Investigating the Role of VDR and Megalin in Semi-Selectivity of Side-Chain Modified 19-nor Analogs of Vitamin D

 ,
, 
Abstract
:1. Introduction
2. Results
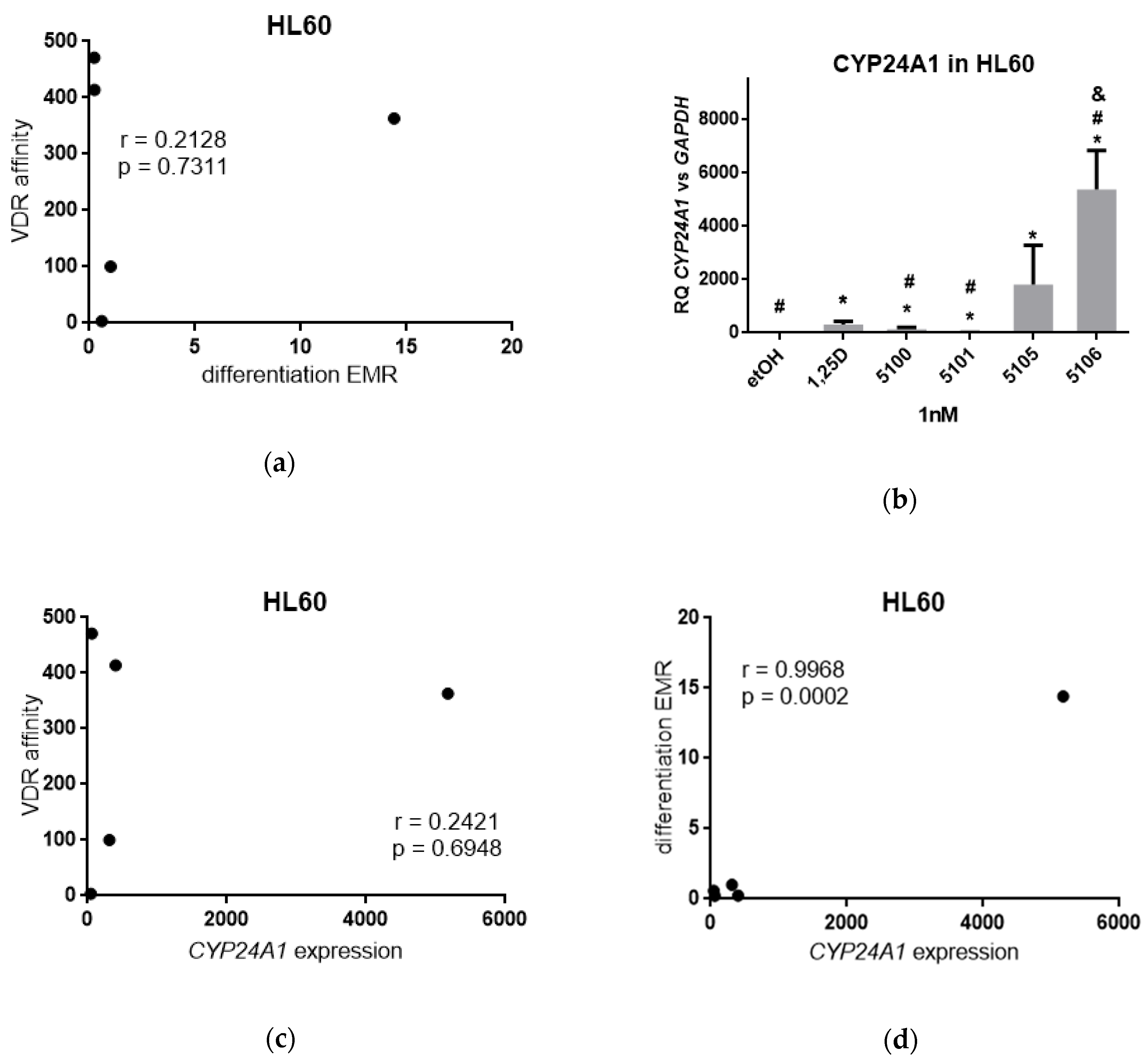
2.1. Binding of Analogs to VDR
2.2. Biological Actions of Analogs in Acute Myeloid Leukemia Cells
2.3. Biological Actions of Analogs in Intestine Cells
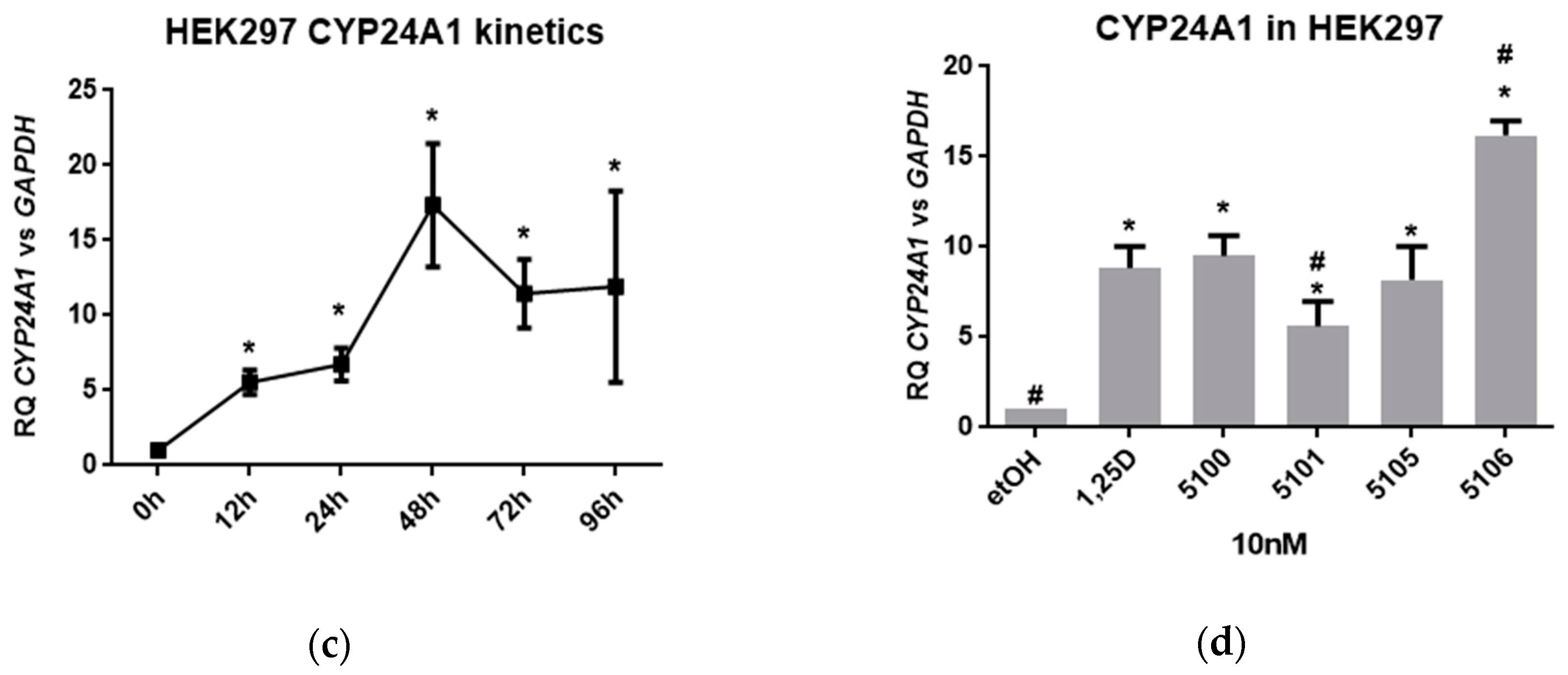
2.4. Biological Actions of Analogs in Bone and Kidney Cells
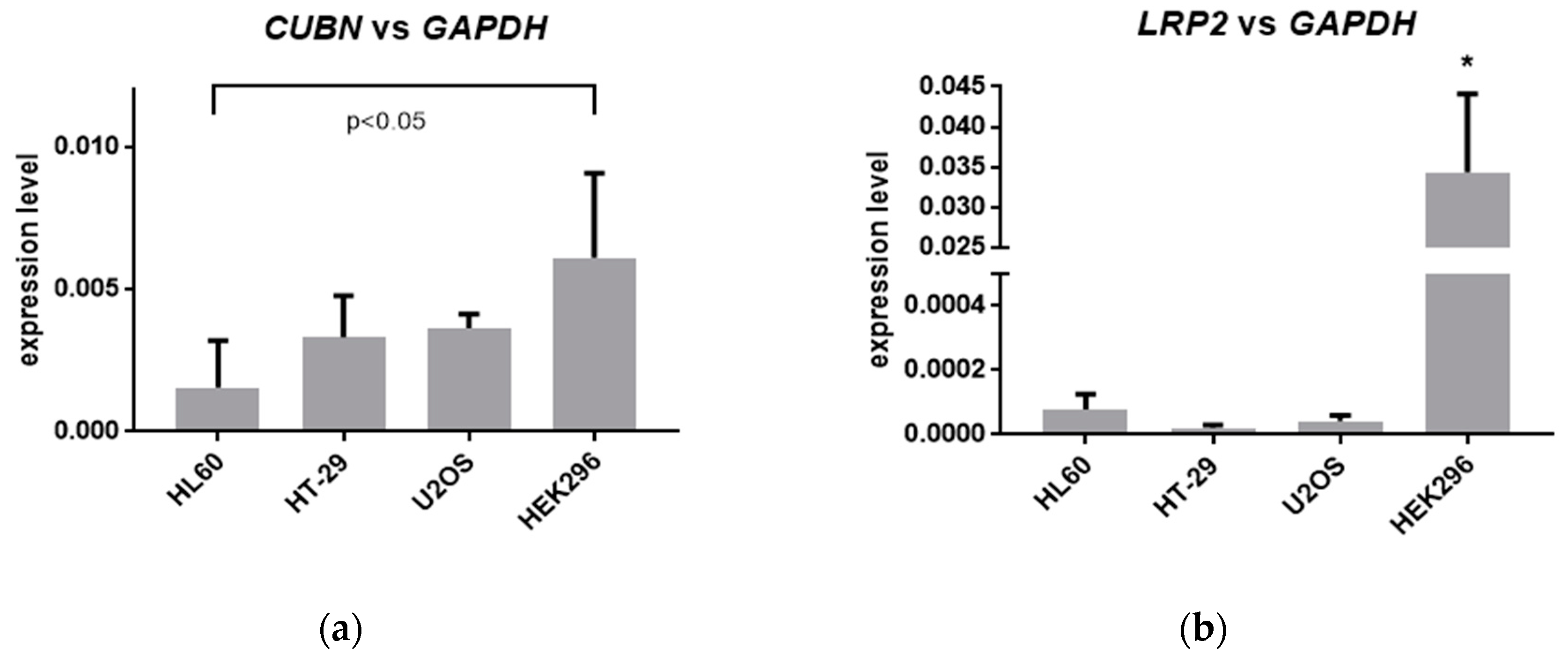
2.5. Expression of Megalin in Tested Cells
2.6. Knock-Out of Megalin in HEK297 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Chemicals and Antibodies
4.3. Determination of Cell Differentiation
4.4. cDNA Synthesis and RealTime PCR
4.5. Human VDR Binding Assay
4.6. Knock-Out of LRP2 Gene Using CRISPR/Cas9 Method
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 1,25D3 | 1,25-dihydroxyvitamin D3 |
| AhR | aryl hydrocarbon receptor |
| C/EBPβ | CCAAT-enhancer-binding protein β |
| CYP24A1 | 24-hydroxylase of 1,25D3 |
| DBP | vitamin D-binding protein |
| EMR | effective molar ratio |
| FP | fluorescence polarization |
| MARRS | membrane associated rapid response steroid-binding protein |
| RBA | relative binding affinity |
| TRPV6 | vanilloid transient receptor potential 6 |
| VDR | vitamin D receptor |
References
- Feldman, D.; Krishnan, A.; Swami, S.; Giovannucci, E.; Feldman, B. The role of vitamin D in reducing cancer risk and progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Holick, M. Vitamin D and bone health. J. Nutr. 1996, 126, 1159S–1164S. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, S.; Chodynski, M.; Corcoran, A.; Marcinkowska, E.; Brown, G.; Kutner, A. Double point modified analogs of vitamin D as potent activators of vitamin D receptor. Curr. Pharm. Des. 2015, 21, 1741–1763. [Google Scholar] [CrossRef] [PubMed]
