Functional Characterization of a Drought-Responsive Invertase Inhibitor from Maize (Zea mays L.)

Abstract
:1. Introduction
2. Results
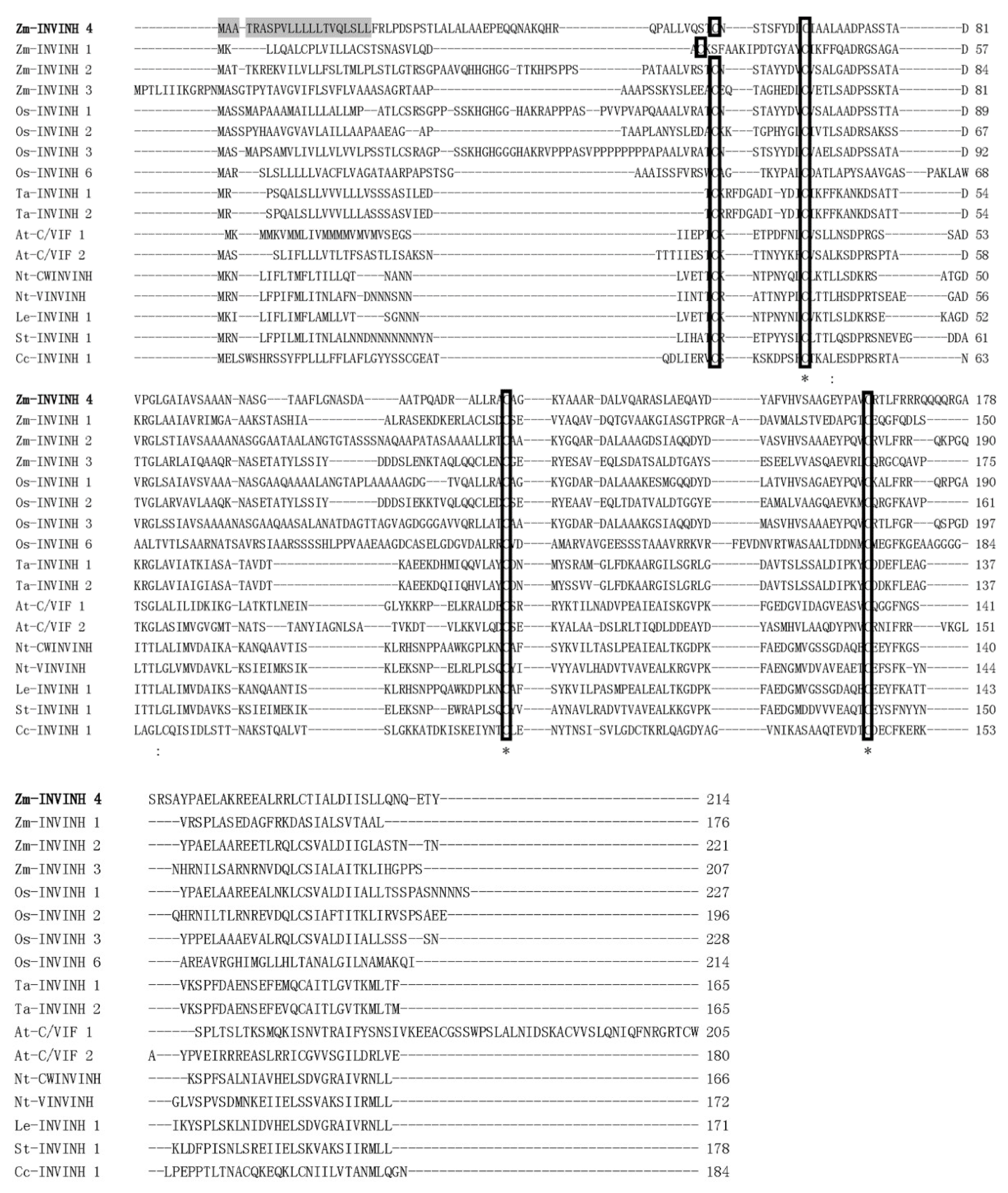
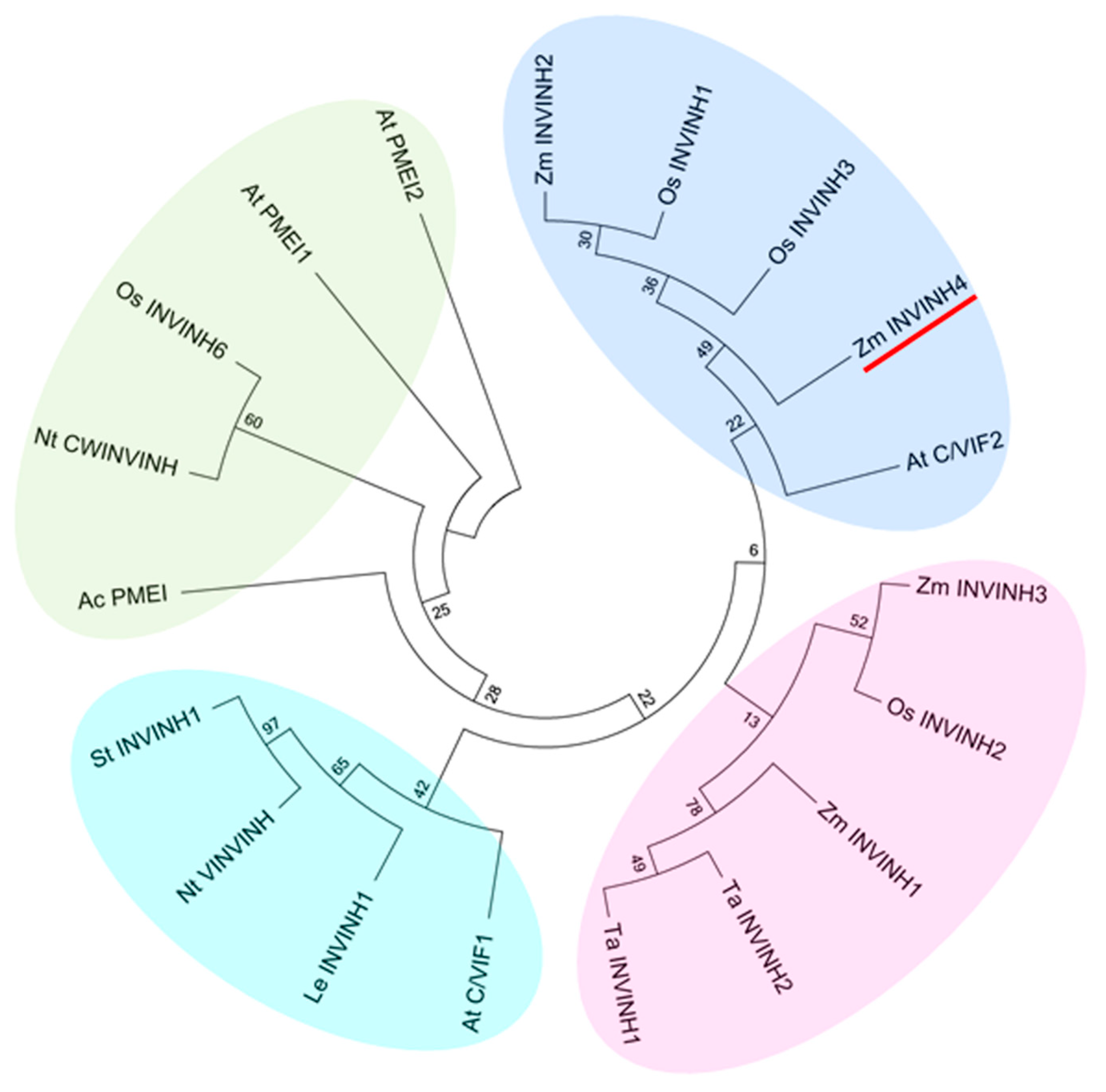
2.1. Identification of the Maize Invertase Inhibitors
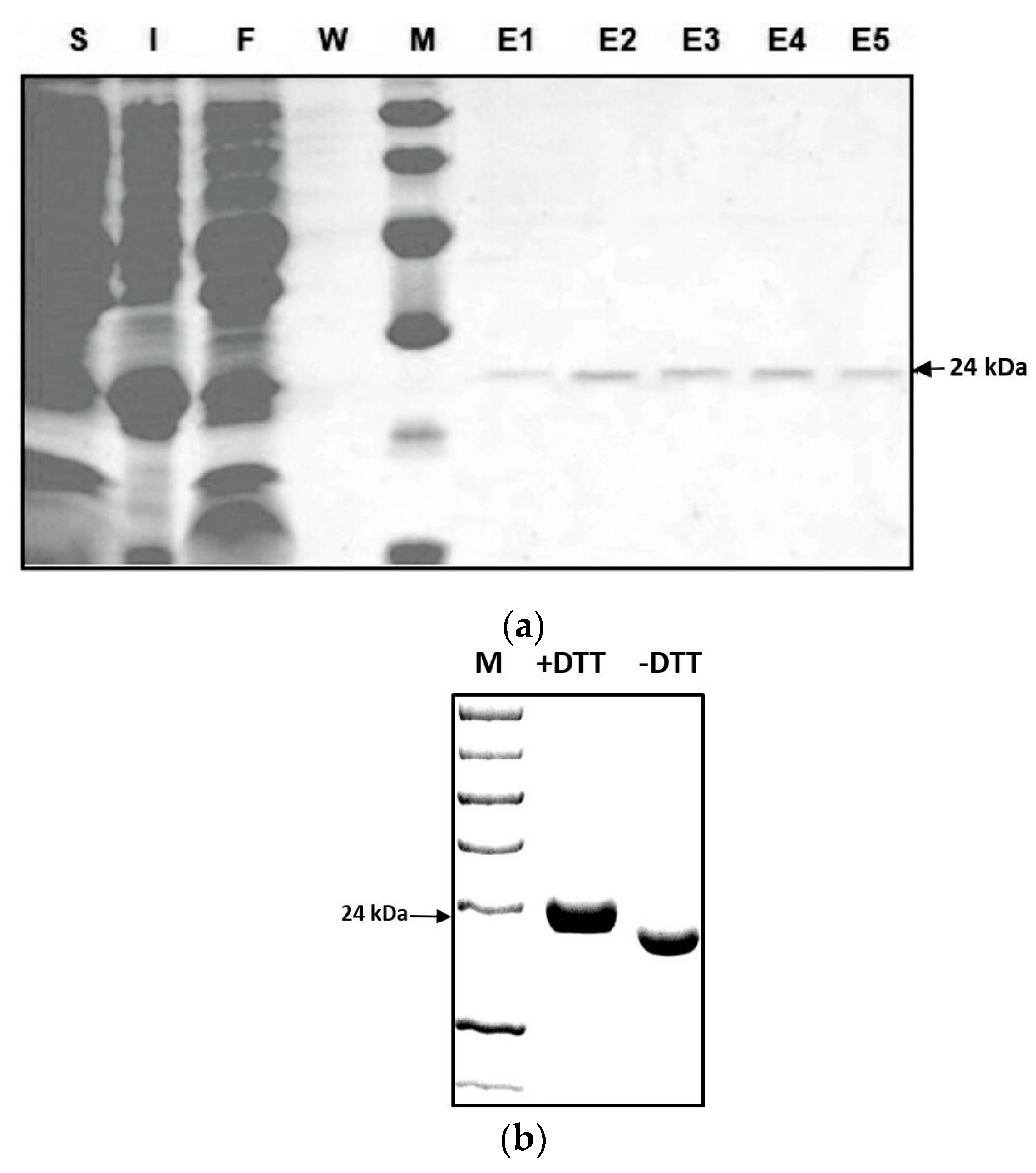
2.2. Characterization of the Zm-INVINH4 Recombinant Protein
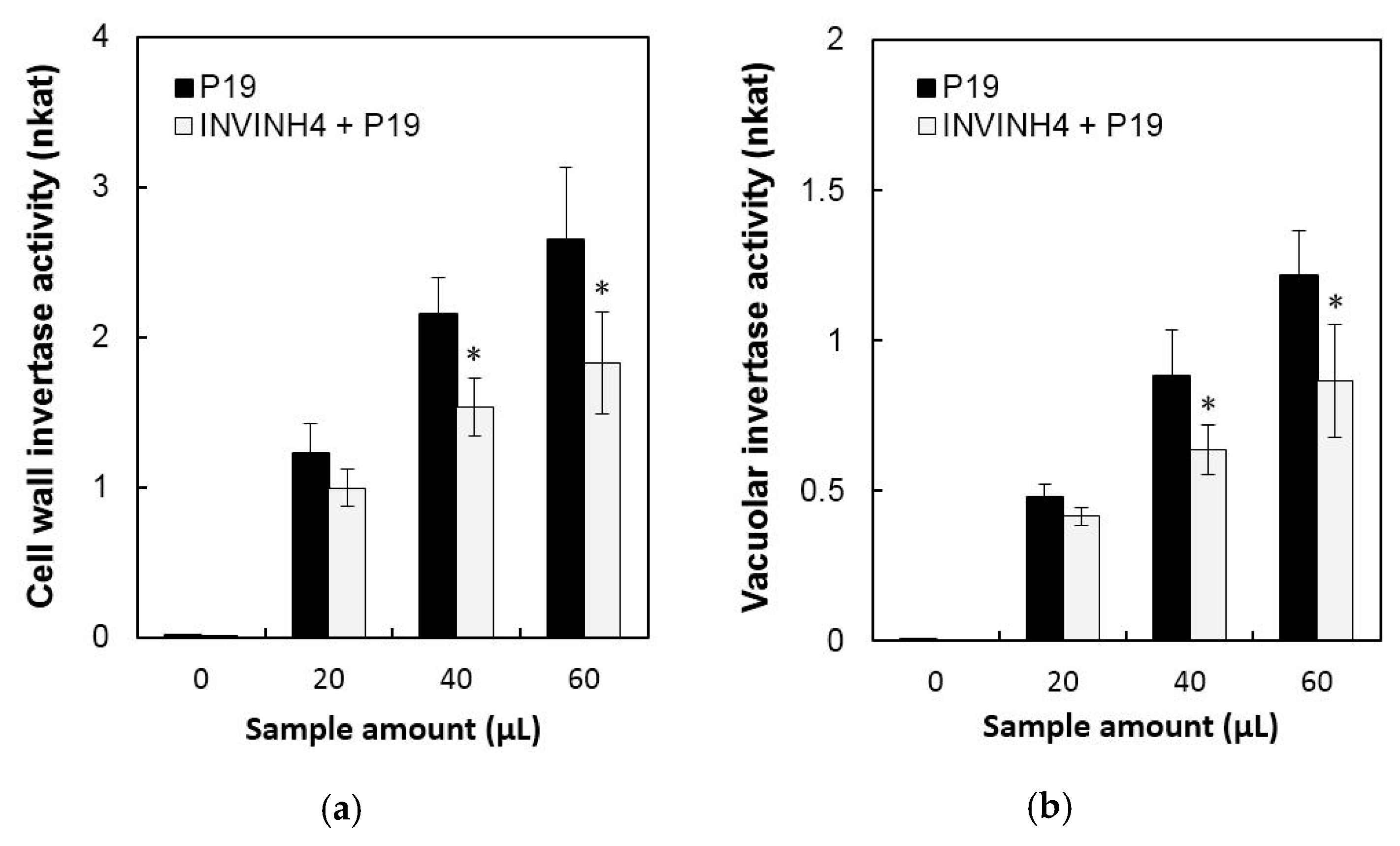
2.3. Transient Expression Assay of Zm-INVINH4 in Potato Leaves
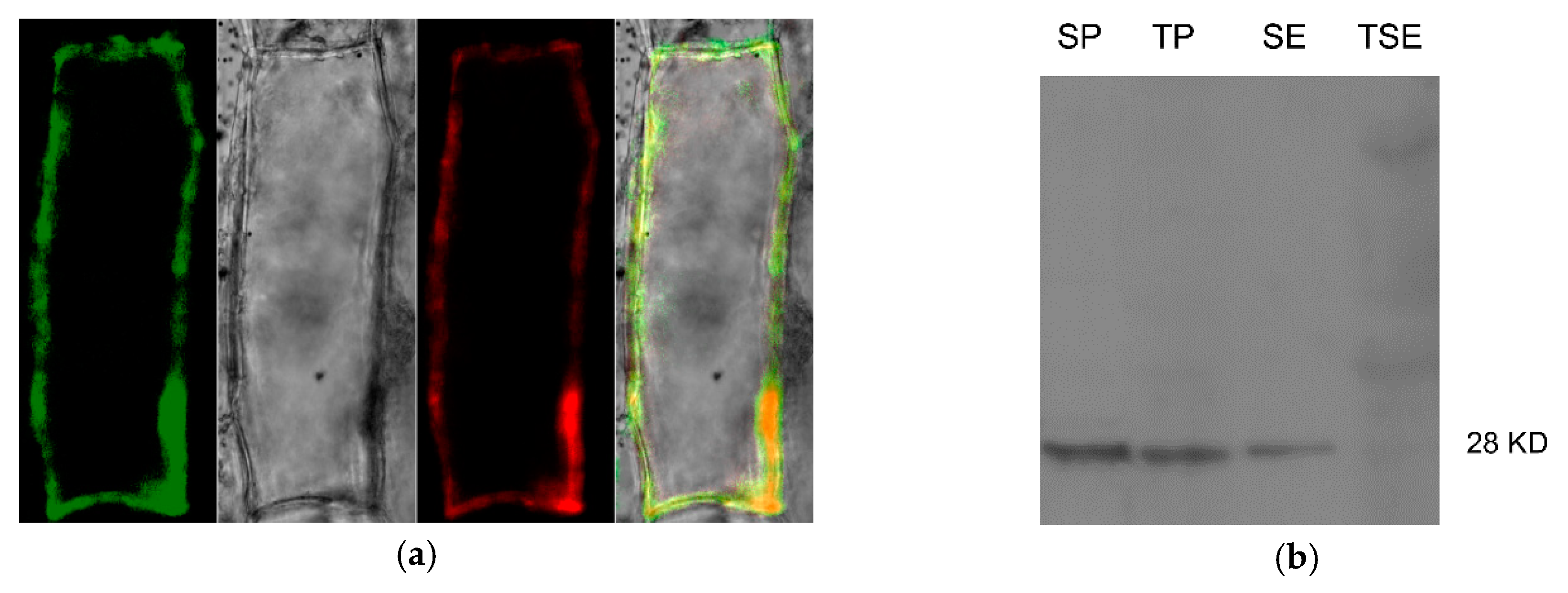
2.4. Subcellular Localization of Zm-INVINH4
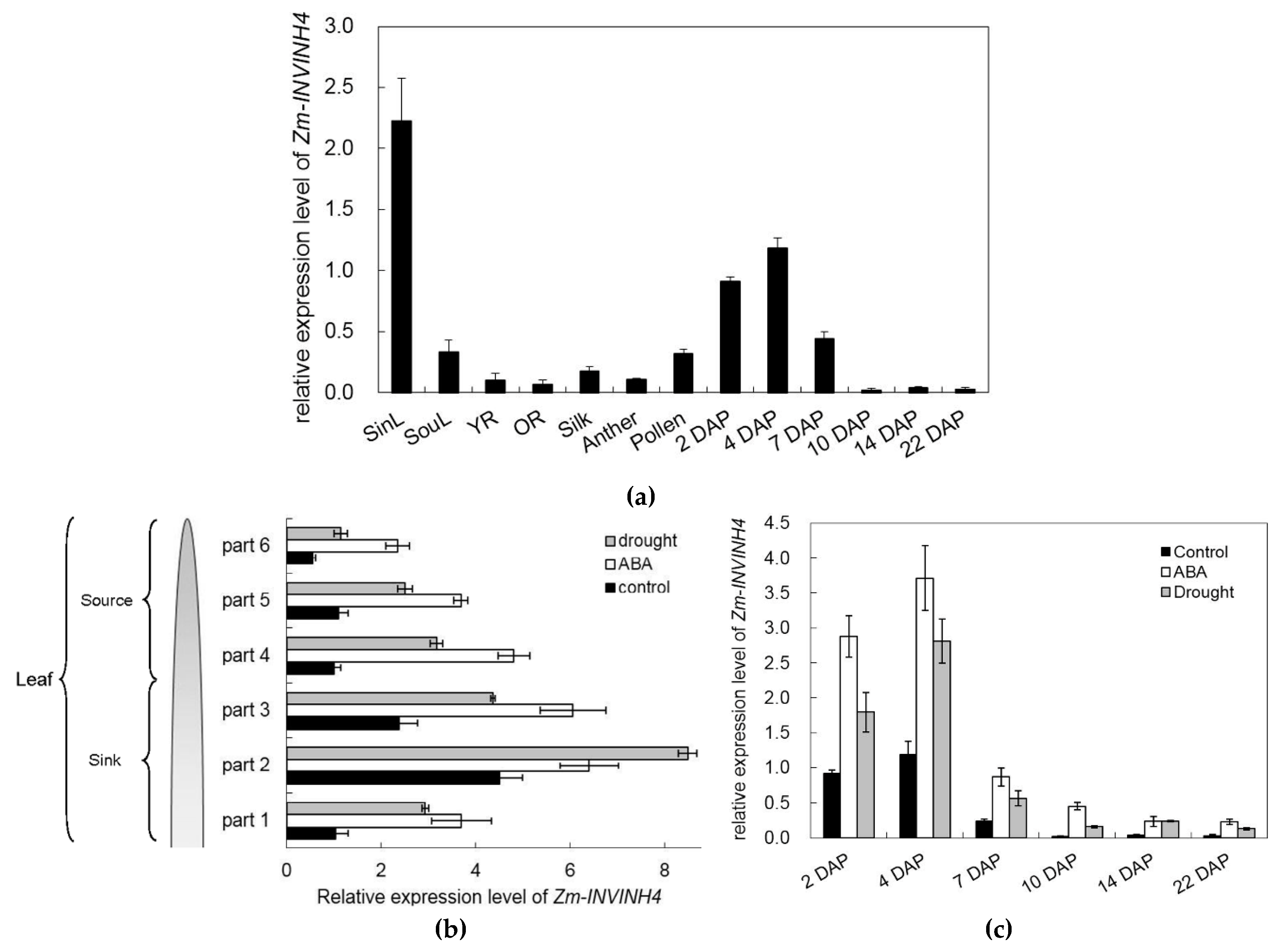
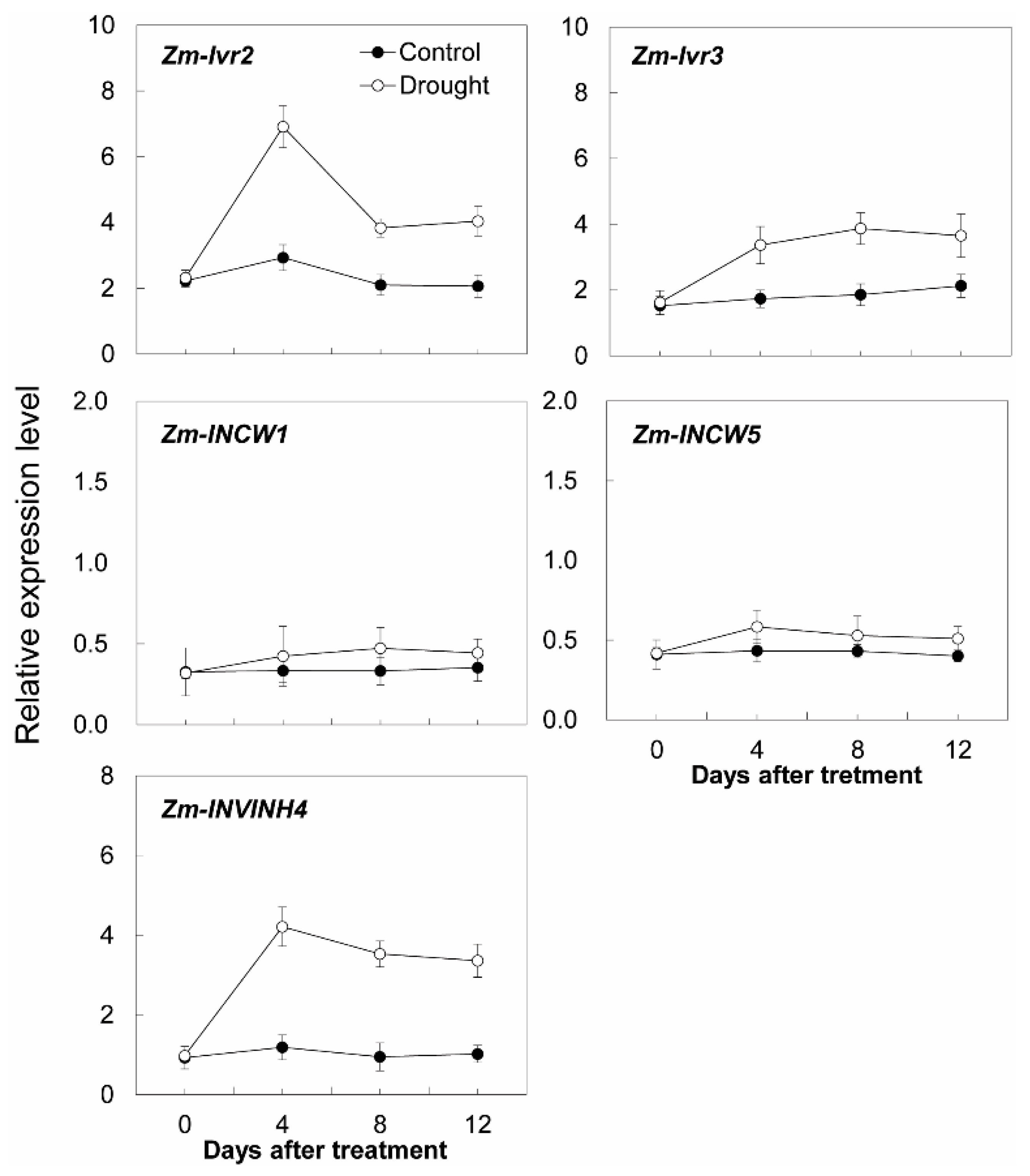
2.5. Expression Pattern of Zm-INVINH4 in Different Maize Tissues under Drought and ABA Stress
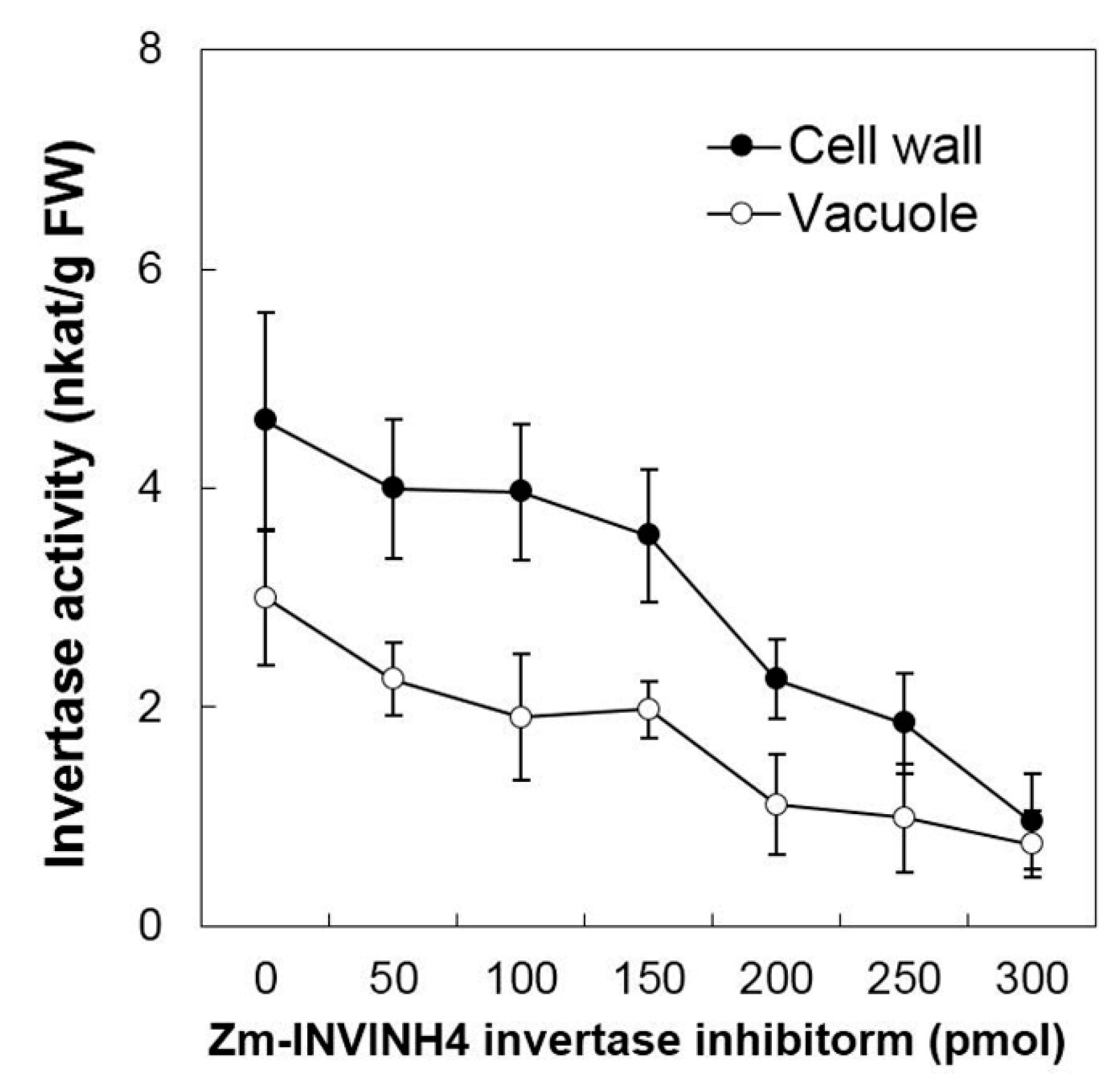
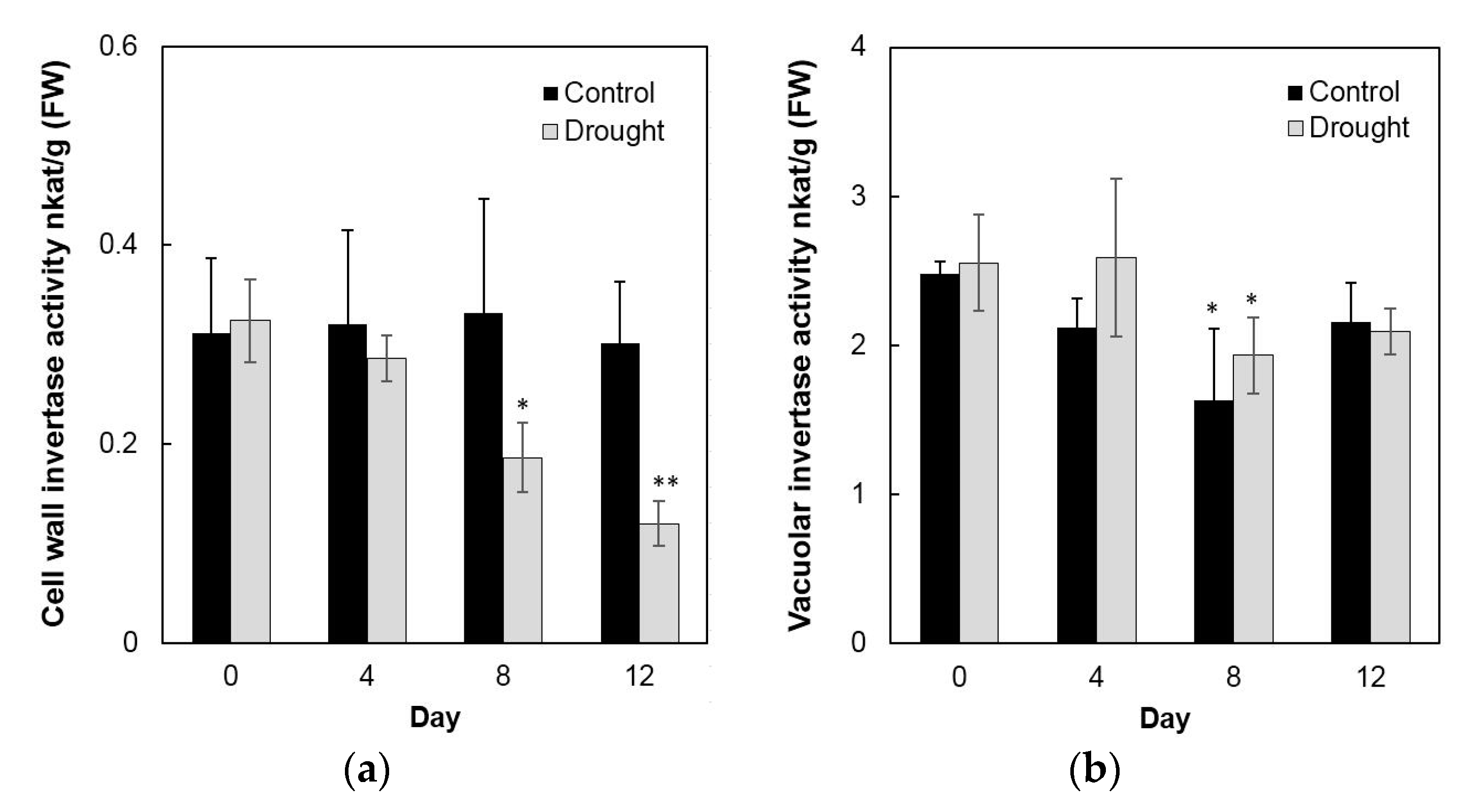
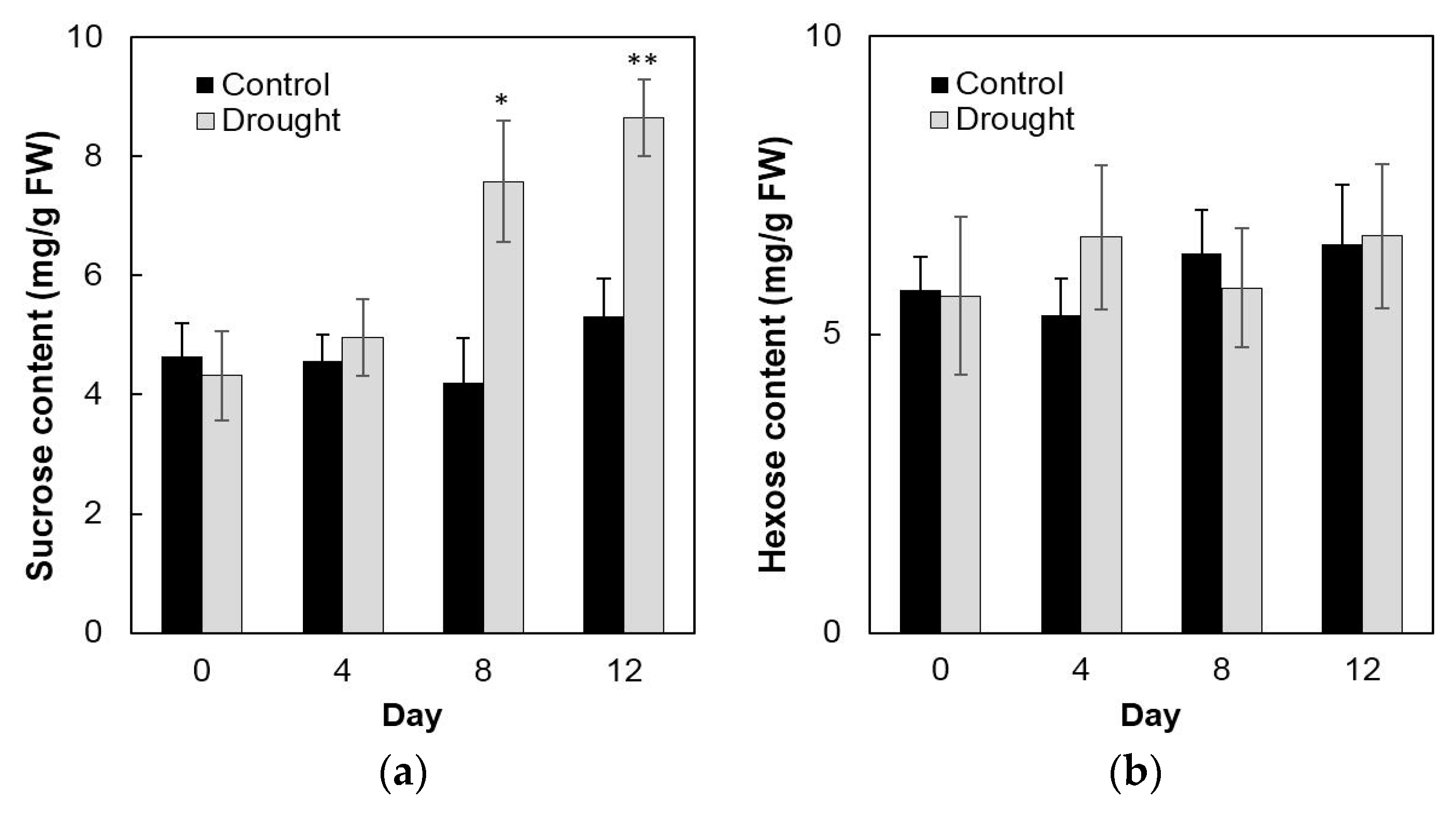
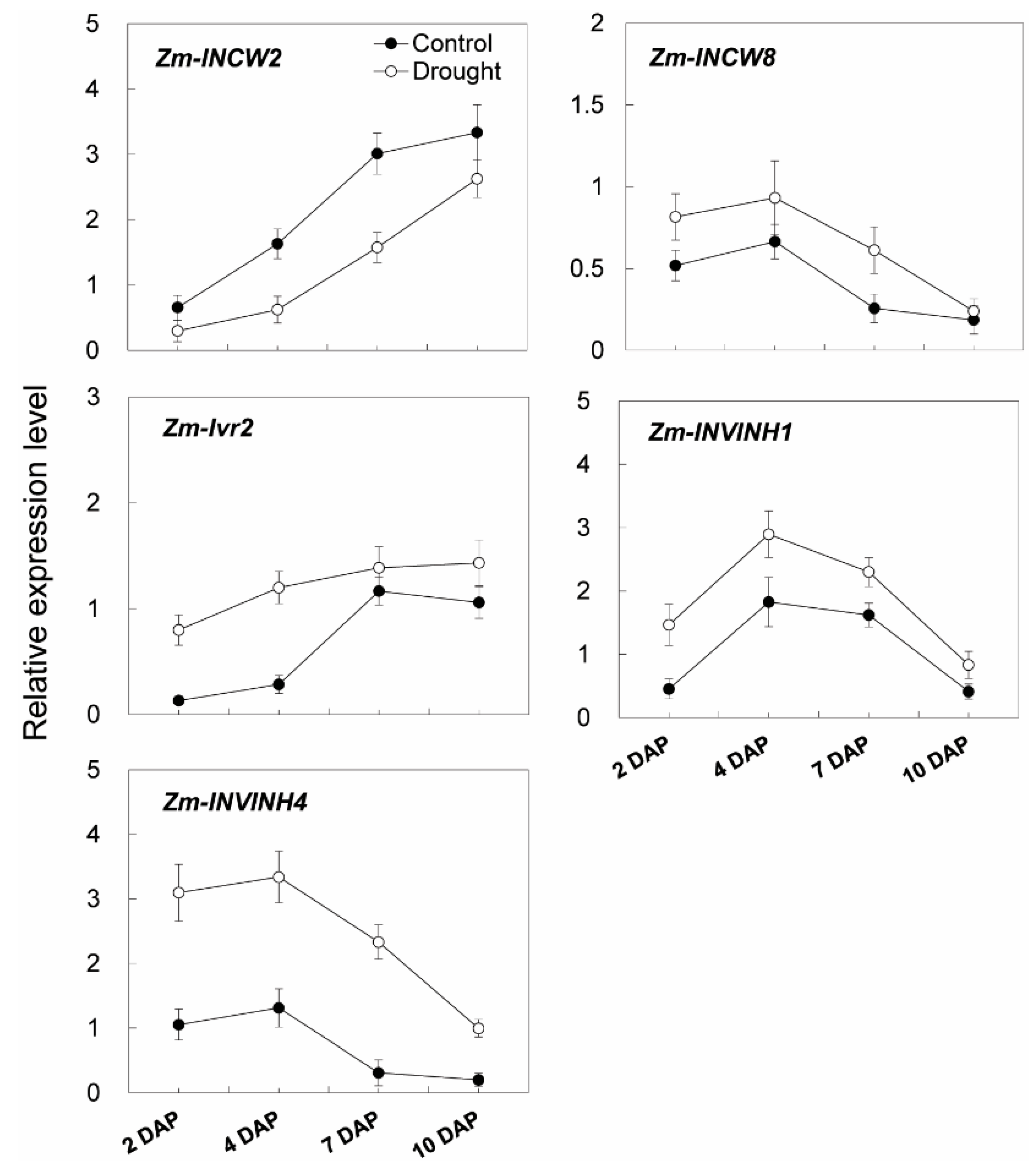
2.6. Invertase Activities, Soluble Sugar Accumulation, and Related Gene Expression in the Sink Part of the Leaf
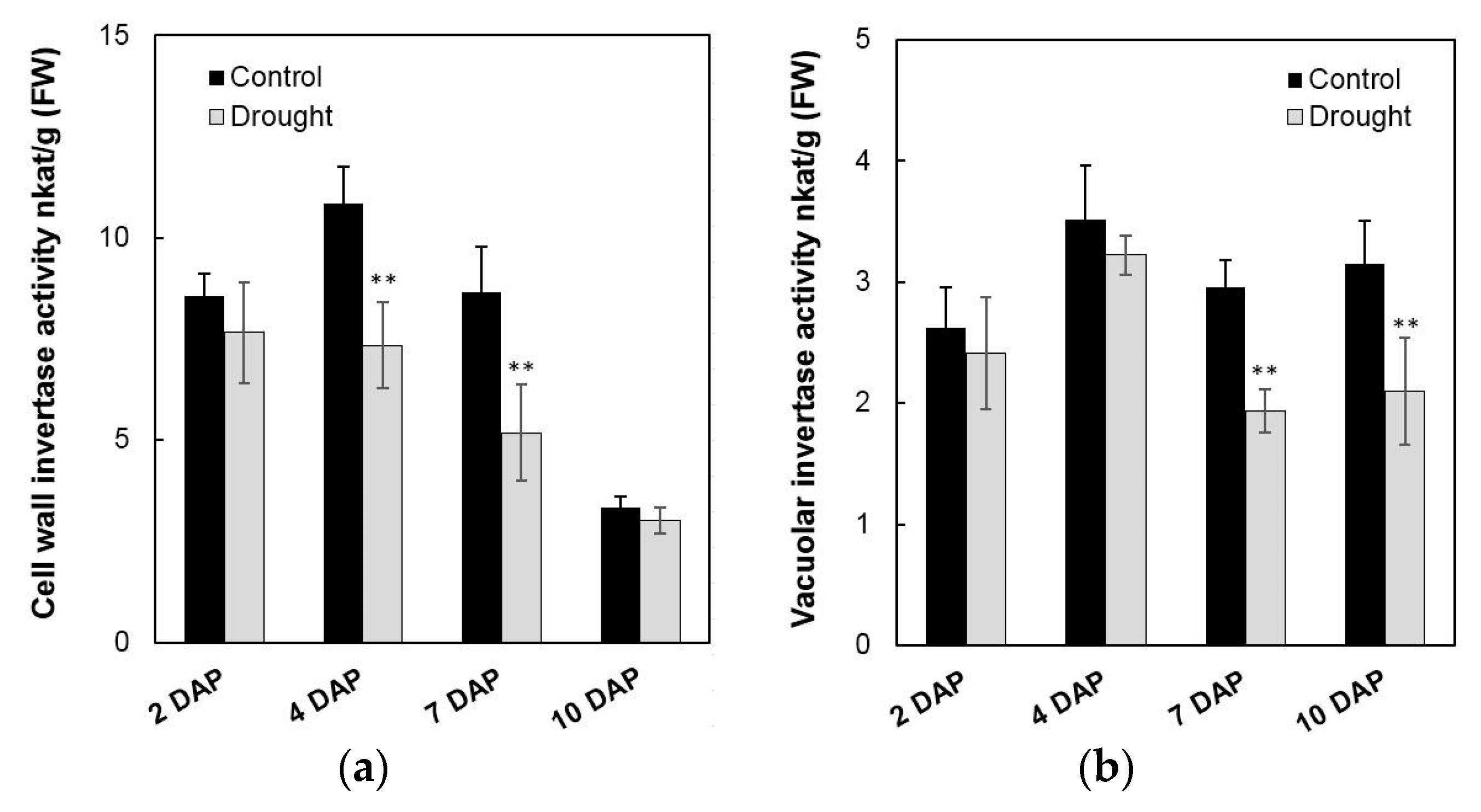
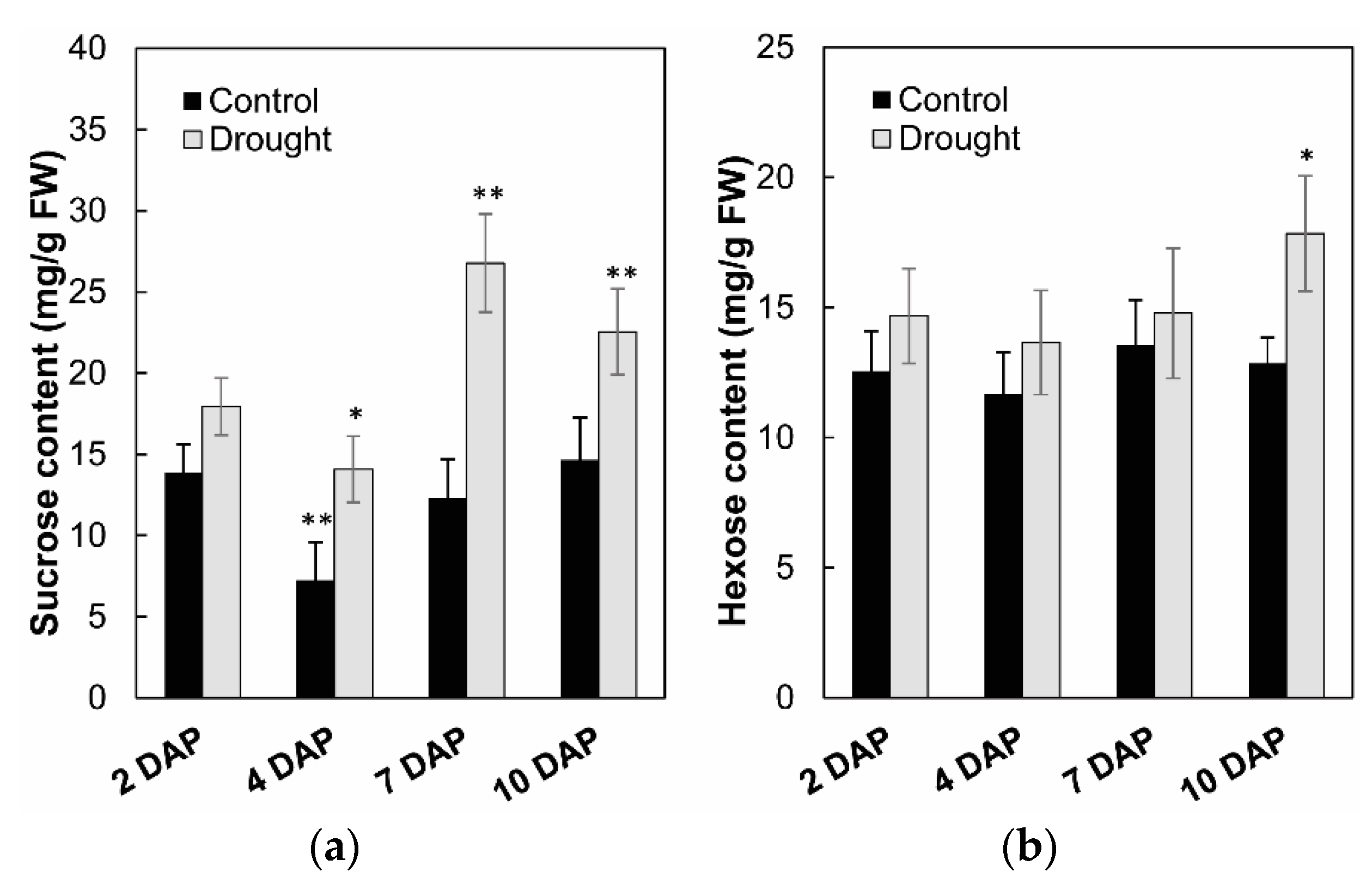
2.7. Effects of Invertase Activities, Soluble Sugar Accumulation, and Related Genes Expression in Young Kernels
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation
4.2. Preparation of RNA, cDNA Synthesis, and cDNA Cloning
4.3. Plasmid Cloning
