P1G10, the Proteolytic Fraction from Vasconcellea cundinamarcensis, Stimulates Tissue Repair after Acute Exposure to Ultraviolet B Radiation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
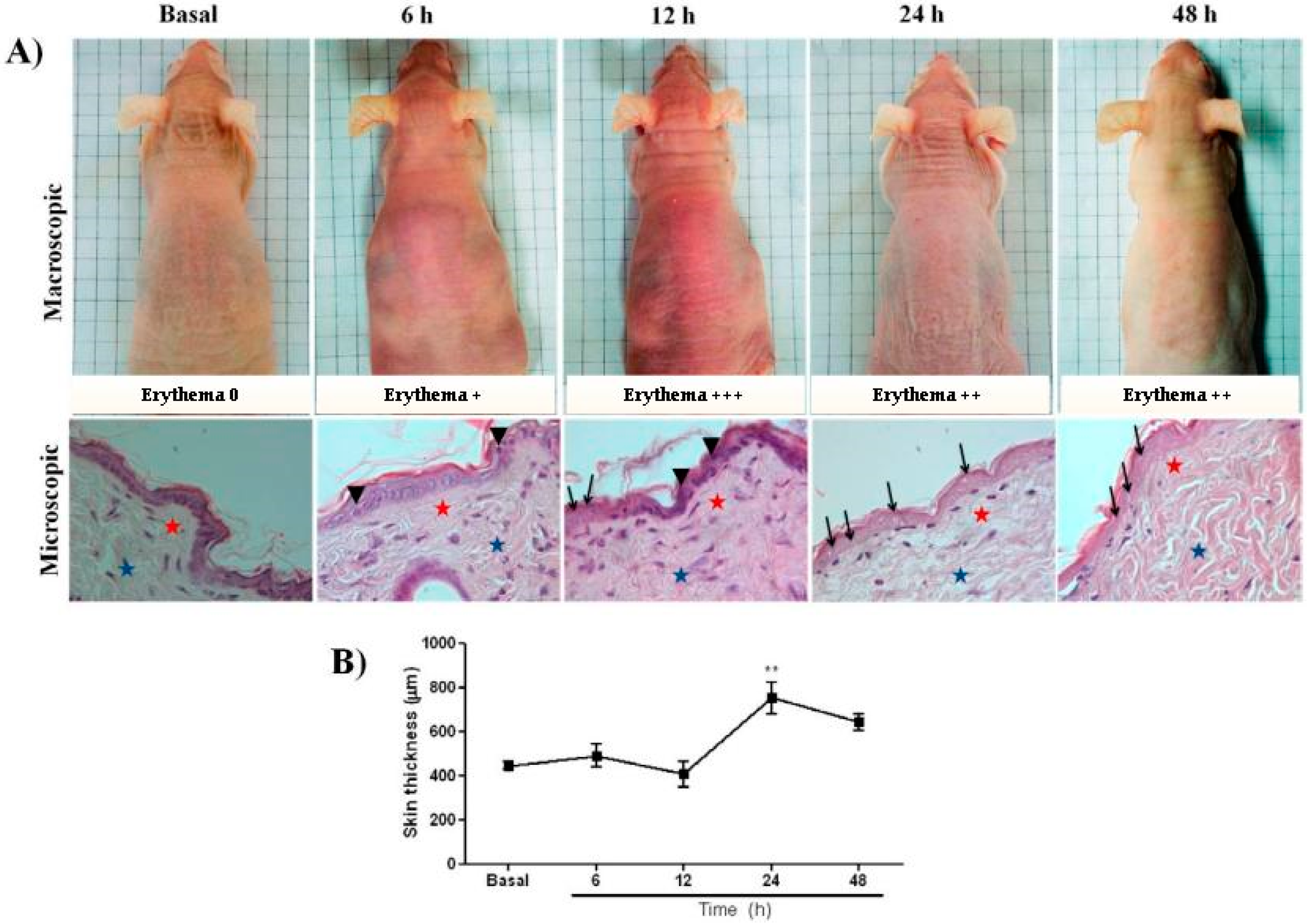
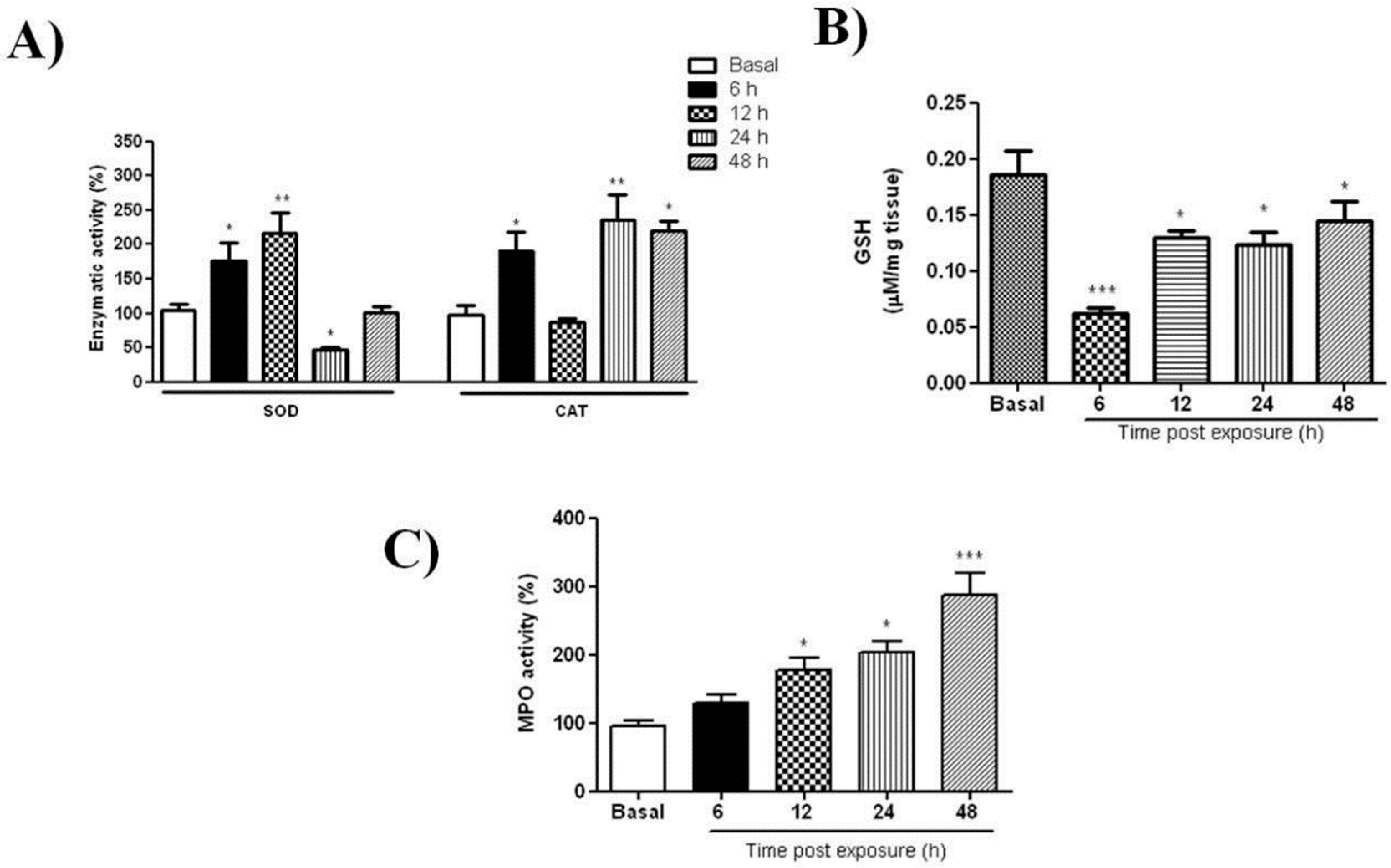
2.1. UVB Radiation-Induced Changes
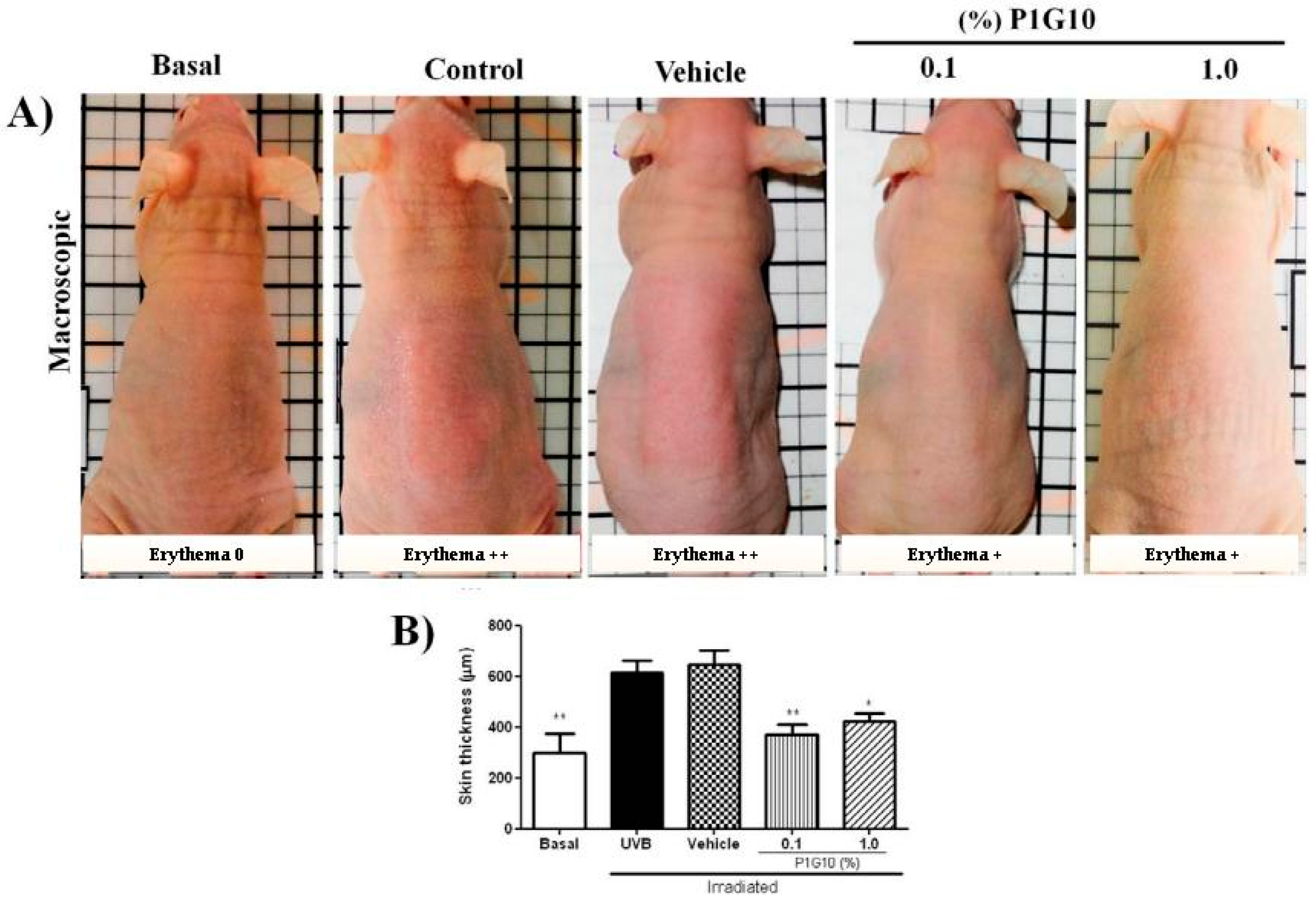
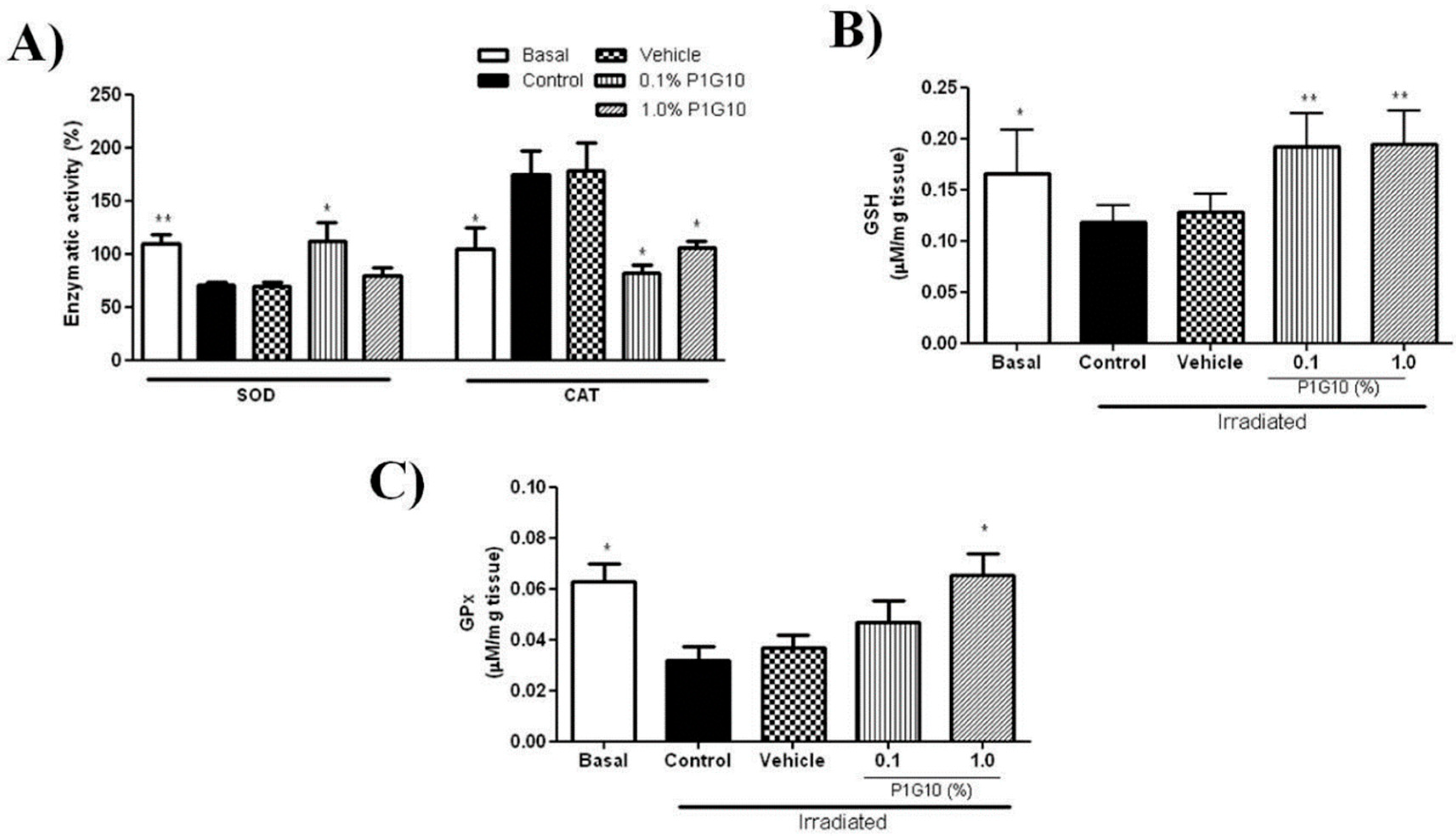
2.2. Effect of P1G10 on UVB Irradiated Mice
2.2.1. P1G10 Modulates UVB-Induced Antioxidant Responses
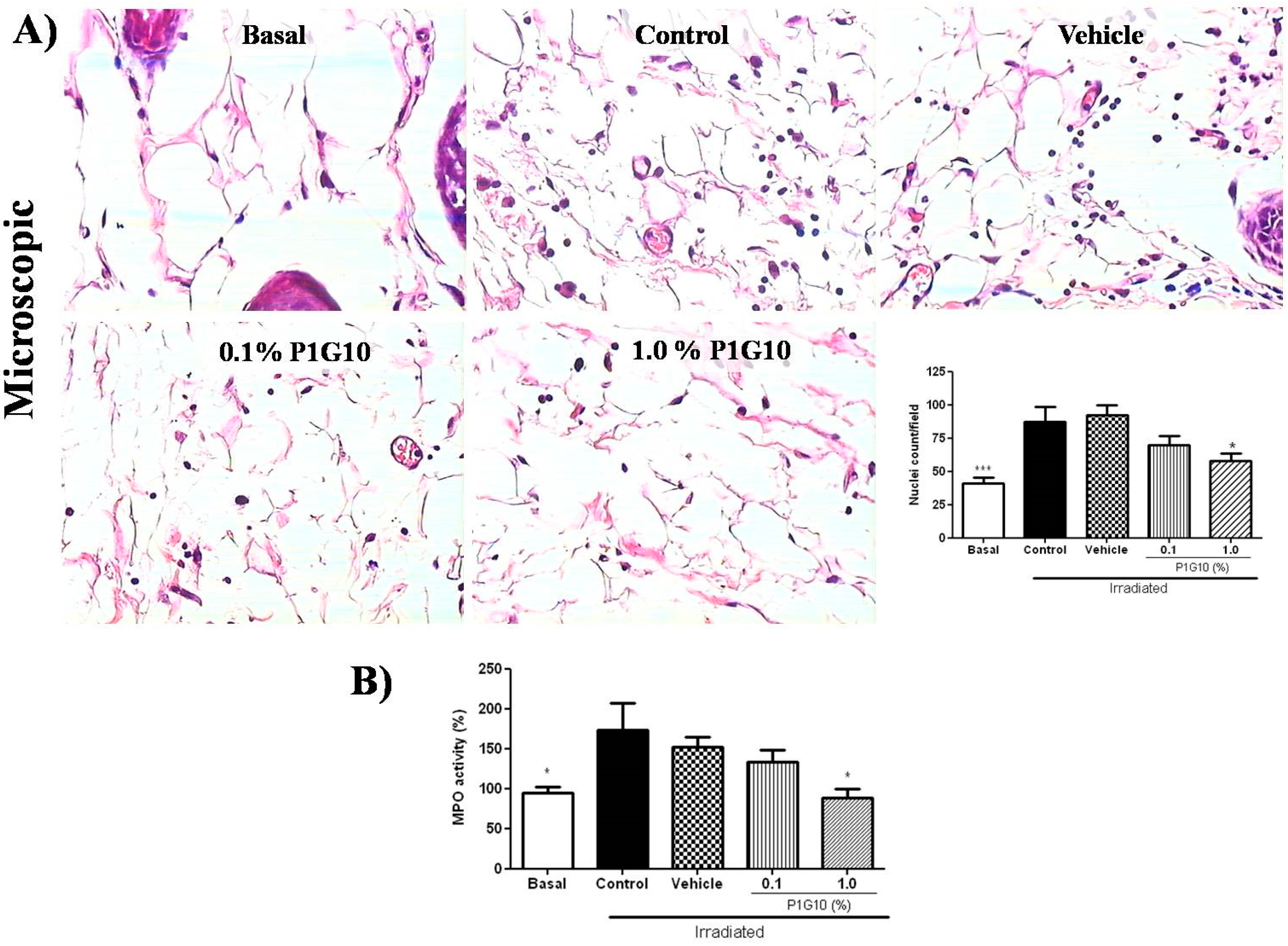
2.2.2. P1G10 Reduced Infiltration and Activation of Inflammatory Cells
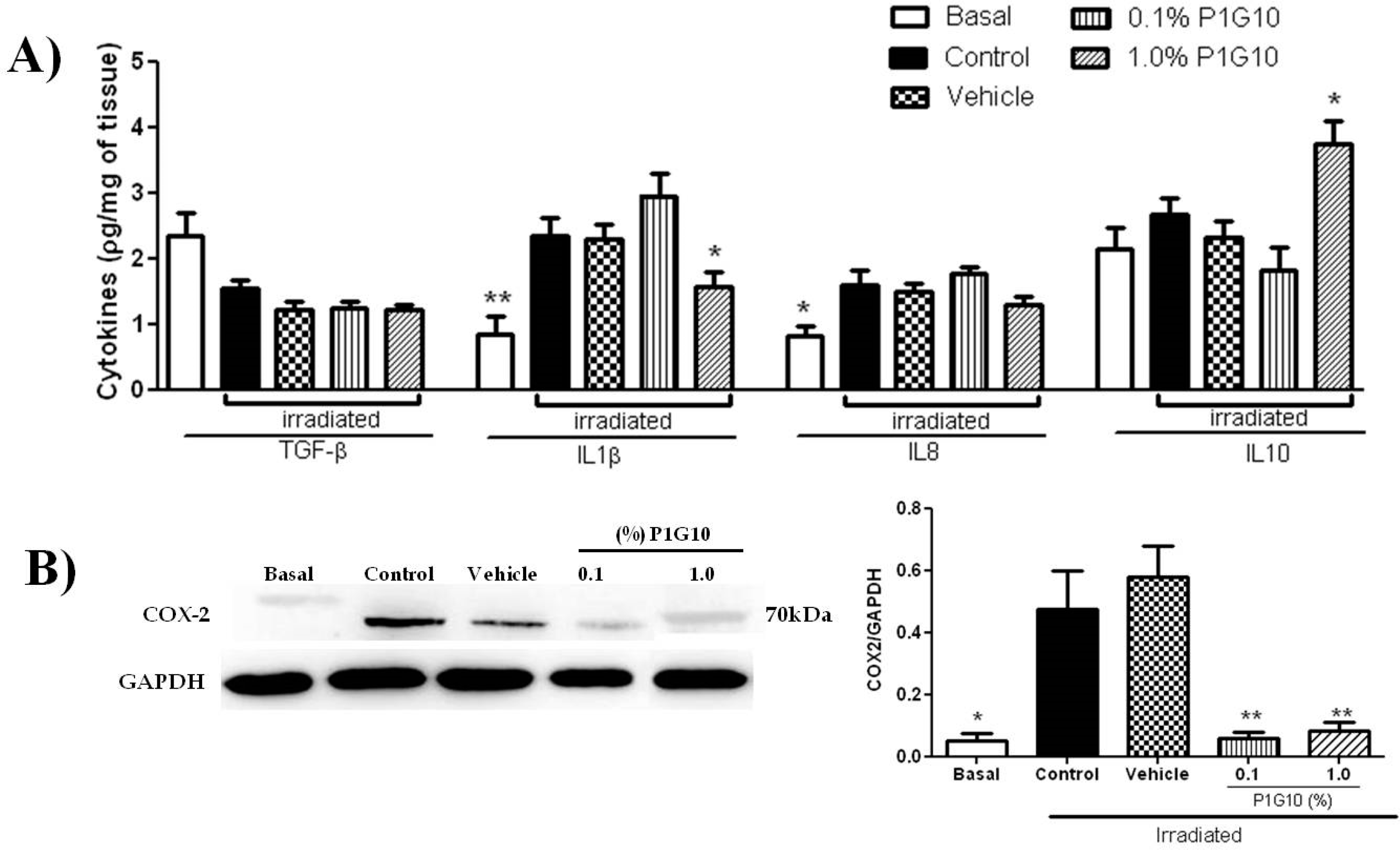
2.2.3. P1G10 Modulates Inflammatory Mediators Induced by UVB
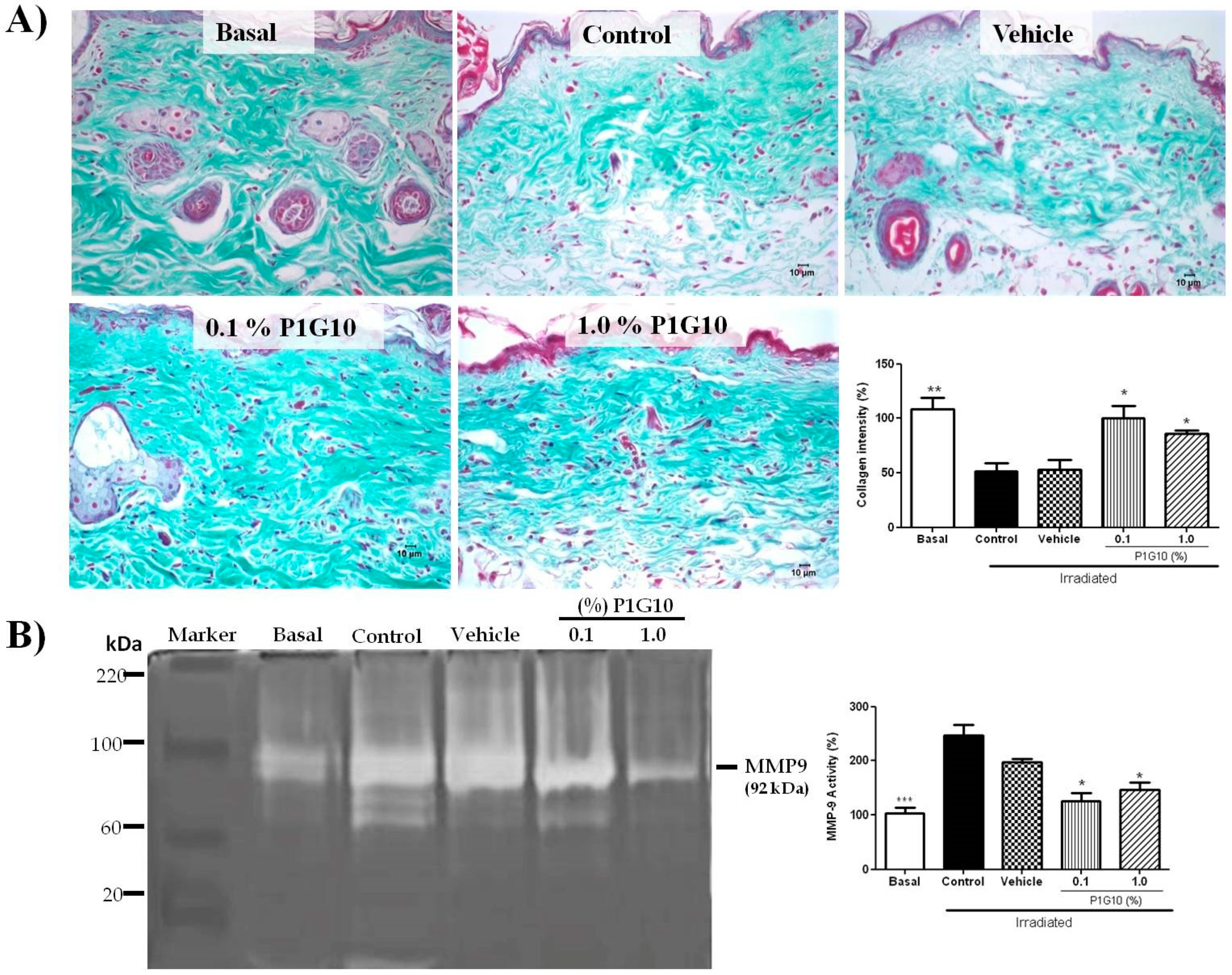
2.2.4. P1G10 Prevents UVB-Induced Collagen Degradation
2.2.5. P1G10 Reduces Oxidative Damage
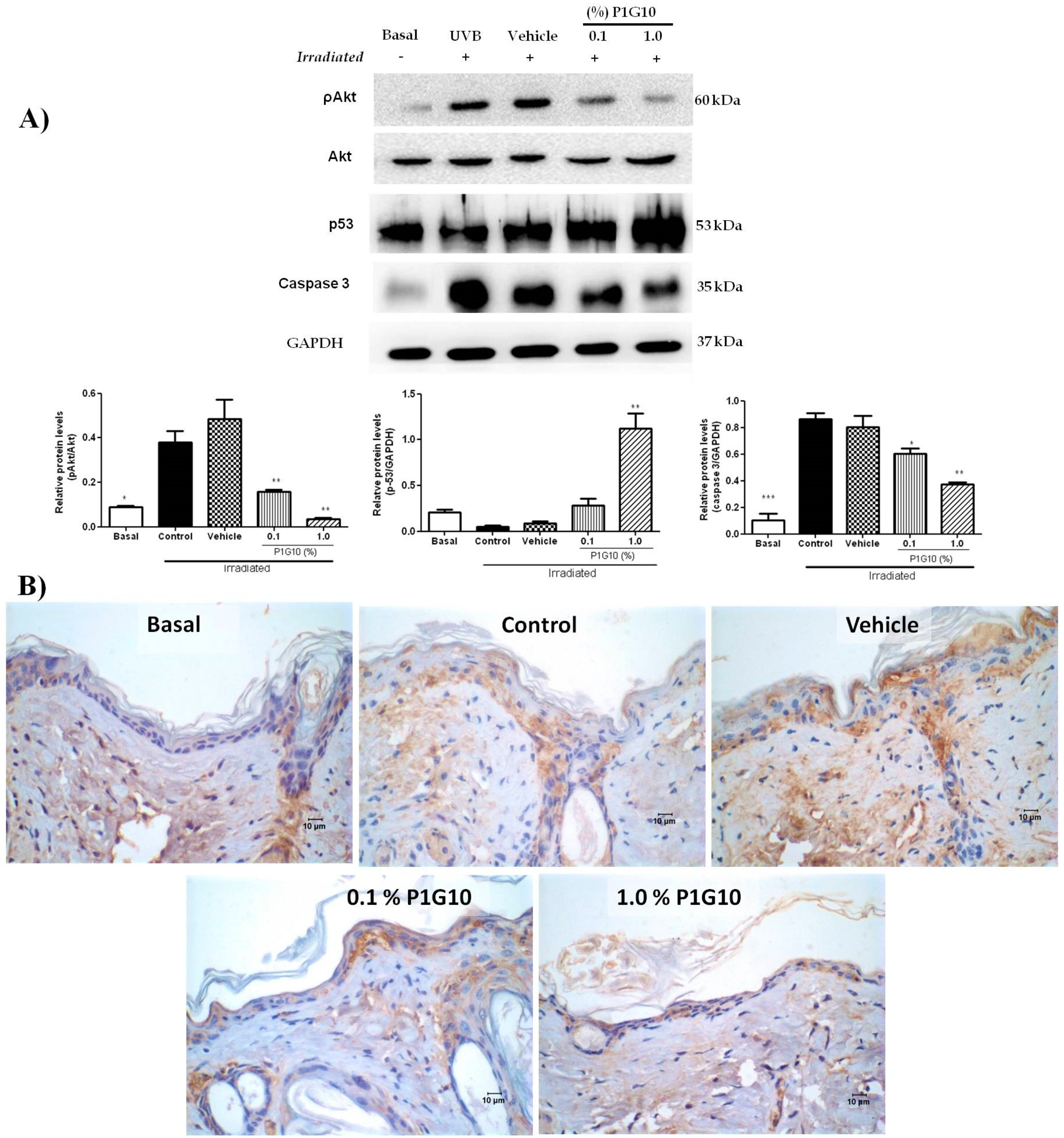
2.2.6. P1G10 Affects Levels of ROS-Related Signaling Proteins
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals
4.3. Production of P1G10
4.4. P1G10 Formulation
4.5. Antioxidant Activity of P1G10
4.6. Exposure to Single UVB Dose
4.7. Analysis of Oxidative Stress
4.8. Generation of Intracellular Reactive Oxygen Species (ROS)
4.8.1. Superoxide Dismutase Activity (SOD)
4.8.2. Catalase Activity (CAT)
4.8.3. Glutathione Peroxidase Activity (GSH-Px)
4.8.4. Reduced Glutathione Content (GSH)
4.8.5. Malondialdehyde Content (MDA)
4.9. Cytokines Analysis
4.10. Myeloperoxidase Activity (MPO)
4.11. Gelatin Zymography
4.12. Western Blot
4.13. Histological Analysis
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ishida, T.; Sakaguchi, I. Protection of human keratinocytes from UVB-induced inflammation using root extract of Lithospermum erythrorhizon. Biol. Pharm. Bull. 2007, 30, 928–934. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Woodcock, A.; Magnus, I.A. The sunburn cell in mouse skin: Preliminary quantitative studies on its production. Br. J. Dermatol. 1976, 95, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, N.J.; Song, L.; Doig, J.; Ritchie, A.M.; Brownstein, D.G.; Melton, D.W. Topical thymidine dinucleotide application protects against uvb-induced skin cancer in mice with dna repair gene (ERCC1)-deficient skin. DNA Repair. 2009, 8, 664–671. [Google Scholar] [CrossRef] [PubMed]
