Rev-erbα Inhibits Proliferation and Promotes Apoptosis of Preadipocytes through the Agonist GSK4112


Abstract
1. Introduction
2. Results
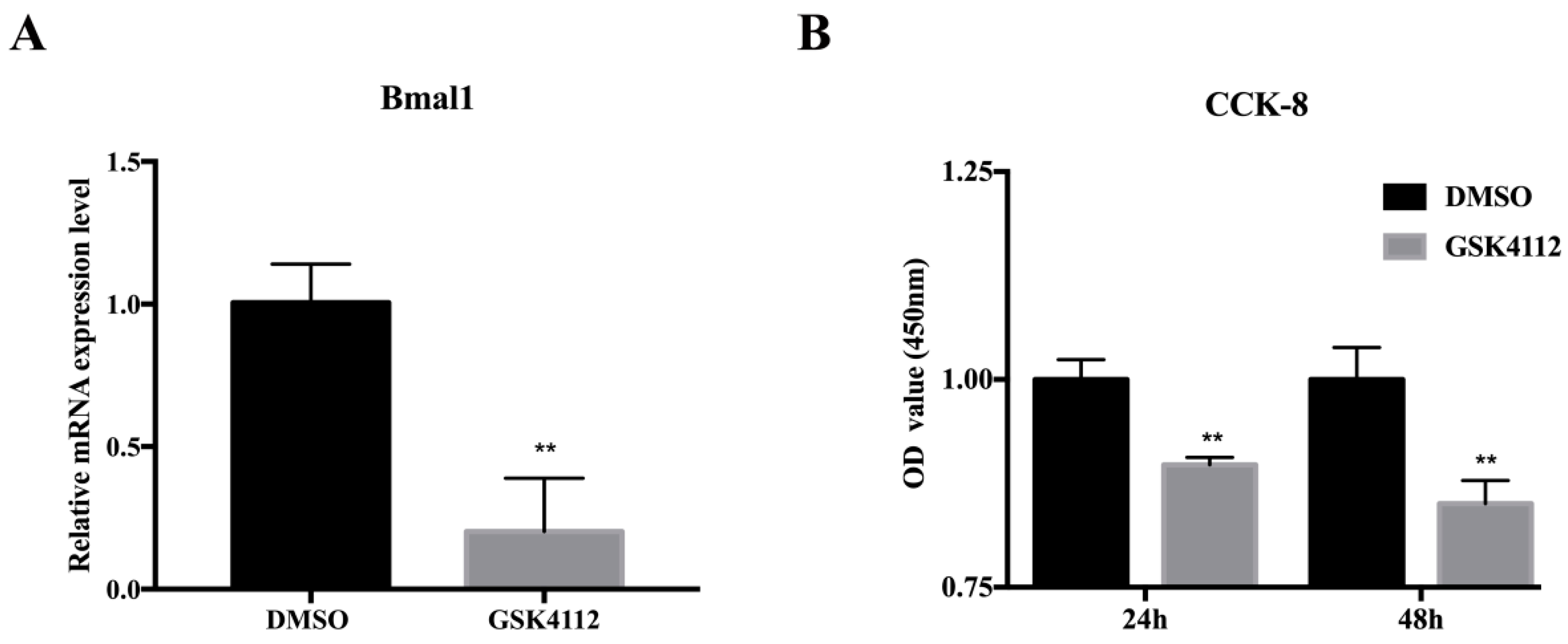
2.1. The Rev-erbα Agonist GSK4112 Inhibited Pre-Adipocyte Viability
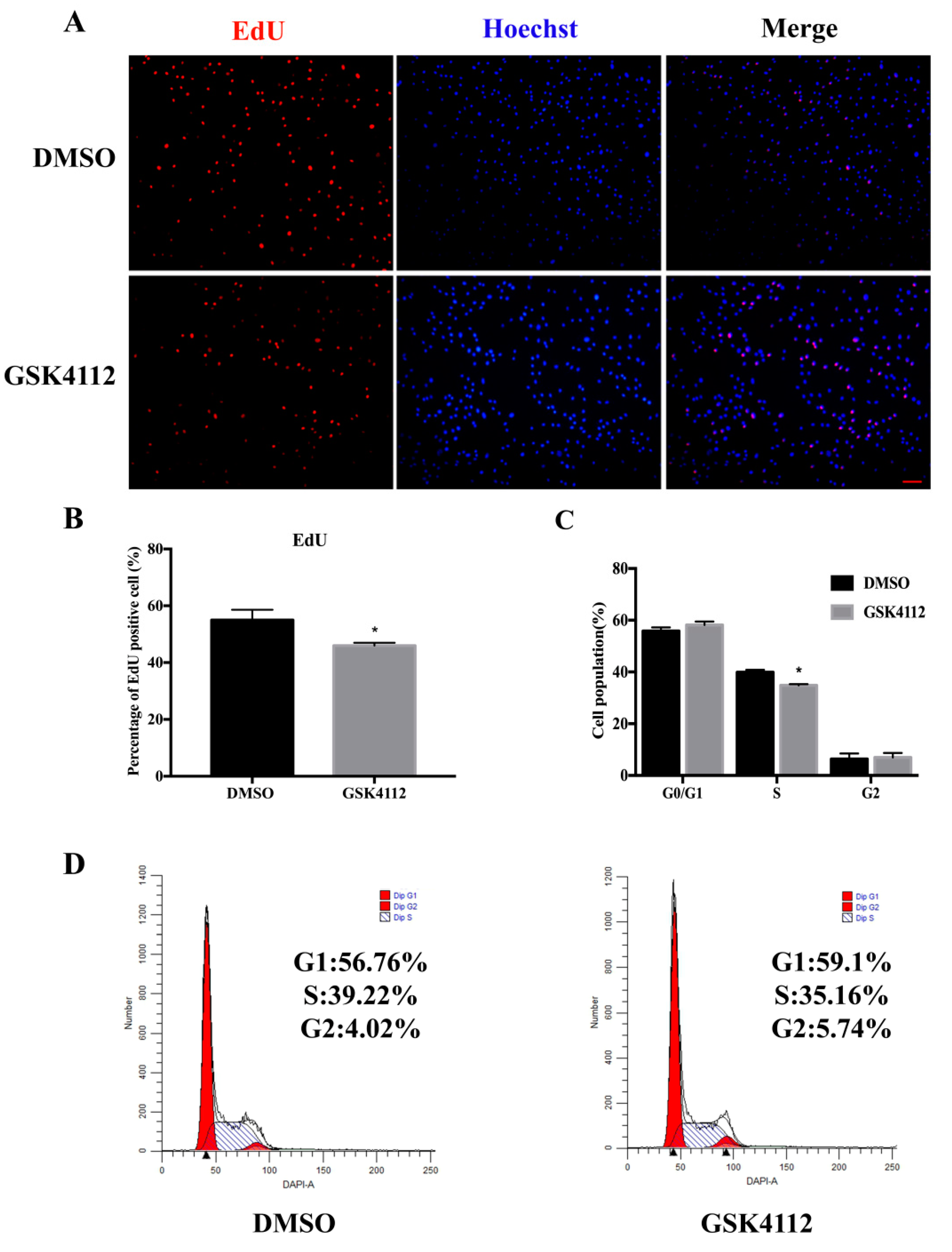
2.2. The Rev-erbα Agonist GSK4112 Inhibited Cell Proliferation
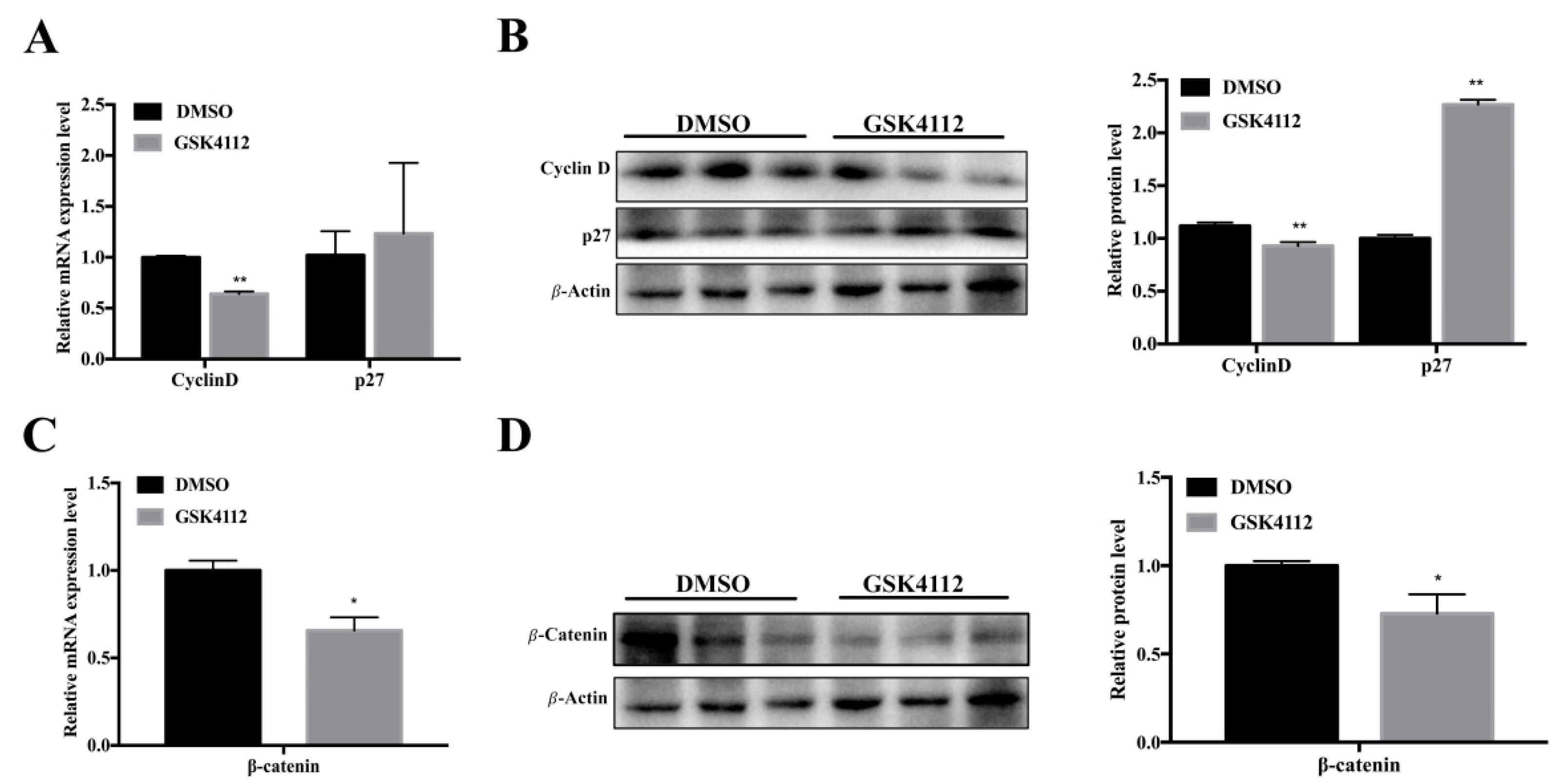
2.3. Rev-erbα Inhibited Proliferation of 3T3-L1 Cells through the Wnt Signaling Pathway
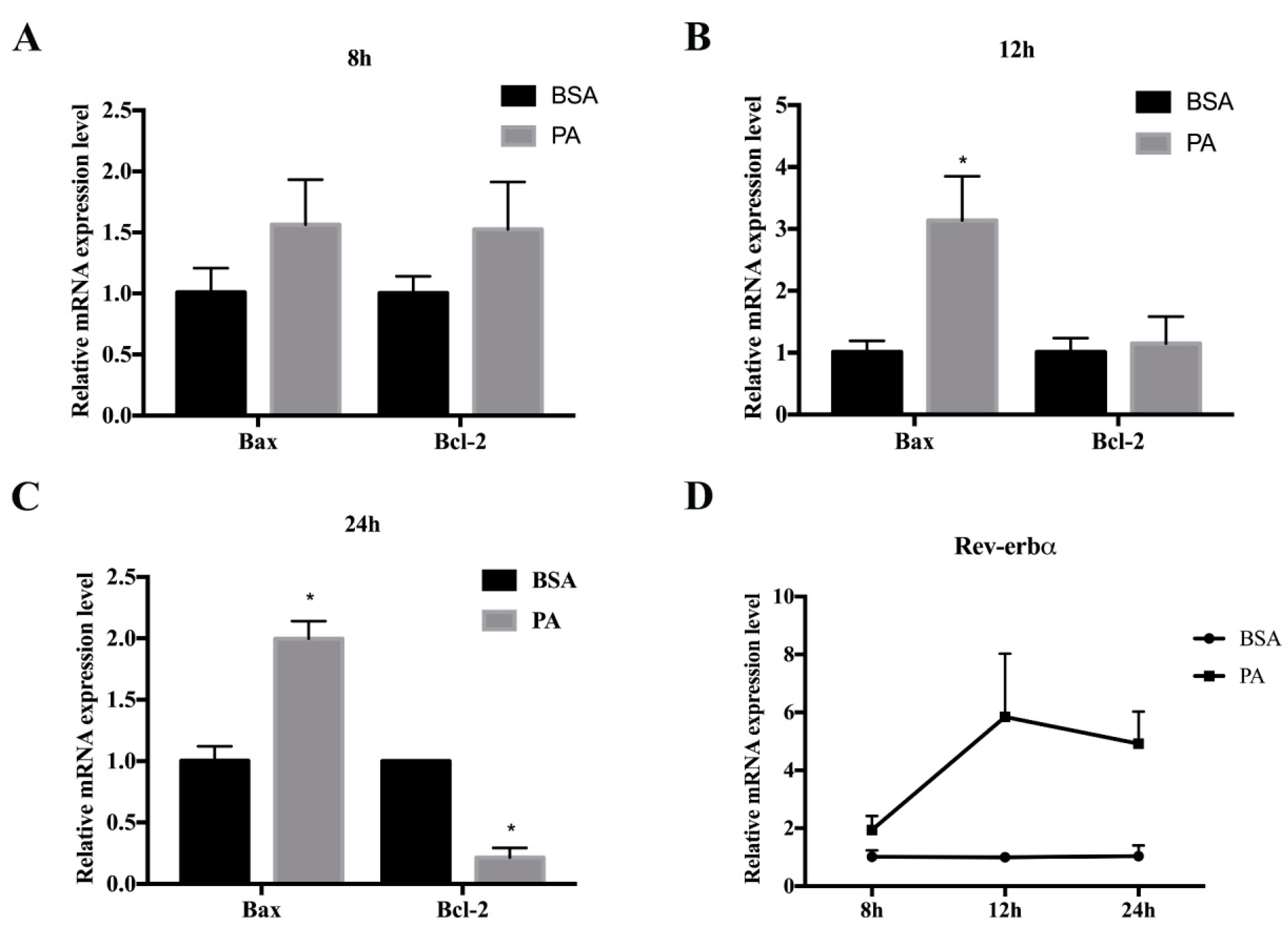
2.4. Cell Model of Palmitate-Induced 3T3-L1 Preadipocyte Apoptosis
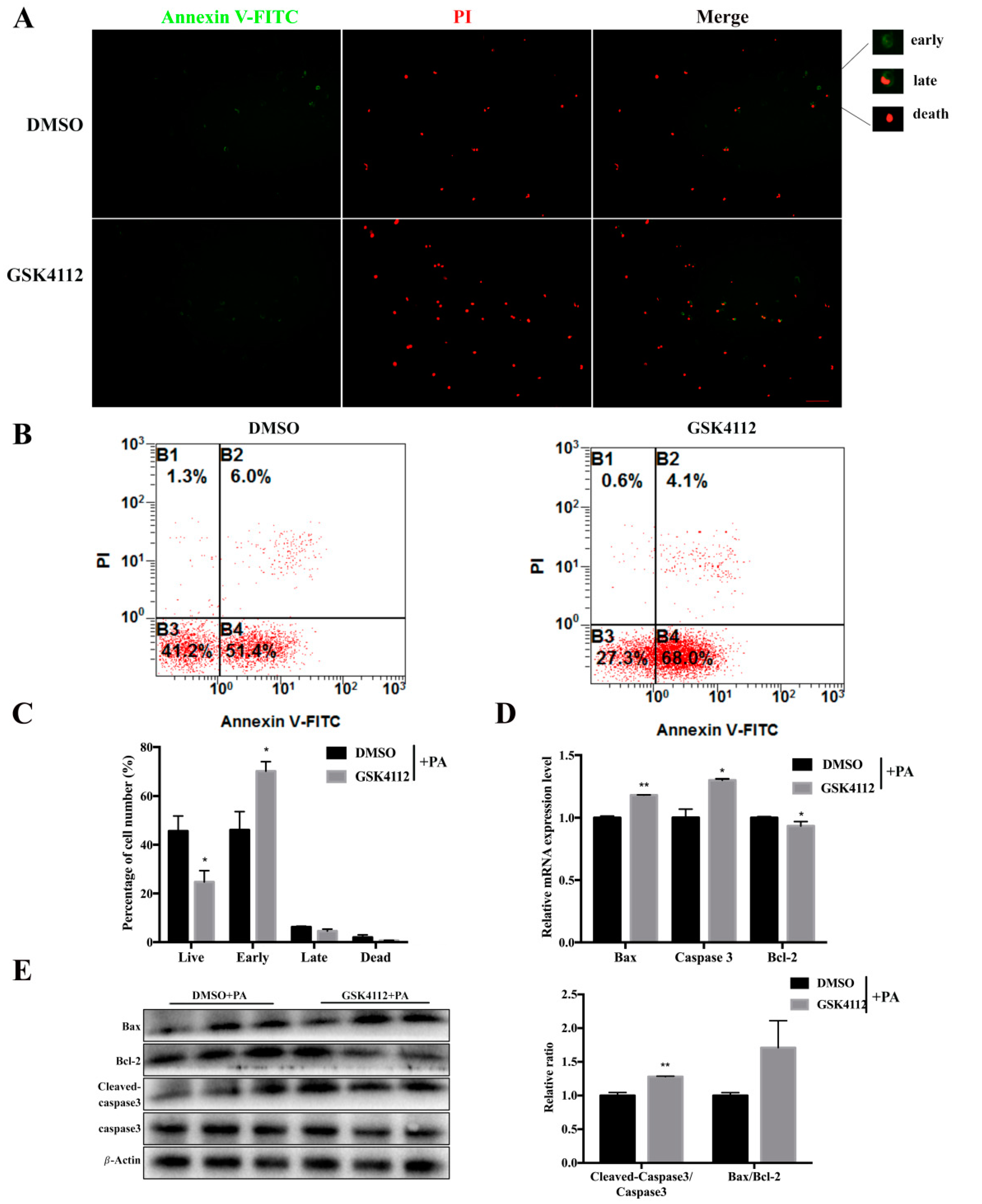
2.5. Rev-erbα Agonist GSK4112 Aggravated Palmitate-Induced Preadipocyte Apoptosis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatment
4.3. CCK-8 Assay
4.4. EdU Imaging Assay
4.5. Flow Cytometry
4.6. Apoptosis Assessment
4.7. RNA Isolation and Quantitative Real-Time PCR
4.8. Western Blot Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kuchta, K.F. Pathophysiologic changes of obesity. Anesthesiol. Clin. North. Am. 2005, 23, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Van Gaal, L.F.; Mertens, I.L.; De Block, C.E. Mechanisms linking obesity with cardiovascular disease. Nature 2006, 444, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Petrus, P.; Mejhert, N.; Corrales, P.; Lecoutre, S.; Li, Q.; Maldonado, E.; Kulyté, A.; Lopez, Y.; Campbell, M.; Acosta, J.R. Transforming