Invariant Natural Killer T-Cells and Total CD1d Restricted Cells Differentially Influence Lipid Metabolism and Atherosclerosis in Low Density Receptor Deficient Mice

 ,
,
Abstract
:1. Introduction
2. Results
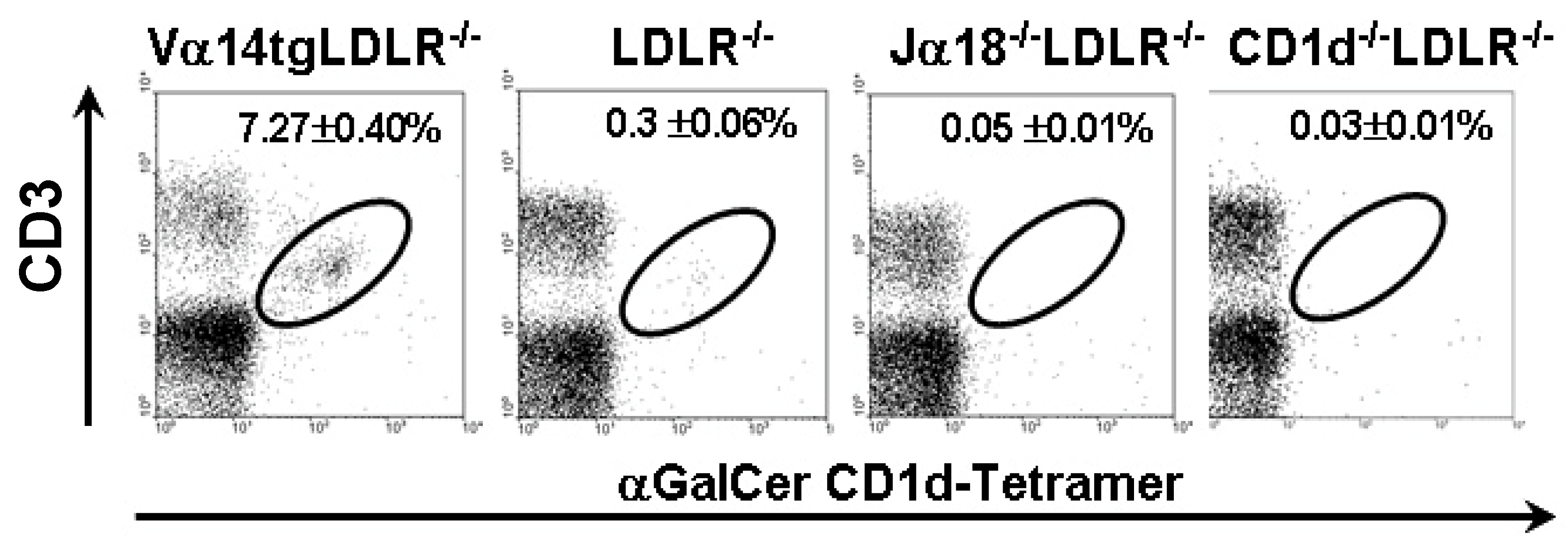
2.1. Animals with Varying Levels of iNKT Cells
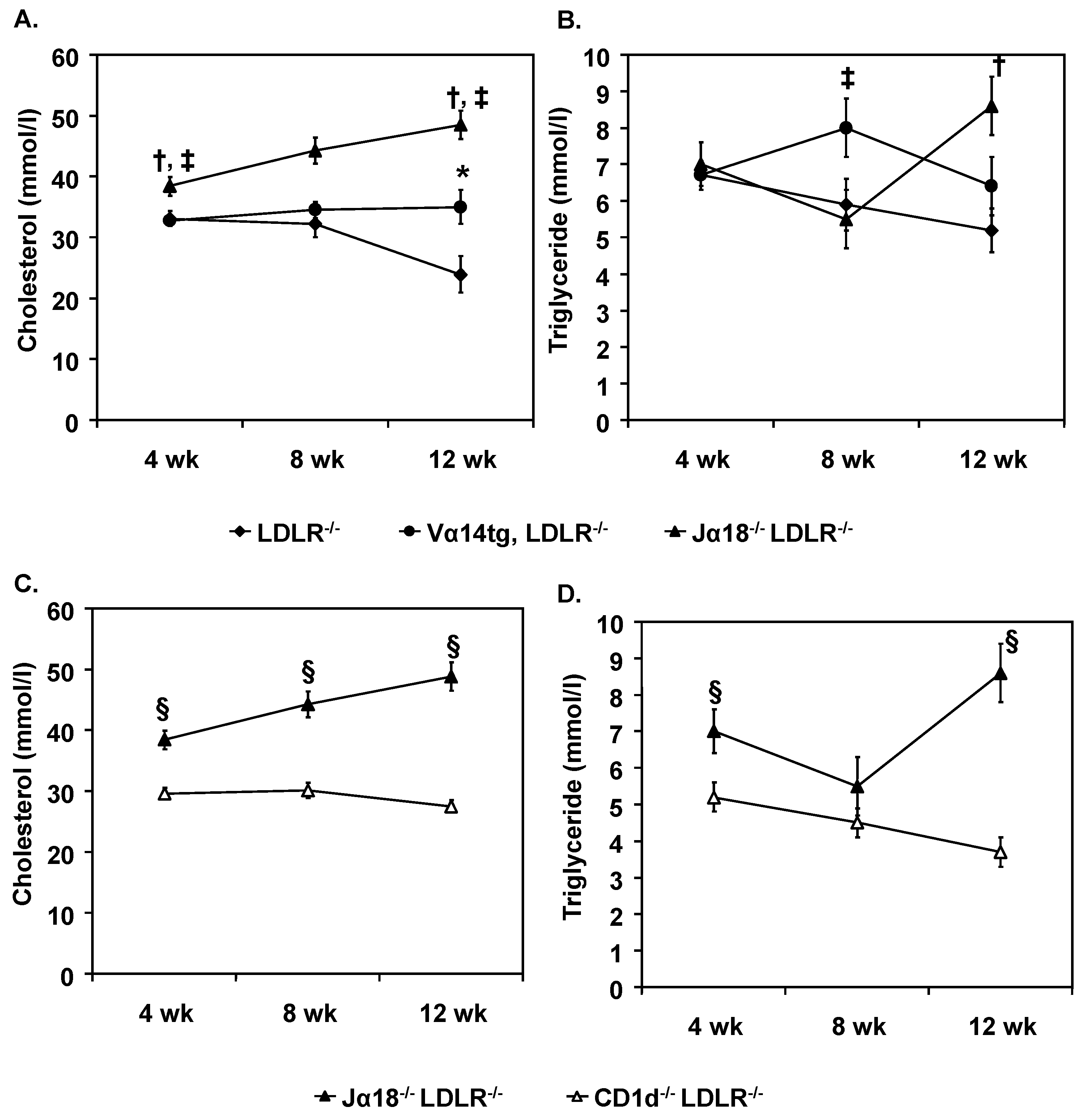
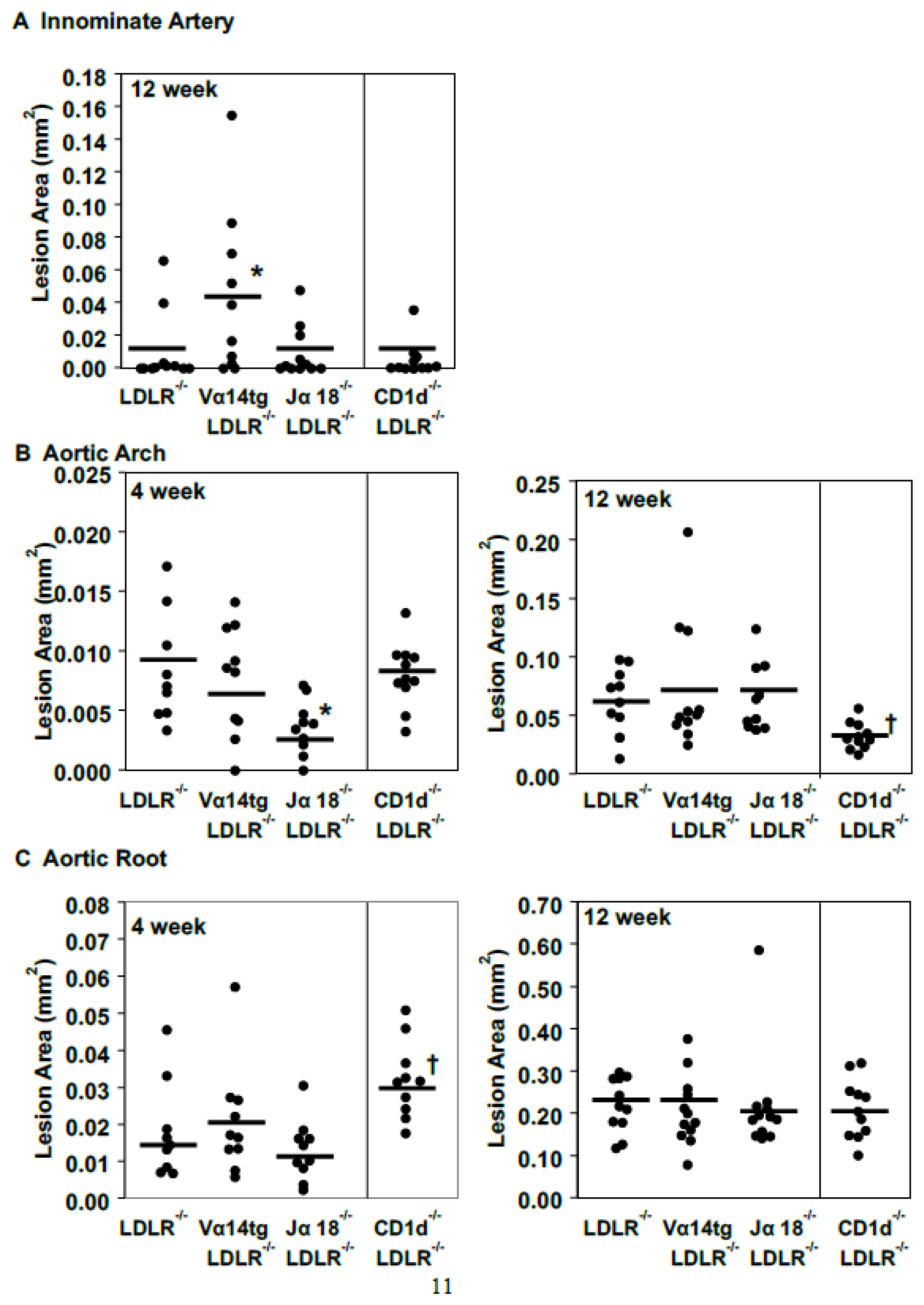
2.2. CD1d Expressing and Restricted Cells Have Separate Influences on Lipoprotein Metabolism and Atherosclerosis
2.3. Hepatic Gene Expression.
3. Discussion.
3.1. NKT Cell Status and Lipoprotein Homeostasis
3.2. NKT Cell Status and Atherosclerosis Phenotype
4. Materials and Methods
4.1. Mice
4.2. Lymphocyte Isolation
4.3. Flow Cytometery
4.4. Tissue Preparation and Histomorphology
4.5. VLDL Production Rate Experiments.
4.6. Statistical Analysis.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansson, G.K.; Robertson, A.K.L.; Söderberg-Nauclér, C. Inflammation and atherosclerosis. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 297–329. [Google Scholar] [CrossRef] [PubMed]
- Tabas, I.; Lichtman, A.H. Monocyte-macrophages and T cells in atherosclerosis. Immunity. 2017, 47, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Getz, G.S.; VanderLaan, P.A.; Reardon, C.A. The immune system and murine atherosclerosis. Curr. Drug Targets 2007, 8, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Reardon, C.A.; Blachowicz, L.; White, T.; Cabana, V.; Wang, Y.; Lukens, J.; Bluestone, J.; Getz, G.S. Effect of immune deficiency on lipoproteins and atherosclerosis in male apoprotein E deficient mice. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Dansky, H.M.; Charlton, S.A.; Harper, M.M.; Smith, J.D. T and B lymphocytes play a minor role in atherosclerotic plaque formation in the apolipoprotein E-deficient mouse. Proc. Natl. Acad. Sci. USA. 1997, 94, 4642–4646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Leung, C.; Schindler, C. Lymphocytes are important in early atherosclerosis. J. Clin. Invest. 2001, 108, 251–259. [Google Scholar] [CrossRef]