- Pike, J.; Meyer, M. Fundamentals of vitamin D hormone-regulated gene expression. J. Steroid Biochem. Mol. Biol. 2014, 144, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, S.; Dunlop, T.; Sinkkonen, L.; Frank, C.; Carlberg, C. Spatio-temporal activation of chromatin on the human CYP24 gene promoter in the presence of 1alpha,25-dihydroxyvitamin D3. J. Mol. Biol. 2005, 350, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Urushino, N.; Yasuda, K.; Ikushiro, S.; Kamakura, M.; Ohta, M.; Sakaki, T. Metabolism of 1α,25-dihydroxyvitamin D2 by human CYP24A1. Biochem. Biophys. Res. Commun 2009, 384, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Gocek, E.; Marchwicka, A.; Baurska, H.; Chrobak, A.; Marcinkowska, E. Opposite regulation of vitamin D receptor by ATRA in AML cells susceptible and resistant to vitamin D-induced differentiation. J. Steroid Biochem. Mol. Biol. 2012, 132, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Riley, E.; Meyer, M.; Benkusky, N.; Plum, L.; DeLuca, H.; Pike, J. 1,25-Dihydroxyvitamin D3 Controls a Cohort of Vitamin D Receptor Target Genes in the Proximal Intestine That Is Enriched for Calcium-regulating Components. J. Biol. Chem. 2015, 290, 18199–18215. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.; Tan, S.; Tenen, D.; Nicholson-Weller, A.; Seed, B. Monocyte antigen CD14 is a phospholipid anchored membrane protein. Blood 1989, 73, 284–289. [Google Scholar] [Green Version]
- Ryynänen, J.; Seuter, S.; Campbell, M.; Carlberg, C. Gene regulatory scenarios of primary 1,25-dihydroxyvitamin D3 target genes in a human myeloid leukemia cell line. Cancers 2013, 5, 1221–1241. [Google Scholar] [CrossRef]
- Marcinkowska, E.; Garay, E.; Gocek, E.; Chrobak, A.; Wang, X.; Studzinski, G. Regulation of C/EBPbeta isoforms by MAPK pathways in HL60 cells induced to differentiate by 1,25-dihydroxyvitamin D3. Exp. Cell Res. 2006, 312, 2054–2065. [Google Scholar] [CrossRef] [PubMed]