Growth Factor-β3 Regulates Adipocyte Number in Subcutaneous White Adipose Tissue. Cell Rep. 2018, 25, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, E.; Church, C.D.; Holtrup, B.; Colman, L.; Rodeheffer, M.S. Rapid Depot-Specific Activation of Adipocyte Precursor Cells at the Onset of Obesity. Nat. Cell Biol. 2015, 17, 376–385. [Google Scholar] [CrossRef]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Stephane, G.; Yu-Hua, T.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar]
- Kojetin, D.J.; Burris, T.P. REV-ERB and ROR nuclear receptors as drug targets. Nat. Rev. Drug Discov. 2014, 13, 197–216. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Marhon, S.A.; Zhang, Y.; Steger, D.J.; Won, K.J.; Lazar, M.A. Rev-erbα dynamically modulates chromatin looping to control circadian gene transcription. Science 2018, 359, 1274–1277. [Google Scholar] [CrossRef]
- Anne, B.; Dan, F.; Everett, L.J.; Briggs, E.R.; Mullican, S.E.; Fenfen, W.; Jennifer, J.; Lazar, M.A. Rev-erbα and Rev-erbβ coordinately protect the circadian clock and normal metabolic function. Genes Dev. 2012, 26, 657–667. [Google Scholar]
- Estelle, W.; Yasmine, S.; Solt, L.A.; Christian, D.; Steve, L.; Jér?Me, E.; Hesselink, M.K.C.; Charlotte, P.; Stéphane, D.; Youseung, S. Rev-erb-α modulates skeletal muscle oxidative capacity by regulating mitochondrial biogenesis and autophagy. Nat. Med. 2013, 19, 1039–1046. [Google Scholar]