4.4. Gene Expression Analysis by qPCR
4.5. Expression of Recombinant Zm-INVINH4 Protein in E. coli
4.6. Plant Transformation
4.7. Protein Extraction From Plant Material and Determination of Invertase Activity
4.8. Functional Characterization of E. coli-Expressed Invertase Inhibitor Protein
4.9. CLSM Analysis
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rockwell, F.E.; Gersony, J.T.; Holbrook, N.M. Where does Münch flow begin? Sucrose transport in the pre-phloem path. Curr. Opin. Plant. Biol. 2018, 43, 101–107. [Google Scholar] [CrossRef]
- Kumar, R.; Bishop, E.; Bridges, W.C.; Tharayil, N.; Sekhon, R.S. Sugar partitioning and source-sink interaction are key determinants of leaf senescence in maize. Plant. Cell Env. 2019. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tao, H.; Zhang, B.; Huang, S.; Wang, P. Timing of water deficit limits maize kernel setting in association with changes in the source-flow-sink relationship. Front. Plant. Sci. 2018, 9, 1326. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Lo, S.F.; Ho, T.D. Source-sink communication: Regulated by hormone, nutrient, and stress cross-signaling. Trends Plant. Sci. 2015, 20, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Kim, H.S.; Choi, S.H.; Jang, J.Y.; Jeong, M.J.; Lee, S.I. The importance of the circadian clock in regulating plant metabolism. Int. J. Mol. Sci. 2017, 18, 2680. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Tang, G.Q. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant. Sci. 1999, 4, 401–407. [Google Scholar] [CrossRef]
- Zhu, J.; Qi, J.; Fang, Y.; Xiao, X.; Li, J.; Lan, J.; Tang, C. Characterization of sugar contents and sucrose metabolizing enzymes in developing leaves of hevea brasiliensis. Front. Plant. Sci. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L.; Patrick, J.W.; Bouzayen, M.; Osorio, S.; Fernie, A.R. Molecular regulation of seed and fruit set. Trends Plant. Sci. 2012, 17, 656–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, T.; Han, M.; Min, J.; Chen, P.X.; Mao, Y.X.; Huang, Q.; Tong, Q.; Liu, Q.C.; Fang, Y.M. Genome-wide survey of invertase encoding genes and functional characterization of an extracellular fungal pathogen-responsive invertase in Glycine max. Int. J. Mol. Sci. 2018, 19, 2395. [Google Scholar] [CrossRef]
- Ruan, Y.L.; Jin, Y.; Yang, Y.J.; Li, G.J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant. 2010, 3, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.N.; Asch, F.; Wu, Y.; Jensen, C.R.; Naested, H.; Mogensen, V.O.; Koch, K.E. Soluble invertase expression is an early target of drought stress during the critical, abortion-sensitive phase of young ovary development in maize. Plant. Physiol. 