- Masaki, H.; Okano, Y.; Sakurai, H. Differential role of catalase and glutathione peroxidase in cultured human fibroblasts under exposure of H2O2 or ultraviolet B light. Arch. Dermatol. Res. 1998, 290, 113–118. [Google Scholar] [CrossRef]
- Morison, W.L.; Parrish, J.A.; Bloch, K.J.; Kruglert, I. In vivo effect of UV-B on lymphocyte function. Br. J. Dermatol. 1979, 101, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Liu-Smith, F.; Jia, J.; Zheng, Y. UV-induced molecular signaling differences in melanoma and non-melanoma skin cancer. Adv. Exp. Med. Biol. 2017, 996, 27–40. [Google Scholar] [PubMed]
- Melnikova, V.O.; Ananthaswamy, H.N. Cellular and molecular events leading to the development of skin cancer. Mutat. Res. Mol. Mech. Mutagen. 2005, 571, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Liou, J.; Kuo, C.; Tsai, Y.; Lin, E.; Hsieh, C.; Lin, S.; Chen, B. UVB promotes the initiation of uveitic inflammatory injury in vivo and is attenuated by UV-blocking protection. Mol. Vis. 2017, 23, 219–227. [Google Scholar] [PubMed]
- Pearse, A.D.; Gaskell, S.A.; Marks, R. Epidermal changes in human skin following irradiation with either UVB or UVA. J. Investig. Dermatol. 1987, 88, 83–87. [Google Scholar] [CrossRef]
- Clydesdale, G.J.; Dandie, G.W.; Muller, H.K. Ultraviolet light induced injury: Immunological and inflammatory effects. Immunol. Cell Biol. 2001, 79, 547–568. [Google Scholar] [CrossRef]
- Filip, G.A.; Postescu, I.D.; Bolfa, P.; Catoi, C.; Muresan, A.; Clichici, S. Inhibition of UVB-induced skin phototoxicity by a grape seed extract as modulator of nitrosative stress, ERK/NF-κB signaling pathway and apoptosis, in SKH-1 mice. Food Chem. Toxicol. 2013, 57, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Darr, D.; Fridovich, I. Free radicals in cutaneous biology. J. Investig. Dermatol. 1994, 102, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Gresham, A.; Masferrer, J.; Chen, X.I.; Masferrer, J.; Chen, X. Increased synthesis of high-molecular-weight cPLA2 mediates synthesis of high-molecular-weight early UV-induced PGE in human skin. Am. J. Physiol-Cell. 1996, 270, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Bais, A.F.; Bernhard, G.; McKenzie, R.L.; Aucamp, P.J.; Young, P.J.; Ilyas, M.; Jöckel, P.; Deushi, M. Ozone–climate interactions and effects on solar ultraviolet radiation. Photochem. Photobiol. Sci. 2019, 18, 602–640. [Google Scholar] [CrossRef] [PubMed]