- Nicolas, P.; Francesca, D.; Luis, L.M.; Joszef, Z.; Denis, D.; Urs, A.; Ueli, S. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar]
- Ramakrishnan, S.N.; Patrick, L.; Burke, L.J.; Muscat, G.E.O. Rev-erbbeta regulates the expression of genes involved in lipid absorption in skeletal muscle cells: Evidence for cross-talk between orphan nuclear receptors and myokines. J. Biol. Chem. 2005, 280, 8651. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, S.N.; Muscat, G.E. The orphan Rev-erb nuclear receptors: A link between metabolism, circadian rhythm and inflammation? Nucl. Recept. Signal. 2006, 4, nrs-04009. [Google Scholar] [CrossRef] [PubMed]
- Xiaoyong, Y.; Michael, D.; Yu, R.T.; Bookout, A.L.; Weimin, H.; Marty, S.; Mangelsdorf, D.J.; Evans, R.M. Nuclear receptor expression links the circadian clock to metabolism. Cell 2006, 126, 801–810. [Google Scholar]
- Laitinen, S.; Fontaine, C.; Fruchart, J.C.; Staels, B. The role of the orphan nuclear receptor Rev-Erbα in adipocyte differentiation and function. Biochimie 2005, 87, 21–25. [Google Scholar] [CrossRef]
- He, Y.; Lin, F.; Chen, Y.; Tan, Z.; Bai, D.; Zhao, Q. Overexpression of the Circadian Clock Gene Rev-erbα Affects Murine Bone Mesenchymal Stem Cell Proliferation and Osteogenesis. Stem Cells Dev. 2015, 24, 1194–1204. [Google Scholar] [CrossRef]