- Zhou, X.; Robertson, A.K.; Hjerpe, C.; Hansson, G.K. Adoptive transfer of CD4+ T cells reactive to modified low-density lipoprotein aggravates atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 864–870. [Google Scholar] [CrossRef]
- Laurat, E.; Poirier, B.; Tupin, E.; Caligiuri, G.; Hansson, G.K.; Bariéty, J.; Nicoletti, A. In vivo downregulation of T helper cell 1 immune responses reduces atherogenesis in apolipoprotein E-knockout mice. Circulation. 2001, 104, 197–202. [Google Scholar] [CrossRef]
- Mallat, Z.; Tedgui, A. Cytokines as regulators of atherosclerosis in mice. Curr. Drug Targets 2007, 8, 1264–1272. [Google Scholar] [CrossRef]
- Gorsman, I.; Sharpe, A.H.; Lichtman, A.H. T-cell costimulation and coinhibition in atherosclerosis. Circ. Res. 2008, 103, 1220–1231. [Google Scholar] [CrossRef]
- Ait-Oufella, H.; Salomon, B.L.; Potteaux, S.; Robertson, A.K.; Gourdy, P.; Zoll, J.; Merval, R.; Esposito, B.; Cohen, J.L.; Fisson, S.; et al. Natural regulatory T cells control the development of atherosclerosis in mice. Nat. Med. 2006, 12, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, G.; Rudling, M.; Ollivier, V.; Jacob, M.P.; Michel, J.B.; Hansson, G.K.; Nicoletti, A. Interleukin-10 deficiency increases atherosclerosis, thrombosis, and low-density lipoproteins in apolipoprotein E knockout mice. Mol. Med. 2003, 9, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Pinderski, L.J.; Fischbein, M.P.; Subbanagounder, G.; Fishbein, M.C.; Kubo, N.; Cheroutre, H.; Curtiss, L.K.; Berliner, J.A.; Boisvert, W.A. Overexpression of interleukin-10 by activated T lymphocytes inhibits atherosclerosis in LDL receptor-deficient mice by altering lymphocyte and macrophage phenotypes. Circ. Res. 2002, 90, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.K.; Rudling, M.; Zhou, X.; Gorelik, L.; Flavell, R.A.; Hansson, G.K. Disruption of TGF-beta signaling in T cells accelerates atherosclerosis. J. Clin. Invest. 2003, 112, 1342–1350. [Google Scholar] [CrossRef] [PubMed]