- Marcinkowska, E. A run for a membrane vitamin D receptor. Biol Signals Recept 2001, 10, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Nemere, I. The 1,25D3-MARRS protein: Contribution to steroid stimulated calcium uptake in chicks and rats. Steroids 2005, 70, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Mizwicki, M.; Keidel, D.; Bula, C.; Bishop, J.; Zanello, L.; Wurtz, J.; Moras, D.; Norman, A. Identification of an alternative ligand-binding pocket in the nuclear vitamin D receptor and its functional importance in 1alpha,25(OH)2-vitamin D3 signaling. Proc. Natl. Acad. Sci. USA 2004, 101, 12876–12881. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, T. The role of plasma-binding proteins in the cellular uptake of lipophilic vitamins and steroids. Horm. Metab. Res. 2006, 38, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.; Birn, H. Megalin and cubilin: Synergistic endocytic receptors in renal proximal tubule. Am. J. Physiol. Ren. Physiol. 2001, 280, F562–F573. [Google Scholar] [CrossRef]
- Rowling, M.; Kemmis, C.; Taffany, D.; Welsh, J. Megalin-mediated endocytosis of vitamin D binding protein correlates with 25-hydroxycholecalciferol actions in human mammary cells. J. Nutr. 2006, 136, 2754–2759. [Google Scholar] [CrossRef]
- Chun, R.; Peercy, B.; Orwoll, E.; Nielson, C.; Adams, J.; Hewison, M. Vitamin D and DBP: The free hormone hypothesis revisited. J. Steroid Biochem. Mol. Biol. 2014, 144, 132–137. [Google Scholar] [CrossRef]