- Sulli, G.; Rommel, A.; Wang, X.; Kolar, M.J.; Puca, F.; Saghatelian, A.; Plikus, M.V.; Verma, I.M.; Panda, S. Pharmacological activation of REV-ERBs is lethal in cancer and oncogene-induced senescence. Nature 2018, 553, 351. [Google Scholar] [CrossRef]
- Delezie, J.; Dumont, S.; Dardente, H.; Oudart, H.; Gréchez-Cassiau, A.; Klosen, P.; Teboul, M.; Delaunay, F.; Pévet, P.; Challet, E. The nuclear receptor REV-ERBα is required for the daily balance of carbohydrate and lipid metabolism. Faseb. J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2012, 26, 3321–3325. [Google Scholar] [CrossRef]
- Duez, H.; van der Veen, J.N.; Duhem, C.; Pourcet, B.; Touvier, T.; Fontaine, C.; Derudas, B.; Baugé, E.; Havinga, R.; Bloks, V.W.; et al. Regulation of bile acid synthesis by the nuclear receptor Rev-erbα. Gastroenterology 2008, 135, 689–698. [Google Scholar] [CrossRef]
- Sitaula, S.; Zhang, J.; Ruiz, F.; Burris, T.P. Rev-erb Regulation of Cholesterologenesis. Biochem. Pharmacol. 2017, 131, 68–77. [Google Scholar] [CrossRef]
- Han, C.; Xuan, Z.; Megumi, H.; Ruth, T.Y.; Grant, D.B.; Michael, T.L.; Ling-Wa, C.; Luciano, D.; Annette, R.A.; Christopher, K.G. Regulation of circadian behaviour and metabolism by REV-ERB-α and REV-ERB-β. Nature 2012, 485, 123. [Google Scholar]
- Chawla, A.; Lazar, M.A. Induction of Rev-ErbA alpha, an orphan receptor encoded on the opposite strand of the alpha-thyroid hormone receptor gene, during adipocyte differentiation. J. Biol. Chem. 1993, 268, 16265–16269. [Google Scholar]