2002, 130, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Linares, L.; Gavilanes-Ruíz, M.; Díaz-Pontones, D.; Guzmán-Chávez, F.; Calzada-Alejo, V.; Zurita-Villegas, V.; Luna-Loaiza, V.; Moreno-Sánchez, R.; Bernal-Lugo, I.; Sánchez-Nieto, S. Early carbon mobilization and radicle protrusion in maize germination. J. Exp. Bot. 2012, 63, 4513–4526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, T.; Luttschwager, D.; Lentzsch, P. Recovery from drought stress at the shooting stage in oilseed rape (Brassica napus). J. Agron. Crop. Sci. 2010, 196, 81–89. [Google Scholar] [CrossRef]
- Li, Z.M.; Palmer, W.M.; Martin, A.P.; Wang, R.Q.; Rainsford, F.; Jin, Y.; Patrick, J.W.; Yang, Y.J.; Ruan, Y.L. High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit. J. Exp. Bot. 2012, 63, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Danijela, M.; Milan, D.; Branislav, S.; Jasmina, N.Z.; Vuk, M.; Ivana, M.; Miroslav, N. Sugars and acid invertase mediate the physiological response of Schenkia spicata root cultures to salt stress. J. Plant. Physiol. 2012, 169, 1281–1289. [Google Scholar]
- Jin, Y.; Ni, D.A.; Ruan, Y.L. Posttranslational elevation of cell wall invertase activity by silencing its inhibitor in tomato delays leaf senescence and increases seed weight and fruit hexose level. Plant. Cell 2009, 21, 2072–2089. [Google Scholar] [CrossRef] [PubMed]
