Aggregation States of Aβ1–40, Aβ1–42 and Aβp3–42 Amyloid Beta Peptides: A SANS Study


 ,
,  ,
,  , ,
, ,
Abstract
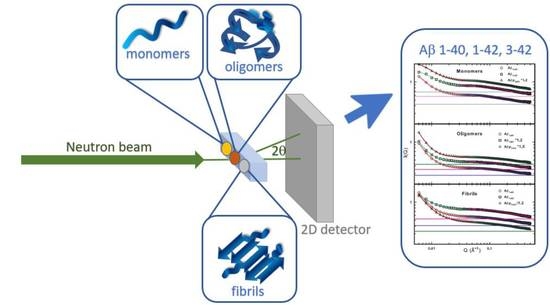
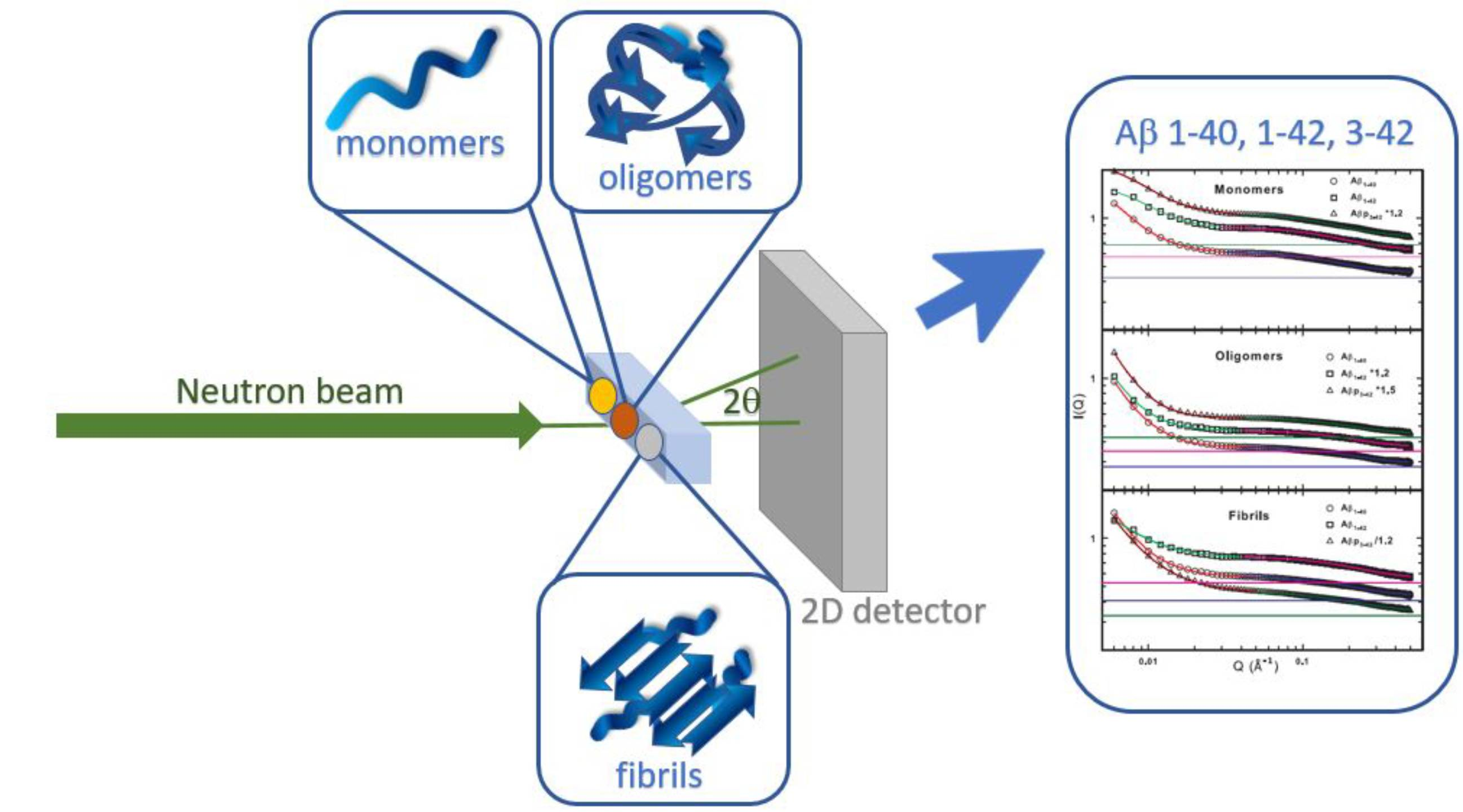
:
1. Introduction
2. Results and Discussion
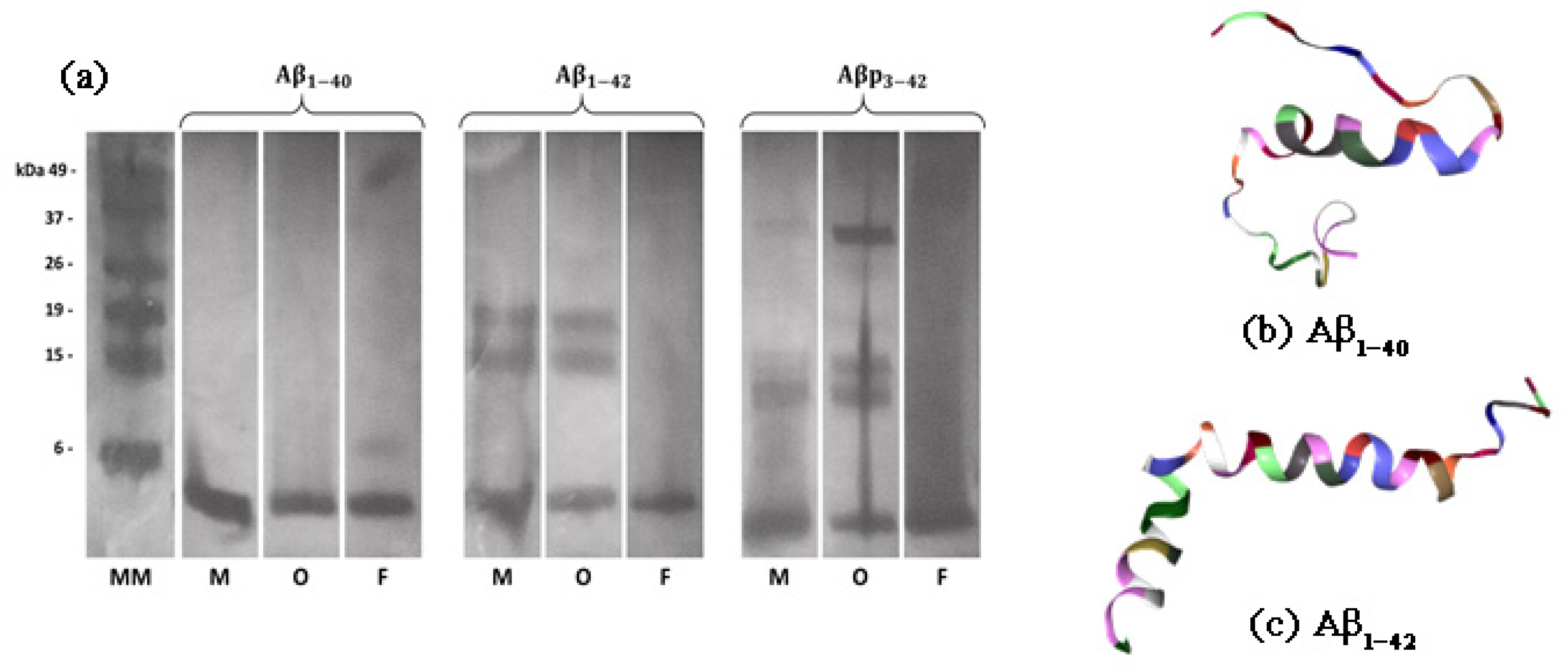
2.1. Formation of Synthetic A, A and A Aggregates
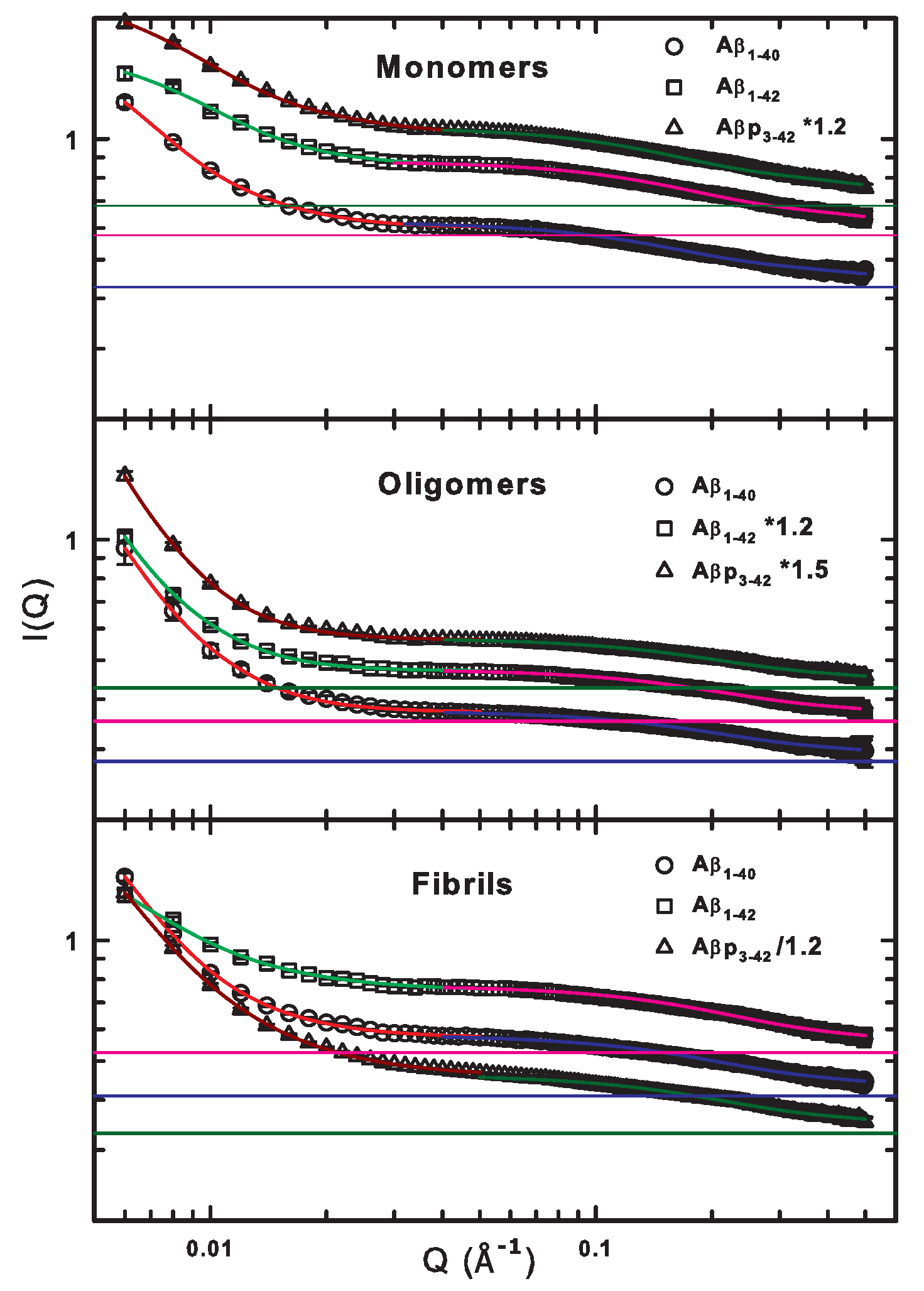
2.2. SANS Results and Discussions of the G, and d Parameters
3. Materials and Methods
3.1. Preparation of A Aggregates and Their Analysis by SDS-PAGE
3.2. Preparation of A for SANS2D
3.3. SDS-PAGE Analysis
3.4. The Small Angle Neutron Scattering Technique
3.5. SANS Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| SANS | Small angle neutron scattering |
| SDS-PAGE | Sodium dodecyl sulfate–poly-acrylamide gel electrophoresis |
References
- Serrano-Pozo, A.; Frosch, M.; Masliah, E.; Hyman, B. Neuropathological alterations in Alzheimer disease. Cold Spring Harb. Perspect. Med. 2011, 1, a006189. [Google Scholar] [CrossRef]
- Tiraboschi, P.; Hansen, L.; Thal, L.; Corey-Bloom, J. The importance of neuritic plaques and tangles to the development and evolution of AD. Neurology 2004, 62, 1984–1989. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef] [PubMed]