- Kojetin, D.; Wang, Y.; Kamenecka, T.M.; Burris, T.P. Identification of SR8278, a synthetic antagonist of the nuclear heme receptor REV-ERB. ACS Chem. Biol. 2011, 6, 131–134. [Google Scholar] [CrossRef]
- Kojetin, D.J.; Burris, T.P. A role for rev-erbα ligands in regulation of adipogenesis. Curr. Pharm. Des. 2011, 17, 320–324. [Google Scholar] [CrossRef]
- Naresh, K.; Solt, L.A.; Yongjun, W.; Rogers, P.M.; Gargi, B.; Kamenecka, T.M.; Stayrook, K.R.; Christine, C.; Elizabeth, F.Z.; Gimble, J.M. Regulation of adipogenesis by natural and synthetic REV-ERB ligands. Endocrinology 2010, 151, 3015–3025. [Google Scholar]
- Kioussi, C.; Briata, P.; Baek, S.H.; Rose, D.W.; Hamblet, N.S.; Herman, T.; Ohgi, K.A.; Lin, C.; Gleiberman, A.; Wang, J. Identification of a Wnt/Dvl/β-Catenin→Pitx2 Pathway Mediating Cell-Type-Specific Proliferation during Development. Cell 2002, 111, 673–685. [Google Scholar] [CrossRef]
- Qu, Q.; Sun, G. Orphan nuclear receptor TLX activates Wnt/beta-catenin signalling to stimulate neural stem cell proliferation and self-renewal. Nat. Cell Biol. 2010, 12, 1–9. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Tetsu, O.; Mccormick, F. Beta-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef]
- Wang, J.; Yin, L.; Lazar, M.A. The Orphan Nuclear Receptor Rev-erbα Regulates Circadian Expression of Plasminogen Activator Inhibitor Type 1. J. Biol. Chem. 2006, 281, 33842–33848. [Google Scholar] [CrossRef]