- Rausch, T.; Greiner, S. Plant protein inhibitors of invertases. Biochimica et Biophysica Acta 2004, 1696, 253–261. [Google Scholar] [CrossRef]
- McLaughlin, J.E.; Boyer, J.S. Sugar-responsive gene expression, invertase activity, and senescence in aborting maize ovaries at low water potentials. Ann. Bot. 2004, 94, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.M.; Jiang, T.B.; Fountain, J.C.; Scully, B.T.; Lee, R.D.; Kemerait, R.C.; Chen, S.X.; Guo, B.Z. Protein profiles reveal diverse responsive signaling pathways in kernels of two maize inbred lines with contrasting drought sensitivity. Int. J. Mol. Sci. 2014, 15, 18892–18918. [Google Scholar] [CrossRef]
- Trouverie, J.; Prioul, J.L. Increasing leaf export and grain import capacities in maize plants under water stress. Funct. Plant. Biol. 2006, 33, 209–218. [Google Scholar] [CrossRef]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant. Cell Environ. 2008, 31, 11–38. [Google Scholar] [PubMed]
- Bate, N.J.; Niu, X.; Wang, Y.; Reimann, K.S.; Helentjaris, T.G. An invertase inhibitor from maize localizes to the embryo surrounding region during early kernel development. Plant. Physiol. 