- Chapron, B.; Chapron, A.; Phillips, B.; Okoli, M.; Shen, D.; Kelly, E.; Himmelfarb, J.; Thummel, K. Reevaluating the role of megalin in renal vitamin D homeostasis using a human cell-derived microphysiological system. ALTEX 2018, 35, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Cashman, K.; Kinsella, M.; McNulty, B.; Walton, J.; Gibney, M.; Flynn, A.; Kiely, M. Dietary vitamin D2--a potentially underestimated contributor to vitamin D nutritional status of adults? Br. J. Nutr. 2014, 112, 193–202. [Google Scholar] [CrossRef]
- Corcoran, A.; Bermudez, M.; Seoane, S.; Perez-Fernandez, R.; Krupa, M.; Pietraszek, A.; Chodyński, M.; Kutner, A.; Brown, G.; Marcinkowska, E. Biological evaluation of new vitamin D2 analogues. J. Steroid Biochem. Mol. Biol. 2016, 164, 66–71. [Google Scholar] [CrossRef]
- Trynda, J.; Turlej, E.; Milczarek, M.; Pietraszek, A.; Chodyński, M.; Kutner, A.; Wietrzyk, J. Antiproliferative Activity and in Vivo Toxicity of Double-Point Modified Analogs of 1,25-Dihydroxyergocalciferol. Int. J. Mol. Sci. 2015, 16, 24873–24894. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Hetherington, C.; Zhang, D. CCAAT/enhancer-binding protein activates the CD14 promoter and mediates transforming growth factor beta signaling in monocyte development. J. Biol. Chem. 1999, 274, 23242–23248. [Google Scholar] [CrossRef]
- Baurska, H.; Kłopot, A.; Kiełbiński, M.; Chrobak, A.; Wijas, E.; Kutner, A.; Marcinkowska, E. Structure-function analysis of vitamin D2 analogs as potential inducers of leukemia differentiation and inhibitors of prostate cancer proliferation. J. Steroid Biochem. Mol. Biol. 2011, 126, 46–54. [Google Scholar] [CrossRef]
- Nijenhuis, T.; Hoenderop, J.; Nilius, B.; Bindels, R. (Patho)physiological implications of the novel epithelial Ca2+ channels TRPV5 and TRPV6. Pflug. Arch. 2003, 446, 401–409. [Google Scholar] [CrossRef]
- Meyer, M.; Watanuki, M.; Kim, S.; Shevde, N.; Pike, J. The human transient receptor potential vanilloid type 6 distal promoter contains multiple vitamin D receptor binding sites that mediate activation by 1,25-dihydroxyvitamin D3 in intestinal cells. Mol. Endocrinol. 2006, 20, 1447–1461. [Google Scholar] [CrossRef]