- Guo, D.K.; Zhu, Y.; Sun, H.Y.; Xu, X.Y.; Zhang, S.; Hao, Z.B.; Wang, G.H.; Mu, C.C.; Ren, H.G. Pharmacological activation of REV-ERBalpha represses LPS-induced microglial activation through the NF-kappaB pathway. Acta Pharmacol. Sin. 2019, 40, 26–34. [Google Scholar] [CrossRef]
- Alenzi, F.Q.B. Links between apoptosis, proliferation and the cell cycle. Br. J. Biomed. Sci. 2004, 61, 99–102. [Google Scholar] [CrossRef]
- Grant, D.; Yin, L.; Collins, J.L.; Parks, D.J.; Orband-Miller, L.A.; Wisely, G.B.; Joshi, S.; Lazar, M.A.; Willson, T.M.; Zuercher, W.J. GSK4112, a small molecule chemical probe for the cell biology of the nuclear heme receptor rev-erbα. ACS Chem. Biol. 2010, 5, 925–932. [Google Scholar] [CrossRef]
- Yongjun, W.; Douglas, K.; Burris, T.P. Anti-proliferative actions of a synthetic REV-ERBα/β agonist in breast cancer cells. Biochem. Pharmacol. 2015, 96, 315–322. [Google Scholar]
- Lu, G.; Zhenjiang, L.; Wei, J.; Zhongjie, Z.; Chao, S. Foxc2 enhances proliferation and inhibits apoptosis through activating Akt/mTORC1 signaling pathway in mouse preadipocytes. J. Lipid Res. 2015, 56, 1471–1480. [Google Scholar]
- Yanling, W.; Masaaki, S.; Yuan, G.; Tore, B.; Jan, N. Protein kinase A-mediated cell proliferation in brown preadipocytes is independent of Erk1/2, PI3K and mTOR. Exp. Cell Res. 2014, 328, 143–155. [Google Scholar]