- Antón, M.; Serrano, A.; Cancillo, M.L.; Garcia, J.A. Relationship between erythemal irradiance and total solar irradiance in south-western Spain. J. Geophys. Res. Atmos. 2008, 113, D14208. [Google Scholar] [CrossRef]
- Saewan, N.; Jimtaisong, A. Natural products as photoprotection. J. Cosmet. Dermatol. 2015, 14, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.X.; Huang, H.H.; Chang, H.R.; Kuo, S.M. Evaluation of the UVB-screening capacity and restorative effects exerted by farnesol gel on UVB-caused sunburn. Environ. Toxicol. 2018, 33, 488–507. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S.; Knowland, J. Ultraviolet radiation screening compounds. Biol. Rev. 2007, 74, 311–345. [Google Scholar] [CrossRef]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef]
- Araujo e Silva, A.C.; de Oliveira Lemos, F.; Gomes, M.T.R.; Salas, C.E.; Lopes, M.T.P. Role of gastric acid inhibition, prostaglandins and endogenous-free thiol groups on the gastroprotective effect of a proteolytic fraction from Vasconcellea cundinamarcensis latex. J. Pharm. Pharmacol. 2015, 67, 133–141. [Google Scholar] [CrossRef]
- Mello, V.J.; Gomes, M.T.R.; Lemos, F.O.; Delfino, J.L.; Andrade, S.P.; Lopes, M.T.P.; Salas, C.E. The gastric ulcer protective and healing role of cysteine proteinases from Carica candamarcensis. Phytomedicine 2008, 15, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Sawane, M.; Kidoya, H. Apelin attenuates UVB-induced edema and inflammation by promoting vessel function. Am. J. Pathol. 2011, 179, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Cotran, R.S.; Pathak, M.A. The Pattern of vascular leakage induced by monochromatic uv irradiation in rats, guinea pigs and Hairless mice. J. Investig. Dermatol. 1968, 51, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Pence, B.C.; Naylor, M.F. Effects of Single-dose ultraviolet radiation on skin superoxide dismutase, catalase, and xanthine oxidase in Hairless mice. J. Investig. Dermatol. 