- Thal, D.; Rüb, U.; Orantes, M.; Braak, H. Phases of Aβ-deposition in the human brain and its relevance for the development of AD. Neurology 2002, 58, 1791–1800. [Google Scholar] [CrossRef] [PubMed]
- Hyman, B.; Marzloff, K.; Arriagada, P. The lack of accumulation of senile plaques or amyloid burden in alzheimer’s disease suggests a dynamic balance between amyloid deposition and resolution. J. Neuropathol. Exp. Neurol. 1993, 52, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Pozo, A.; Mielke, M.; Muzitansky, A.; Gómez-Isla, T.; Growdon, J.; Bacskai, B.; Betensky, R.; Frosch, M.; Hyman, B. Stable size distribution of amyloid plaques over the course of alzheimer disease. J. Neuropathol. Exp. Neurol. 2012, 71, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Priller, C.; Bauer, T.; Mitteregger, G.; Krebs, B.; Kretzschmar, H.; Herms, J. Synapse formation and function is modulated by the amyloid precursor protein. J. Neurosci. 2006, 26, 7212–7221. [Google Scholar] [CrossRef] [PubMed]
- Iwatsubo, T.; Saido, T.; Mann, D.; Lee, V.Y.; Trojanowski, J. Full-length amyloid-β(1-42(43)) and amino-terminally modified and truncated amyloid-β42(43) deposit in diffuse plaques. Am. J. Pathol. 1996, 149, 1823–1830. [Google Scholar] [PubMed]
- Saido, T.; Iwatsubo, T.; Mann, D.; Shimada, H.; Ihara, Y.; Kawashima, S. Dominant and differential deposition of distinct β-amyloid peptide species, AβN3(pE), in senile plaques. Neuron 1995, 14, 457–466. [Google Scholar] [CrossRef]