2004, 134, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetic analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, P.B.; Venkateshwaran, M.; Wu, L.; Ané, J.M.; Jiang, J. Agrobacterium-mediated transient gene expression and silencing: A rapid tool for functional gene assay in potato. PLoS ONE 2009, 5, e5812. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Rausch, T.; Greiner, S. The N-terminal pro region mediates retention of unprocessed type-I PME in the Golgi apparatus. Plant. J. 2009, 58, 361–375. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.; Wyatt, S.; Tsou, P.L.; Robertson, D.; Allen, N.S. Model system for plant cell biology: GFP imaging in living onion epidermal cells. Biotechniques 1999, 26, 1128–1132. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, M.; D’Angelo, I.; Marquez, J.; Greiner, S.; Scheffzek, K. The invertase inhibitor Nt-CIF from tobacco: A highly thermostable four-helix bundle with an unusual n-terminal extension. J. Mol. Biol. 2004, 335, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, M.; Wolf, S.; Aloy, P.; Greiner, S.; Scheffzek, K. Structural insights into the target specificity of plant invertase and pectin methylesterase inhibitory proteins. Plant. Cell 2004, 16, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Taliercio, E.; Chourey, P. The Miniature1 seed locus of maize encodes a cell wall invertase required for normal development of endosperm and maternal cells in the pedicel. Plant. Cell 1996, 8, 971–983. [Google Scholar] [CrossRef]