- Zhu, Z.; Hua, B.; Xu, L.; Yuan, G.; Li, E.; Li, X.; Sun, N.; Yan, Z.; Lu, C.; Qian, R. CLOCK promotes 3T3-L1 cell proliferation via Wnt signaling. IUBMB Life 2016, 68, 557–568. [Google Scholar] [CrossRef]
- Junying, Y.; Guido, K. Alternative cell death mechanisms in development and beyond. Genes Dev. 2010, 24, 2592–2602. [Google Scholar]
- Guo, W.; Wong, S.; Xie, W.; Lei, T.; Luo, Z. Palmitate modulates intracellular signaling, induces endoplasmic reticulum stress, and causes apoptosis in mouse 3T3-L1 and rat primary preadipocytes. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E576. [Google Scholar] [CrossRef]
- Antonsson, B.; Montessuit, S.; Lauper, S.; Eskes, R.; Martinou, J.C. Bax oligomerization is required for channel-forming activity in liposomes and to trigger cytochrome c release from mitochondria. Biochem. J. 2000, 345, 271–278. [Google Scholar] [CrossRef]
- Llambi, F.; Wang, Y.M.; Victor, B.; Yang, M.; Schneider, D.; Gingras, S.; Parsons, M.; Zheng, J.; Brown, S.; Pelletier, S. BOK Is a Non-canonical BCL-2 Family Effector of Apoptosis Regulated by ER-Associated Degradation. Cell 2016, 165, 421–433. [Google Scholar] [CrossRef]
- Suzanne, C.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneName | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| Cyclin D | TAGGCCCTCAGCCTCACTC | CCACCCCTGGGATAAAGCAC |
| p27 | AGAAGCACTGCCGGGATATG | GACCCAATTAAAGGCACCGC |
| Bmal1 | GGCTGTCATCATGAGCCTCT | TGAGGAAACACTGGAGCAGG |