- Bayer, T.; Wirths, O. Focusing the amyloid cascade hypothesis on N-truncated Abeta peptides as drug targets against Alzheimer’s disease. Acta Neuropathol. 2014, 127, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Watzky, M.; Finke, R. Protein aggregation kinetics, mechanism, and curve-fitting: A review of the literature. Biochim. Biophys. Acta-Proteins Proteom. 2009, 1794, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Zhang-Haagen, B.; Biehl, R.; Nagel-Steger, L.; Radulescu, A.; Richter, D.; Willbold, D. Monomeric Amyloid beta peptide in hexafluoroisopropanol detected by small angle neutron scattering. PLoS ONE 2016, 11, e0150267. [Google Scholar] [CrossRef] [PubMed]
- Festa, G.; Sancesario, G.; Corsaro, C.; Longo, S.; Mallamace, D.; Fazio, E.; Arcidiacono, L.; Sakai, V.; Senesi, R.; Sancesario, G.; et al. SANS study of Amyloid β1-40: Unfolded monomers in DMSO, multidimensional aggregates in water medium. Phys. A 2019, 517, 385–391. [Google Scholar] [CrossRef]
- Sancesario, G.; Cencioni, M.; Esposito, Z.; Borsellino, G.; Nuccetelli, M.; Martorana, A.; Battistini, L.; Sorge, R.; Spalletta, G.; Ferrazzoli, D.; et al. The load of amyloid-β oligomers is decreased in the cerebrospinal fluid of Alzheimer’s disease patients. J. Alzheimer’s Dis. 2012, 31, 865–878. [Google Scholar] [CrossRef]