- Chourey, P.S.; Jain, M.; Li, Q.B.; Carlson, S.J. Genetic control of cell wall invertases in developing endosperm of maize. Planta 2006, 223, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Chourey, P.S.; Li, Q.B.; Juan, C.C. Pleiotropy and its dissection through a metabolic gene Miniature1 (Mn1) that encodes a cell wall invertase in developing seeds of maize. Plant. Sci. 2012, 184, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Su, T.; Han, M.; Min, J.; Cao, D.; Zhai, G.Q.; Zhou, H.Y.; Li, N.Y.; Li, M.Z. Genome-wide characterization of AspATs in Populus: Gene expression variation and enzyme activities in response to nitrogen perturbations. Forests 2019, 10, 449. [Google Scholar] [CrossRef]
- Eufinger, J. Regulation of taproot development and sucrose stabilization in sugar beet: Influence of invertase inhibitors and occurrence of mitochondrial energy-dissipating proteins. Ph.D. Thesis, University of Heidelberg, Heidelberg, Germany, June 2006. [Google Scholar]
- Link, M.; Rausch, T.; Greiner, S. In Arabidopsis thaliana, the invertase inhibitors AtC/VIF1 and 2 exhibit distinct target enzyme specifcities and expression profiles. FEBS Lett. 2004, 573, 105–109. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Program | Apoplast | ER | Golg | CHL | MT | Other |
|---|---|---|---|---|---|---|