- Atkins, G.; Anderson, P.; Findlay, D.; Welldon, K.; Vincent, C.; Zannettino, A.; O’Loughlin, P.; Morris, H. Metabolism of vitamin D3 in human osteoblasts: Evidence for autocrine and paracrine activities of 1 alpha,25-dihydroxyvitamin D3. Bone 2007, 40, 1517–1528. [Google Scholar] [CrossRef]
- Dusso, A.; Brown, A.; Slatopolsky, E. Vitamin D. Am. J. Physiol. Renal. Physiol. 2005, 289, F8–F28. [Google Scholar] [CrossRef]
- Bikle, D. Vitamin D Metabolism, Mechanism of Action, and Clinical Applications. Chem. Biol. 2014, 21, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.; Slatopolsky, E. Vitamin D analogs: Therapeutic applications and mechanisms for selectivity. Mol. Asp. Med. 2008, 29, 433–452. [Google Scholar] [CrossRef]
- Bouillion, R.; Okamura, W.; Norman, A. Structure-function relationships in the vitamin D endocrine system. Endocr. Rev. 1995, 16, 200–216. [Google Scholar]
- Slominski, A.; Kim, T.; Takeda, Y.; Janjetovic, Z.; Brozyna, A.; Skobowiat, C.; Wang, J.; Postlethwaite, A.; Li, W.; Tuckey, R.; et al. RORα and ROR γ are expressed in human skin and serve as receptors for endogenously produced noncalcemic 20-hydroxy- and 20,23-dihydroxyvitamin D. FASEB J. 2014, 28, 2775–2789. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.; Janjetovic, Z.; Brożyna, A.; Żmijewski, M.; Xu, H.; Sutter, T.; Tuckey, R.; Jetten, A.; Crossman, D. Differential and Overlapping Effects of 20,23(OH)2D3 and 1,25(OH)2D3 on Gene Expression in Human Epidermal Keratinocytes: Identification of AhR as an Alternative Receptor for 20,23(OH)2D3. Int. J. Mol. Sci. 2018, 19, 3072. [Google Scholar] [CrossRef]