- Bitan, G.; Kirkitadze, M.D.; Lomakin, A.; Vollers, S.S.; Benedek, G.B.; Teplow, D.B. Amyloid β-protein (Aβ) assembly: Aβ40 and Aβ42 oligomerize through distinct pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 330–335. [Google Scholar] [CrossRef]
- Hammouda, B. Analysis of the Beaucage model. J. Appl. Crystallogr. 2010, 43, 1474–1478. [Google Scholar] [CrossRef]
- Beaucage, G. Approximations Leading to a Unified Exponential/Power-Law Approach to Small-Angle Scattering. J. Appl. Crystallogr. 1995, 28, 717–728. [Google Scholar] [CrossRef]
- Magazu, S.; Maisano, G.; Mallamace, F.; Micali, N. Growth of fractal aggregates in water solutions of macromolecules by light scattering. Phys. Rev. A 1989, 39, 4195–4200. [Google Scholar] [CrossRef]
- Sengupta, U.; Nilson, A.; Kayed, R. The Role of Amyloid-β Oligomers in Toxicity, Propagation, and Immunotherapy. EBioMedicine 2016, 6, 42–49. [Google Scholar] [CrossRef]
- Mallamace, D.; Fazio, E.; Mallamace, F.; Corsaro, C. The role of hydrogen bonding in the folding/unfolding process of hydrated lysozyme: A review of recent NMR and FTIR results. Int. J. Mol. Sci. 2018, 19, 3825. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Freire, S.; Al-Soufi, W. Critical aggregation concentration for the formation of early Amyloid-β (1-42) oligomers. Sci. Rep. 2018, 8, 1783. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, K.; Manelli, A.; Blaine Stine, W., Jr.; Baker, L.; Krafft, G.; Ladu, M. Oligomeric and fibrillar species of amyloid-β peptides differentially affect neuronal viability. J. Biol. Chem. 2002, 277, 32046–32053. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.; Barlow, A.; Chromy, B.; Edwards, C.; Freed, R.; Liosatos, M.; Morgan, T.; Rozovsky, I.; Trommer, B.; Viola, K.; et al. Diffusible, nonfibrillar ligands derived from Aβ1-42 are potent central nervous system neurotoxins. Proc. Natl. Acad. Sci. USA 1998, 95, 6448–6453. [Google Scholar] [CrossRef] [PubMed]
- Sancesario, G.; Nuccetelli, M.; Cerri, A.; Zegeer, J.; Severini, C.; Ciotti, M.; Pieri, M.; Martorana, A.; Caltagirone, C.; Nisticò, R.; et al. Bromelain degrades Aβ1-42 monomers and soluble aggregates: An in vitro study in cerebrospinal fluid of alzheimer’s disease patients. Curr. Alzheimer Res. 2018, 15, 628–636. [Google Scholar] [CrossRef]
- Heenan, R.; Rogers, S.; Turner, D.; Terry, A.; Treadgold, J.; King, S. Small angle neutron scattering using Sans2d. Neutron News 2011, 22, 19–21. [Google Scholar] [CrossRef]
- Sans2d: Time-of-Flight Small-Angle Neutron Scattering Instrument. 2019. Available online: https://www.isis.stfc.ac.uk/Pages/Sans2d.aspx (accessed on 22 August 2019).