1990, 95, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Iizawa, O.; Kato, T.; Tagami, H.; Akamatsu, H.; Niwa, Y. Long-term follow-up study of changes in lipid peroxide levels and the activity of superoxide dismutase, catalase and glutathione peroxidase in mouse skin after acute and chronic UV irradiation. Arch. Dermatol. Res. 1994, 286, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Sarrasague, M.M.; Barrado, D.A.; Zubillaga, M.; Hager, A.; De Paoli, T.; Boccio, J. Current concepts on glutathione metabolism. Usefulness of stable isotopes of its homeostasis. Acta Bioquim. Clin. Latinoam. 2006, 40, 45–51. [Google Scholar]
- Connor, M.J.; Wheeler, L.A. Depletion of cutaneous glutathione by radiation Photochem. Photobiol. 1987, 46, 239–245. [Google Scholar] [CrossRef]
- Griswold, D.E.; Connor, J.R.; Dalton, B.J.; Lee, J.C.; Simon, P.; Hillegass, L.; Sieg, D.J.; Hanna, N. Activation of the IL-1 gene in UV-irradiated mouse skin: Association with inflammatory sequelae and pharmacologic intervention. J. Investig. Dermatol. 1991, 97, 1019–1023. [Google Scholar] [CrossRef]
- Strzepa, A.; Pritchard, K.A.; Dittel, B.N. Myeloperoxidase: A new player in autoimmunity. Cell. Immunol. 2017, 317, 1–8. [Google Scholar] [CrossRef]
- Gomes, F.S.L.; Spínola, C.V.; Ribeiro, H.A.; Lopes, M.T.P.; Cassali, G.D.; Salas, C.E. Wound-healing activity of a proteolytic fraction from Carica candamarcensis on experimentally induced burn. Burns 2010, 36, 277–283. [Google Scholar] [CrossRef]
- Freitas, K.M.; Barcelos, L.S.; Caliari, M.V.; Salas, C.E.; Lopes, M.T.P. Healing activity of proteolytic fraction (P1G10) from Vasconcellea cundinamarcensis in a cutaneous wound excision model. Biomed. Pharmacother. 2017, 96, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Chen, H.C.; Lin, T.J.; Shih, I.C.; Wen, K.C. Michelia alba extract attenuates UVB-induced expression of matrix metalloproteinases via MAP kinase pathway in human dermal fibroblasts. Food Chem. Toxicol. 2012, 50, 4260–4269. [Google Scholar] [CrossRef] [PubMed]
- Sterenborg, H.J.C.M.; Putte, S.C.J.; Leun, J.C. The dose-response relationship of tumorigenesis by ultraviolet radiation of 254 nm. Photochem. Photobiol. 2008, 47, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.M.A.; Saeed, A.K.; Mehdi, A.H. Role of Green Tea in Reducing Epidermal Thickness upon Ultraviolet Light-B Injury in BALB/c Mice. Adv. Biol. 2015, 2015, 890632. [Google Scholar] [CrossRef]