- Perlman, K.; Prahl, J.; Smith, C.; Kutner, A.; DeLuca, H. 6,27-Dihomo-1 alpha-hydroxy- and 26,27-dihomo-24-epi-1 alpha,25-dihydroxyvitamin D2 analogs that differ markedly in biological activity in vivo. J. Biol. Chem. 1994, 269, 24014–24019. [Google Scholar]
- Liu, W.; Xie, X.; Ma, X.; Li, J.; Chen, J.; Liu, Y. DSDecode: A Web-Based Tool for Decoding of Sequencing Chromatograms for Genotyping of Targeted Mutations. Mol. Plant. 2015, 8, 1431–1433. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1,25D3 | PRI-5100 | PRI-5101 | PRI-5105 | PRI-5106 | |
|---|---|---|---|---|---|
| IC50 (M) | 2.320 × 10−9 | 5.599 × 10−10 | 4.921 × 10−10 | 6.336 × 10−8 | 6.389 × 10−10 |
| RBA a | 100 | 414 | 471 | 3.66 | 363 |
| 1,25D3 | PRI-5100 | PRI-5101 | PRI-5105 | PRI-5106 | |
|---|---|---|---|---|---|
| EC50 (M) | 2.745 × 10−10 | 1.128 × 10−9 | 1.179 × 10−9 | 4.763 × 10−10 | 1.901 × 10−11 |
| EMR | 1 | 0.24 | 0.23 | 0.58 | 14.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berkowska, K.; Corcoran, A.; Grudzień, M.; Jakuszak, A.; Chodyński, M.; Kutner, A.; Marcinkowska, E. Investigating the Role of VDR and Megalin in Semi-Selectivity of Side-Chain Modified 19-nor Analogs of Vitamin D. Int. J. Mol. Sci. 2019, 20, 4183. https://doi.org/10.3390/ijms20174183
Berkowska K, Corcoran A, Grudzień M, Jakuszak A, Chodyński M, Kutner A, Marcinkowska E. Investigating the Role of VDR and Megalin in Semi-Selectivity of Side-Chain Modified 19-nor Analogs of Vitamin D. International Journal of Molecular Sciences. 2019; 20(17):4183. https://doi.org/10.3390/ijms20174183
Chicago/Turabian StyleBerkowska, Klaudia, Aoife Corcoran, Małgorzata Grudzień, Agnieszka Jakuszak, Michał Chodyński, Andrzej Kutner, and Ewa Marcinkowska. 2019. "Investigating the Role of VDR and Megalin in Semi-Selectivity of Side-Chain Modified 19-nor Analogs of Vitamin D" International Journal of Molecular Sciences 20, no. 17: 4183. https://doi.org/10.3390/ijms20174183