- Wignall, G.; Bates, F. Absolute calibration of small-angle neutron scattering data. J. Appl. Crystallogr. 1987, 20, 28–40. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
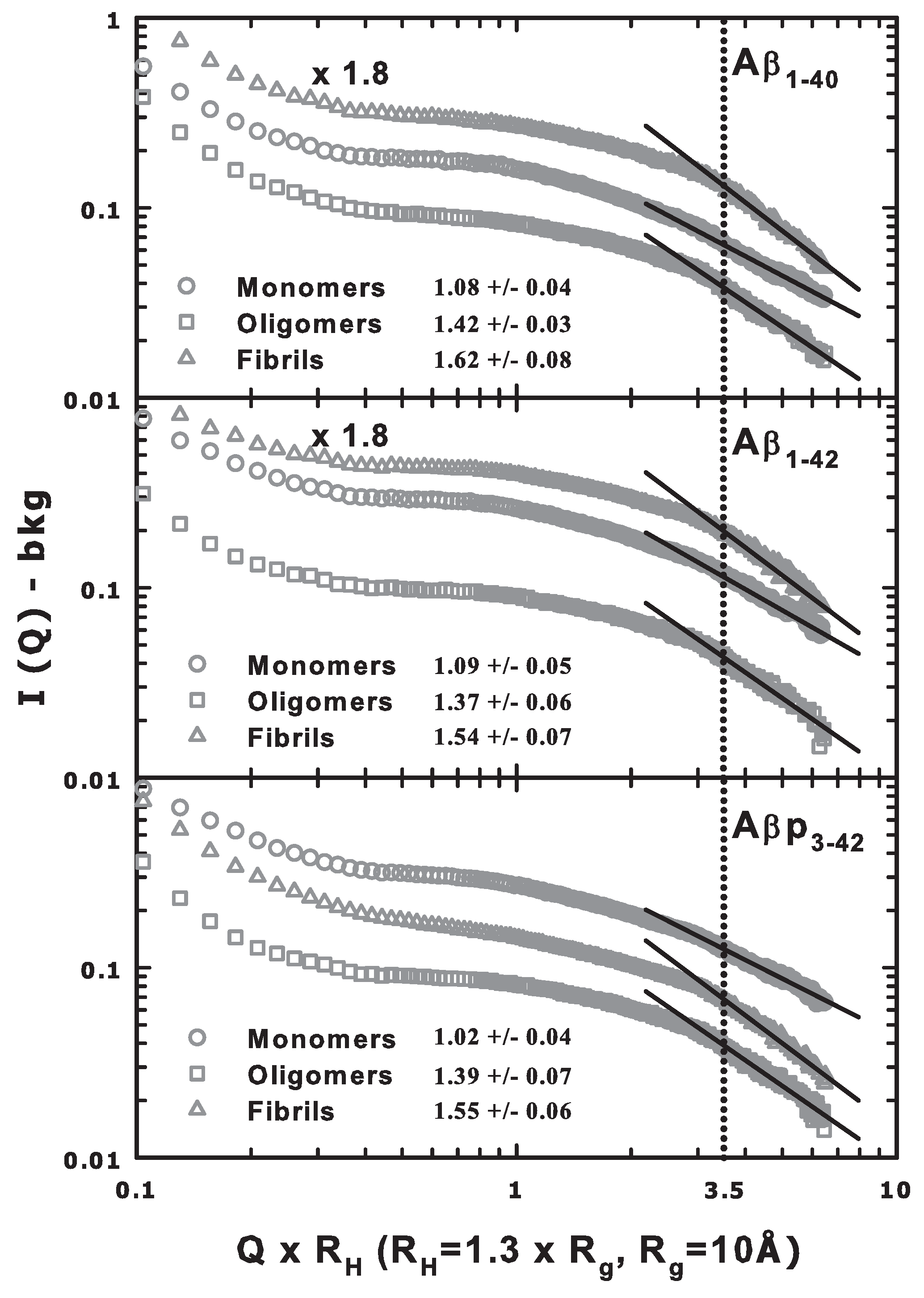{kind=link}
| G (cm) | R (Å) | d | bkg (cm) | ||
|---|---|---|---|---|---|
| low Q | |||||
| Monomers | 1.30 ± 0.07 | 253.3 ± 9.6 | 2.8 ± 0.2 | 0.596 ± 0.006 | |
| Oligomers | 2.50 ± 0.56 | 377.9 ± 40.6 | 3.1 ± 0.1 | 0.371 ± 0.001 | |
| Fibrils | 11.72 ± 1.34 | 705.6 ± 51.7 | 2.4 ± 0.1 | 0.570 ± 0.002 | |
| A | |||||
| high Q | |||||
| Monomers | 0.19 ± 0.01 | 10.9 ± 0.4 | 1.2 ± 0.2 | 0.427 ± 0.007 | |
| Oligomers | 0.09 ± 0.01 | 8.1 ± 1.0 | 1.3 ± 0.5 | 0.278 ± 0.012 | |
| Fibrils | 0.17 ± 0.03 | 8.4 ± 0.9 | 1.3 ± 0.6 | 0.409 ± 0.026 | |
| low Q | |||||
| Monomers | 0.88 ± 0.03 | 174.8 ± 5.7 | 3.1 ± 0.4 | 0.857 ± 0.006 | |
| Oligomers | 2.61 ± 0.72 | 425.9 ± 45.1 | 3.1 ± 0.4 | 0.392 ± 0.002 | |
| Fibrils | 11.49 ± 3.22 | 879.6 ± 85.8 | 2.1 ± 0.1 | 0.753 ± 0.002 | |
| A | |||||
| high Q | |||||
| Monomers | 0.30 ± 0.01 | 10.1 ± 0.4 | 1.1 ± 0.2 | 0.575 ± 0.014 | |
| Oligomers | 0.10 ± 0.02 | 8.1 ± 1.0 | 1.3 ± 0.8 | 0.294 ± 0.019 | |
| Fibrils | 0.24 ± 0.06 | 7.8 ± 1.2 | 1.3 ± 0.8 | 0.525 ± 0.058 | |
| low Q | |||||
| Monomers | 1.15 ± 0.02 | 186.1 ± 3.7 | 2.4 ± 0.1 | 0.855 ± 0.003 | |
| Oligomers | 6.14 ± 1.60 | 541.5 ± 47.0 | 3.1 ± 0.1 | 0.375 ± 0.001 | |