| Bcl-2 | GTCGCTACCGTCGTGACTTC | CAGACATGCACCTACCCAGC |
| Bax | TGAAGACAGGGGCCTTTTTG | AATTCGCCGGAGACACTCG |
| Caspase-3 | ATGGAGAACAACAAAACCTCAGT | TTGCTCCCATGTATGGTCTTTAC |
| Rev-erbα | AACGGATGCTTGCCGAGAT | GGAGCCAGAGGTGGGATGT |
| β-actin | GTCCCTGACCCTCCCAAAAG | GCTGCCTCAACACCTCAACCC |
| β-catenin | TCCCATCCACGCAGTTTGAC | TCCTCATCGTTTAGCAGTTTTGT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, G.; Zhou, X.; Hu, Y.; Shi, S.; Yang, G. Rev-erbα Inhibits Proliferation and Promotes Apoptosis of Preadipocytes through the Agonist GSK4112. Int. J. Mol. Sci. 2019, 20, 4524. https://doi.org/10.3390/ijms20184524
Chu G, Zhou X, Hu Y, Shi S, Yang G. Rev-erbα Inhibits Proliferation and Promotes Apoptosis of Preadipocytes through the Agonist GSK4112. International Journal of Molecular Sciences. 2019; 20(18):4524. https://doi.org/10.3390/ijms20184524
Chicago/Turabian StyleChu, Guiyan, Xiaoge Zhou, Yamei Hu, Shengjie Shi, and Gongshe Yang. 2019. "Rev-erbα Inhibits Proliferation and Promotes Apoptosis of Preadipocytes through the Agonist GSK4112" International Journal of Molecular Sciences 20, no. 18: 4524. https://doi.org/10.3390/ijms20184524
APA StyleChu, G., Zhou, X., Hu, Y., Shi, S., & Yang, G. (2019). Rev-erbα Inhibits Proliferation and Promotes Apoptosis of Preadipocytes through the Agonist GSK4112. International Journal of Molecular Sciences, 20(18), 4524. https://doi.org/10.3390/ijms20184524