- Gojova, A.; Brun, V.; Esposito, B.; Cottrez, F.; Gourdy, P.; Ardouin, P.; Tedgui, A.; Mallat, Z.; Groux, H. Specific abrogation of transforming growth factor-beta signaling in T cells alters atherosclerotic lesion size and composition in mice. Blood 2003, 102, 4052–4058. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Gojova, A.; Marchiol-Fournigault, C.; Esposito, B.; Kamaté, C.; Merval, R.; Fradelizi, D.; Tedgui, A. Inhibition of transforming growth factor-beta signaling accelerates atherosclerosis and induces an unstable plaque phenotype in mice. Circ. Res. 2001, 89, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Bendelac, A.; Savage, PB.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M. Towards an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 2005, 26, 877–900. [Google Scholar] [CrossRef]
- Van Kaer, L.; Parekh, V.V.; Wu, L. In variant natural killer T cells: Bridging innate and adaptive immunity. Cell Tissue Res. 2011, 343, 43–55. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Stankovic, S.; Baxter, A.G. Raising the NKT cell family. Nat. Immunol. 2010, 11, 197–206. [Google Scholar] [CrossRef]
- Liao, D.M.; Zimmer, M.I.; Wang, C.R. The functions of type 1 and type II natural killer T cells in inflammatory bowel diseases. Inflamm. Bowel. Dis. 2013, 19, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Iwabuchi, K.; Fujii, S.; Ishimori, N.; Dashtsoodol, N.; Watano, K.; Mishima, T.; Iwabuchi, C.; Tanaka, S.; Bezbradica, J.S.; et al. Natural killer T cells accelerate atherogenesis in mice. Blood 2004, 104, 2051–2059. [Google Scholar] [CrossRef] [PubMed]