| Fibrils | 11.64 ± 0.69 | 771.2 ± 33.1 | 2.0 ± 0.1 | 0.547 ± 0.002 | |
| Ap | |||||
| high Q | |||||
| Monomers | 0.32 ± 0.01 | 11.3 ± 0.4 | 1.0 ± 0.1 | 0.567 ± 0.011 | |
| Oligomers | 0.09 ± 0.01 | 8.4 ± 0.9 | 1.3 ± 0.6 | 0.285 ± 0.014 | |
| Fibrils | 0.16 ± 0.02 | 8.3 ± 0.8 | 1.3 ± 0.6 | 0.396 ± 0.022 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Festa, G.; Mallamace, F.; Sancesario, G.M.; Corsaro, C.; Mallamace, D.; Fazio, E.; Arcidiacono, L.; Garcia Sakai, V.; Senesi, R.; Preziosi, E.; et al. Aggregation States of Aβ1–40, Aβ1–42 and Aβp3–42 Amyloid Beta Peptides: A SANS Study. Int. J. Mol. Sci. 2019, 20, 4126. https://doi.org/10.3390/ijms20174126
Festa G, Mallamace F, Sancesario GM, Corsaro C, Mallamace D, Fazio E, Arcidiacono L, Garcia Sakai V, Senesi R, Preziosi E, et al. Aggregation States of Aβ1–40, Aβ1–42 and Aβp3–42 Amyloid Beta Peptides: A SANS Study. International Journal of Molecular Sciences. 2019; 20(17):4126. https://doi.org/10.3390/ijms20174126
Chicago/Turabian StyleFesta, Giulia, Francesco Mallamace, Giulia Maria Sancesario, Carmelo Corsaro, Domenico Mallamace, Enza Fazio, Laura Arcidiacono, Victoria Garcia Sakai, Roberto Senesi, Enrico Preziosi, and et al. 2019. "Aggregation States of Aβ1–40, Aβ1–42 and Aβp3–42 Amyloid Beta Peptides: A SANS Study" International Journal of Molecular Sciences 20, no. 17: 4126. https://doi.org/10.3390/ijms20174126