- Williams, J.D.; Bermudez, Y.; Park, S.L.; Stratton, S.P.; Uchida, K.; Hurst, C.A.; Wondrak, G.T. Malondialdehyde-derived epitopes in human skin result from acute exposure to solar UV and occur in nonmelanoma skin cancer tissue. J. Photochem. Photobiol. B 2014, 132, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Manosroi, A.; Chankhampan, C.; Pattamapun, K.; Manosroi, W.; Manosroi, J. Antioxidant and gelatinolytic activities of papain from papaya latex and bromelain from pineapple fruits. Chiang Mai J. Sci. 2014, 41, 635–648. [Google Scholar]
- Anggakusuma, Y.; Hwang, J.K. Effects of Macelignan isolated from Myristica fragrans Houtt. on UVB-induced matrix metalloproteinase-9 and cyclooxygenase-2 in HaCaT cells. J. Dermatol. Sci. 2010, 57, 114–122. [Google Scholar] [CrossRef]
- Goldenberg, M.M. Celecoxib, a selective cyclooxygenase-2 inhibitor for the treatment of rheumatoid arthritis and osteoarthritis. Clin. Ther. 1999, 21, 1497–1513. [Google Scholar] [CrossRef]
- Bhui, K.; Prasad, S.; George, J.; Shukla, Y. Bromelain inhibits COX-2 expression by blocking the activation of MAPK regulated NF-Kappa B against skin tumor-initiation triggering mitochondrial death pathway. Cancer Lett. 2009, 282, 167–176. [Google Scholar] [CrossRef]
- Agoro, R.; Piotet-Morin, J.; Palomo, J.; Michaudel, C.; Vigne, S.; Maillet, I.; Chenuet, P.; Guillou, N.; Le Bérichel, J.; Kisielow, M.; et al. IL-1R1-MyD88 axis elicits papain-induced lung inflammation. Eur. J. Immunol. 2016, 46, 2531–2541. [Google Scholar] [CrossRef]
- Shreedhar, V.; Giese, T.; Sung, V.W.; Ullrich, S.E. A cytokine cascade including prostaglandin E2, IL-4, and IL-10 is responsible for UV-induced systemic immune suppression. J. Immunol. 1998, 160, 3783–3789. [Google Scholar] [PubMed]
- Strozyk, E.; Kulms, D. The Role of AKT/MTOR Pathway in stress response to UV-irradiation: Implication in skin carcinogenesis by regulation of apoptosis, Autophagy and Senescence. Int. J. Mol. Sci. 2013, 14, 15260–15285. [Google Scholar] [CrossRef] [PubMed]