- Tupin, E.; Nicoletti, A.; Elhage, R.; Rudling, M.; Ljunggren, H.G.; Hansson, G.K.; Berne, G.P. CD1d-dependent activation of NKT cells aggravates atherosclerosis. J. Exp. Med. 2004, 199, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Major, A.S.; Wilson, M.T.; McCaleb, J.L.; Ru Su, Y.; Stanic, A.K.; Joyce, S.; Van Kaer, L.; Fazio, S.; Linton, MF. Quantitative and qualitative differences in proatherogenic NKT cells in apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 2351–2357. [Google Scholar] [CrossRef] [PubMed]
- Aslanian, A.M.; Chapman, H.A.; Charo, I.F. Transient role for CD1d-restricted natural killer T cells in the formation of atherosclerotic lesions. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 628–632. [Google Scholar] [CrossRef]
- Rogers, L.; Burchat, S.; Gage, J.; Hasu, M.; Thabet, M.; Willcox, L.; Ramsamy, T.A.; Whitman, S.C. Deficiency of invariant Vα14 natural killer T cells decreases atherosclerosis in LDL receptor null mice. Cardiovasc. Res. 2008, 78, 167–174. [Google Scholar] [CrossRef]
- van Puijvelede, G.H.M.; van Wanrooij, A.J.A.; Hauer, A.D.; de Vos, P.; van Berkel, T.J.C.; Kuiper, J. Effect of natural killer T cell activation on initiation of atherosclerosis. Thromb. Haemost. 2009, 102, 223–230. [Google Scholar] [CrossRef] [Green Version]
- VanderLaan, P.A.; Reardon, C.A.; Sagiv, Y.; Blachowicz, L.; Lukens, J.; Nissenbaum, M.; Wang, C.R.; Getz, G.S. Characterization of the natural killer T-cell response in an adoptive transfer model of atherosclerosis. Am. J. Pathol. 2007, 170, 1100–1107. [Google Scholar] [CrossRef]
- Li, Y.; To, K.; Kanellakis, P.; Hosseini, H.; Deswaerte, V.; Tipping, P.; Smyth, M.J.; Toh, B.H.; Bobik, A.; Kyaw, T. CD4+ natural killer T cells potently augment aortic root atherosclerosis by perforin- and granzyme B-dependent cytotoxicity. Circ. Res. 2015, 116, 245–254. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Natural killer T cells in atherosclerosis. Nat. Rev. Cardiol. 2017, 14, 304–314. [Google Scholar] [CrossRef]