| PSORT | 0.805 a | 0.2 | 0.100 | NA | NA | NA |
| TargetP | 0.642 | NA | NA | 0.025 | 0.248 | 0.006 |
| SignalP | 0.931 a | NA | NA | NA | NA | NA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Liu, X.; Huang, X.; Luo, W.; Long, Y.; Greiner, S.; Rausch, T.; Zhao, H. Functional Characterization of a Drought-Responsive Invertase Inhibitor from Maize (Zea mays L.). Int. J. Mol. Sci. 2019, 20, 4081. https://doi.org/10.3390/ijms20174081
Chen L, Liu X, Huang X, Luo W, Long Y, Greiner S, Rausch T, Zhao H. Functional Characterization of a Drought-Responsive Invertase Inhibitor from Maize (Zea mays L.). International Journal of Molecular Sciences. 2019; 20(17):4081. https://doi.org/10.3390/ijms20174081
Chicago/Turabian StyleChen, Lin, Xiaohong Liu, Xiaojia Huang, Wei Luo, Yuming Long, Steffen Greiner, Thomas Rausch, and Hongbo Zhao. 2019. "Functional Characterization of a Drought-Responsive Invertase Inhibitor from Maize (Zea mays L.)" International Journal of Molecular Sciences 20, no. 17: 4081. https://doi.org/10.3390/ijms20174081
APA StyleChen, L., Liu, X., Huang, X., Luo, W., Long, Y., Greiner, S., Rausch, T., & Zhao, H. (2019). Functional Characterization of a Drought-Responsive Invertase Inhibitor from Maize (Zea mays L.). International Journal of Molecular Sciences, 20(17), 4081. https://doi.org/10.3390/ijms20174081