- Mir-Palomo, S.; Nácher, A.; Díez-Sales, O.; Vila-Busó, O.; Caddeo, C.; Manca, M.L.; Manconi, M.; Fadda, A.M.; Saurí, A.R. Inhibition of skin inflammation by baicalin ultradeformable vesicles. Int. J. Pharm. 2016, 511, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Manconi, M.; Manca, M.L.; Caddeo, C.; Valenti, D.; Cencetti, C.; Diez-Sales, O.; Nacher, A.; Mir-Palomo, S.; Terencio, M.C.; Demurtas, D.; et al. Nanodesign of new self-assembling core-shell gellan-transfersomes loading baicalin and in vivo evaluation of repair response in skin. Nanomedicine 2018, 14, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Sahu, K.; Kaurav, M.; Pandey, R.S. Protease loaded permeation enhancer liposomes for treatment of skin fibrosis arisen from second degree burn. Biomed. Pharmacother. 2017, 94, 747–757. [Google Scholar] [CrossRef]
- Lemos, F.O.; Imaculada, M.; Villalba, C.; Tagliati, C.A.; Cardoso, V.N.; Salas, C.E.; Lopes, M.T.P. Biodistribution, pharmacokinetics and toxicity of a Vasconcellea cundinamarcensis proteinase fraction with pharmacological activity. Rev. Bras. Farmacogn. 2016, 26, 94–101. [Google Scholar] [CrossRef]
- Teixeira, R.D.; Ribeiro, H.A.; Gomes, M.T.; Lopes, M.T.; Salas, C.E. The proteolytic activities in latex from Carica candamarcensis. Plant. Physiol. Bioch. 2008, 46, 956–961. [Google Scholar] [CrossRef]
- Ivan, A.L.; Campanini, M.Z.; Martinez, R.M.; Ferreira, V.S.; Steffen, V.S.; Vicentini, F.T.; Vilela, F.M.; Martins, F.S.; Zarpelon, A.C.; Cunha, T.M.; et al. Pyrrolidine dithiocarbamate inhibits UVB-induced skin inflammation and oxidative stress in hairless mice and exhibits antioxidant activity in vitro. J. Photochem. Photobiol. B 2014, 138, 124–133. [Google Scholar] [CrossRef]
- Casagrande, R.; Georgetti, S.R.; Verri, W.A.; Dorta, D.J.; dos Santos, A.C.; Fonseca, M.J.V. Protective effect of topical formulations containing quercetin against UVB-induced oxidative stress in Hairless mice. J. Photochem. Photobiol. B 2006, 84, 21–27. [Google Scholar] [CrossRef]
- Avila Acevedo, J.G.; Espinosa González, A.M.; Campos, D.M.D.M.; Benitez Flores, J.C.; Delgado, T.H.; Maya, S.F.; Contreras, J.C.; López, J.L.M.; García Bores, A.M. Photoprotection of Buddleja cordata extract against UVB-induced skin damage in SKH-1 Hairless mice. BMC Complement. Altern. Med. 2014, 14, 281. [Google Scholar] [CrossRef][Green Version]