- Braun, N.A.; Mendez-Fernandez, Y.V.; Covarrubias, R.; Porcelli, S.A.; Savage, P.B.; Yagita, H.; Van Kaer, L.; Major, A.S. Development of spontaneous anergy in invariant natural killer T cells in a mouse model of dyslipidemia. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1758–1765. [Google Scholar] [CrossRef] [PubMed]
- Crispe, I.N. The liver as a lymphoid organ. Ann. Rev. Immunol. 2009, 27, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Dougan, S.K.; Kaser, A.; Blumberg, R.S. CD1 expression on antigen-presenting cells. Curr. Top. Microbiol. Immunol. 2007, 314, 113–141. [Google Scholar] [PubMed]
- Subramanian, S.; Goodspeed, L.; Wang, S.; Ding, Y.; O’Brien, K.D.; Getz, G.S.; Chait, A.; Reardon, C.A. Deficiency of invariant natural killer T cells does not protect against obesity but exacerbates atherosclerosis in Ldlr−/− mice. Int. J. Mol. Sci. 2018, 19, 510. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Shin, T.; Kawano, T.; Sato, H.; Kondo, E.; Toura, I.; Kaneko, Y.; Koseki, H.; Kanno, M.; Taniguchi, M. Requirement for Vα14 NKT cells in IL-12-mediated rejection of tumors. Science 1997, 278, 1623–1626. [Google Scholar] [CrossRef]
- Bedel, R.; Matsuda, JL.; Brigl, M.; White, J.; Kappler, J.; Marrack, P.; Laurent, G. Lower TCR repertoire diversity in Traj18-deficient mice. Nat. Immunol. 2012, 13, 705–706. [Google Scholar] [CrossRef] [PubMed]
- Rouxel, O.; Lehuen, A. Mucosal-associated invariant T cells in autoimmunity and immune-mediated diseases. Immunol. Cell Biol. 2018, 96, 618–619. [Google Scholar] [CrossRef]
- Pasquet, L.; Kato, S.; Adams, T.; Sharrow, S.O.; Davies-Hill, T.; Jaffe, E.S.; Xia, Z.; Suzuki, M.; Kovalovski, D.; Berzofsky, J.A.; et al. Functionality of lung type II NKT cells developing in CD1d−/− animals. J. Immunol. 2017, 198. Available online: https://www.jimmunol.org/content/198/1_Supplement/208.18 (accessed on 30 July 2019).
