 251, 174–181. [Google Scholar] [PubMed]
- Hunter, S.W.; Brennan, P.J. Evidence for the Presence of a Phosphatidylinositol Anchor on the Lipoarabinomannan and Lipomannan of Mycobacterium Tuberculosis. J. Biol. Chem. 1990, 265, 9272–9279. [Google Scholar] [PubMed]
- Maruyama, Y.; Nakajima, T. The Aman6 Gene Encoding a Yeast Mannan Backbone Degrading 1,6-Alpha-D-Mannanase in Bacillus Circulans: Cloning, Sequence Analysis, and Expression. Biosci. Biotechnol. Biochem. 2000, 64, 2018–2020. [Google Scholar] [CrossRef] [PubMed]
- Guerin, M.E.; Kordulakova, J.; Alzari, P.M.; Brennan, P.J.; Jackson, M. Molecular Basis of Phosphatidyl-Myo-Inositol Mannoside Biosynthesis and Regulation in Mycobacteria. J. Biol. Chem. 2010, 285, 33577–33583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, P.H.; Besra, G.S.; Lowary, T.L. Exploring the Substrate Specificity of a Mycobacterial Polyprenol Monophosphomannose-Dependent Alpha-(1-->6)-Mannosyltransferase. ChemBioChem 2008, 9, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Angala, S.K.; McNeil, M.R.; Zou, L.; Liav, A.; Zhang, J.; Lowary, T.L.; Jackson, M. Identification of a Novel Mycobacterial Arabinosyltransferase Activity Which Adds an Arabinosyl Residue to Alpha-D-Mannosyl Residues. ACS Chem. Biol. 2016, 11, 1518–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, D.; Obregon-Henao, A.; Pham, H.; Chatterjee, D.; Brennan, P.J.; Jackson, M. Lipoarabinomannan of Mycobacterium: Mannose Capping by a Multifunctional Terminal Mannosyltransferase. Proc. Natl. Acad. Sci. USA 2008, 105, 17973–17977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brearley, C.A.; Hanke, D.E. Assaying Inositol Phospholipid Turnover in Plant Cells. Signal. Inosit. A Pract. Approach 1997, 176, 1–32. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angala, S.k.; Li, W.; Palčeková, Z.; Zou, L.; Lowary, T.L.; McNeil, M.R.; Jackson, M. Cloning and Partial Characterization of an Endo-α-(1→6)-d-Mannanase Gene from Bacillus circulans. Int. J. Mol. Sci. 2019, 20, 6244. https://doi.org/10.3390/ijms20246244
Angala Sk, Li W, Palčeková Z, Zou L, Lowary TL, McNeil MR, Jackson M. Cloning and Partial Characterization of an Endo-α-(1→6)-d-Mannanase Gene from Bacillus circulans. International Journal of Molecular Sciences. 2019; 20(24):6244. https://doi.org/10.3390/ijms20246244
Chicago/Turabian StyleAngala, Shiva kumar, Wei Li, Zuzana Palčeková, Lu Zou, Todd L. Lowary, Michael R. McNeil, and Mary Jackson. 2019. "Cloning and Partial Characterization of an Endo-α-(1→6)-d-Mannanase Gene from Bacillus circulans" International Journal of Molecular Sciences 20, no. 24: 6244. https://doi.org/10.3390/ijms20246244