- Bradford, M.M. Rapid and sensitive method for the quantitation microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 254, 248–254. [Google Scholar] [CrossRef]
- Siqueira, I.R.; Cimarosti, H.; Fochesatto, C.; Salbego, C.; Netto, C.A. Age related susceptibility to oxygen and glucose deprivation damage in rat hippocampal slices. Brain Res. 2004, 1025, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Madesh, M.; Balasubramanian, K. Microtiter plate assay for superoxide dismutase using MTT reduction by superoxide. Indian J. Biochem. Biophys. 1997, 34, 535–539. [Google Scholar] [PubMed]
- Shangari, N.; O’Brien, P.J. Catalase activity assays. Curr. Protoc. Toxicol. 2006, 27, 1–16. [Google Scholar]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [PubMed]
- Hissin, P.J.; Hilf, R. A Fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef]
- Lapenna, D.; Ciofani, G.; Pierdomenico, S.D.; Giamberardino, M.A.; Cuccurullo, F. Reaction conditions affecting the relationship between thiobarbituric acid reactivity and lipid peroxides in human plasma. Free Radic. Biol. Med. 2001, 31, 331–335. [Google Scholar] [CrossRef]
- Barcelos, L.S.; Duplaa, C.; Kränkel, N.; Graiani, G.; Invernici, G.; Katare, R.; Siragusa, M.; Meloni, M.; Campesi, I.; Monica, M.; et al. Human CD133 + progenitor cells promote the healing of diabetic ischemic ulcers by paracrine stimulation of angiogenesis and activation of wnt signaling. Circ. Res. 2010, 104, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Kupai, K.; Szucs, G.; Cseh, S.; Hajdu, I.; Csonka, C.; Csont, T.; Ferdinandy, P. Methods matrix metalloproteinase activity assays: Importance of zymography. J. Pharmacol. Toxicol. Methods 2010, 61, 205–209. [Google Scholar] [CrossRef]
- Campos, P.P.; Bakhle, Y.S.; Andrade, S.P. Mechanisms of wound healing responses in lupus-prone new zealand white mouse strain. Wound Repair Regen. 2008, 16, 416–424. [Google Scholar] [CrossRef]
- Gomori, G. Aldehyde-Fuchsin: A new stain for elastic tissue. Am. J. Clin. Pathol. 1950, 20, 665–666. [Google Scholar] [PubMed]