- Berzofsky, J.A.; Terabe, M. The contrasting roles of NKT cells in tumor immunity. Curr. Mol. Med. 2009, 9, 667–672. [Google Scholar] [CrossRef]
- Arrenberg, P.; Halder, R.; Kumar, V. Cross-regulation between distinct natural killer T cell subsets influences immune response to self and foreign antigens. J. Cell. Physiol. 2009, 218, 246–250. [Google Scholar] [CrossRef]
- Lewis, K.E.; Kirk, E.A.; McDonald, T.O.; Wang, S.; Wight, T.N.; O’Brien, K.D.; Chait, A. Increase in serum amyloid A evoked by dietary cholesterol is associated with increased atherosclerosis in mice. Circulation 2004, 110, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Krijgsman, D.; Hokland, M.; Kuppen, P.J.K. The role of natural killer T cells in cancer- A phenotypical and functional approach. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- De Santo, C.; Arscott, R.; Booth, S.; Karydis, I.; Jones, M.; Asher, R.; Salio, M.; Middleton, M.; Cerundolo, V. Invariant NKT cells modulate the suppressive activity of IL-10-secreting neutrophils differentiated with serum amyloid A. Nat. Immunol. 2010, 11, 1039–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Elzen, P.; Garg, S.; León, L.; Brigl, M.; Leadbetter, E.A.; Gumperz, J.E.; Dascher, C.C.; Cheng, T.Y.; Sacks, F.M.; Illarionov, P.A.; et al. Apolipoprotein-mediated pathways of lipid antigen presentation. Nature 2009, 437, 906–910. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, M.I.; Nguyen, H.P.; Wang, B.; Xu, H.; Colmone, A.; Felio, K.; Choi, H.J.; Zhou, P.; Alegre, M.L.; Wang, C.R. Polymorphisms in CD1d affect antigen presentation and the activation of CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 1909–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundararaj, S.; Zhang, J.; Krovi, S.H.; Bedel, R.; Tuttle, K.D.; Veerapen, N.; Besra, G.S.; Khandokar, Y.; Praveena, T.; Le Nours, J.; et al. Differing roles of Cd1d2 and CD1d1 proteins in type I natural killer T cells development and function. Proc. Natl. Acad. Sci. USA. 2008, 115, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Mendiratta, S.K.; Martin, W.D.; Hong, S.; Boesteanu, A.; Joyce, S.; Van Kaer, L. CD1d1 mutant mice are deficient in natural T cells that promptly produced IL-4. Immunity 1997, 6, 469–477. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chiu, N.M.; Manual, M.; Wang, N.; Wang, C.R. Impaired NK1+ T cell development and early IL-4 production in CD1- deficient mice. Immunity 1997, 6, 459–467. [Google Scholar] [CrossRef]
- Barral, D.C.; Brenner, M.B. CD1 antigen presentation: How it works. Nat. Rev. Immunol. 2007, 7, 929–941. [Google Scholar] [CrossRef]
- Wingender, G.; Kronenberg, M. Role of NKT cells in the digestive system: IV. The role of canonical natural killer T cells in mucosal immunity and infection. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1–G8. [Google Scholar] [CrossRef]
- Bouaziz, J.D.; Yanaba, K.; Tedder, F.R. Regulatory B cells as inhibitors of immune responses and inflammation. Immunol. Rev. 2008, 224, 201–244. [Google Scholar] [CrossRef] [PubMed]