- Ferreira Junior, M.; Batista, S.A.; Vidigal, P.V.T.; Cordeiro, A.A.C.; Oliveira, F.M.S.; Prata, L.O.; Diniz, A.E.T.; Barral, C.M.; Barbuto, R.C.; Gomes, A.D.; et al. Infection with CagA-positive helicobacter pylori strain containing three epiya c phosphorylation sites is associated with more severe gastric lesions in experimentally infected Mongolian gerbils (Meriones unguiculatus). Eur. J. Histochem. 2015, 59, 137–146. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Da Silva, I.L.; Veloso, E.S.; Gonçalves, I.N.N.; Braga, A.D.; Lopes, M.T.P.; Cassali, G.D.; Quintanilla, M.; Simões, R.T.; Ferreira, E. Qa-2 Expression levels is related with tumor-infiltrating lymphocytes profile during solid ehrlich tumor development. Biomed. Pharmacother. 2017, 92, 750–756. [Google Scholar] [CrossRef] [PubMed][Green Version]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, K.M.; Silva, A.C.A.e.; Veloso, E.S.; Ferreira, Ê.; Barcelos, L.S.; Caliari, M.V.; Salas, C.E.; Lopes, M.T.P. P1G10, the Proteolytic Fraction from Vasconcellea cundinamarcensis, Stimulates Tissue Repair after Acute Exposure to Ultraviolet B Radiation. Int. J. Mol. Sci. 2019, 20, 4373. https://doi.org/10.3390/ijms20184373
Freitas KM, Silva ACAe, Veloso ES, Ferreira Ê, Barcelos LS, Caliari MV, Salas CE, Lopes MTP. P1G10, the Proteolytic Fraction from Vasconcellea cundinamarcensis, Stimulates Tissue Repair after Acute Exposure to Ultraviolet B Radiation. International Journal of Molecular Sciences. 2019; 20(18):4373. https://doi.org/10.3390/ijms20184373
Chicago/Turabian StyleFreitas, Kátia M., Ana C. Araújo e Silva, Emerson S. Veloso, Ênio Ferreira, Lucíola S. Barcelos, Marcelo V. Caliari, Carlos E. Salas, and Miriam T. P. Lopes. 2019. "P1G10, the Proteolytic Fraction from Vasconcellea cundinamarcensis, Stimulates Tissue Repair after Acute Exposure to Ultraviolet B Radiation" International Journal of Molecular Sciences 20, no. 18: 4373. https://doi.org/10.3390/ijms20184373
APA StyleFreitas, K. M., Silva, A. C. A. e., Veloso, E. S., Ferreira, Ê., Barcelos, L. S., Caliari, M. V., Salas, C. E., & Lopes, M. T. P. (2019). P1G10, the Proteolytic Fraction from Vasconcellea cundinamarcensis, Stimulates Tissue Repair after Acute Exposure to Ultraviolet B Radiation. International Journal of Molecular Sciences, 20(18), 4373. https://doi.org/10.3390/ijms20184373