- Kritikou, E.; van Duijn, J.; Nahon, J.E.; van der Heijden, T.; Bouwman, M.; Groeneveldt, C.; Schaftenaar, F.H.; Kröner, M.J.; Kuiper, J.; van Puijvelde, G.H.M.; et al. Disruption of a CD1d-mediated interaction between mast cells and NKT cells aggravates atherosclerosis. Atherosclerosis 2019, 280, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Zeissig, S.; Blumberg, R.S. Commensal microbiota and NKT cells in the control of inflammatory diseases at mucosal surfaces. Curr. Opin. Immunol. 2013, 25, 690–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuis, E.E.; Matumoto, T.; Lindenbergh, D.; Willemsen, R.; Kaser, A.; Simons-Oosterhuis, Y.; Brugman, S.; Yamaguchi, K.; Ishikawa, H.; Aiba, Y.; et al. CD1d-dependent regulation of bacterial colonization in the intestine of mice. J. Clin. Invest. 2009, 119, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Bendelac, A.; Hunziker, R.D.; Lantz, O. Increased interleukin 4 and immunoglobulin E production in transgenic mice over expressing NK1 T cells. J. Exp. Med. 1996, 184, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Reardon, C.A.; Blachowicz, L.; Lukens, J.; Nissenbaum, M.; Getz, G.S. Genetic background selectively influences innominate artery atherosclerosis: Immune system deficiency as a probe. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Tupin, E.; Kronenberg, K. Activation of natural killer T cells by glycolipids. Meth. Enzymol. 2006, 417, 185–201. [Google Scholar] [PubMed]
- Le Boeuf, R.C.; Caldwell, M.; Kirk, E. Regulation by nutritional status of lipids and apolipoproteins A-I, A-II, and A-IV in inbred mice. J. Lipid Res. 1994, 35, 121–133. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| VLDLc | LDLc | HDLc | |
|---|---|---|---|
| mmol/L | |||
| Vα14tg/LDLR−/− | 17.7 ± 4.0 | 15.7 ± 1.5 | 2.4 ± 0.2 |
| LDLR−/− | 13.2 ± 2.4 | 12.3 ± 1.6 | 2.3 ± 0.3 |
| Jα18−/−LDLR−/− | 27.9 ± 2.7 * | 18.6 ± 0.5 * | 2.5 ± 0.2 |
| CD1d−/−LDLR−/− | 12.2 ± 0.8† | 14.4 ± 0.8‡ | 2.9 ± 0.4 |
| Hepatic Triglyceride Secretion mg Triglyceride/dL min−1 mL Plasma−1 g Liver−1 | |
|---|---|
| Vα14tg, LDLR−/− | 0.77 ± 0.33 |
| Jα18−/−LDLR−/− | 1.50 ± 0.65 |
| CD1d−/−LDLR−/− | 1.38 ± 0.30 |
| 4 Weeks WTD | 12 Weeks WTD | ||||
|---|---|---|---|---|---|
| AA | AR | IA | AA | AR | |
| Vα14tg, LDLR−/− | 2.0 ± 0.2 | 1.8 ± 0.2 | 3.9 ± 0.7 | 4.7 ± 0.5 | 4.2 ± 0.3 |
| LDLR−/− | 2.1 ± 0.2 | 1.7 ± 0.2 | 1.7 ± 0.4 | 4.4 ± 0.5 | 4.5 ± 0.3 |
| Jα18−/−LDLR−/− | 1.8 ± 0.1 | 1.6 ± 0.1 | 1.4 ± 0.4 | 4.1 ± 0.3 | 3.5 ± 0.2 |
| CD1d−/−LDLR−/− | 2.2 ± 0.2 | 2.2 ± 0.2 | 2.0 ± 0.3 | 3.6 ± 0.3 | 4.3 ± 0.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
VanderLaan, P.A.; Reardon, C.A.; Cabana, V.G.; Wang, C.-R.; Getz, G.S. Invariant Natural Killer T-Cells and Total CD1d Restricted Cells Differentially Influence Lipid Metabolism and Atherosclerosis in Low Density Receptor Deficient Mice. Int. J. Mol. Sci. 2019, 20, 4566. https://doi.org/10.3390/ijms20184566
VanderLaan PA, Reardon CA, Cabana VG, Wang C-R, Getz GS. Invariant Natural Killer T-Cells and Total CD1d Restricted Cells Differentially Influence Lipid Metabolism and Atherosclerosis in Low Density Receptor Deficient Mice. International Journal of Molecular Sciences. 2019; 20(18):4566. https://doi.org/10.3390/ijms20184566
Chicago/Turabian StyleVanderLaan, Paul A., Catherine A. Reardon, Veneracion G. Cabana, Chyung-Ru Wang, and Godfrey S. Getz. 2019. "Invariant Natural Killer T-Cells and Total CD1d Restricted Cells Differentially Influence Lipid Metabolism and Atherosclerosis in Low Density Receptor Deficient Mice" International Journal of Molecular Sciences 20, no. 18: 4566. https://doi.org/10.3390/ijms20184566