Cloning and Functional Assessments of Floral-Expressed SWEET Transporter Genes from Jasminum sambac


Abstract
:1. Introduction
2. Results
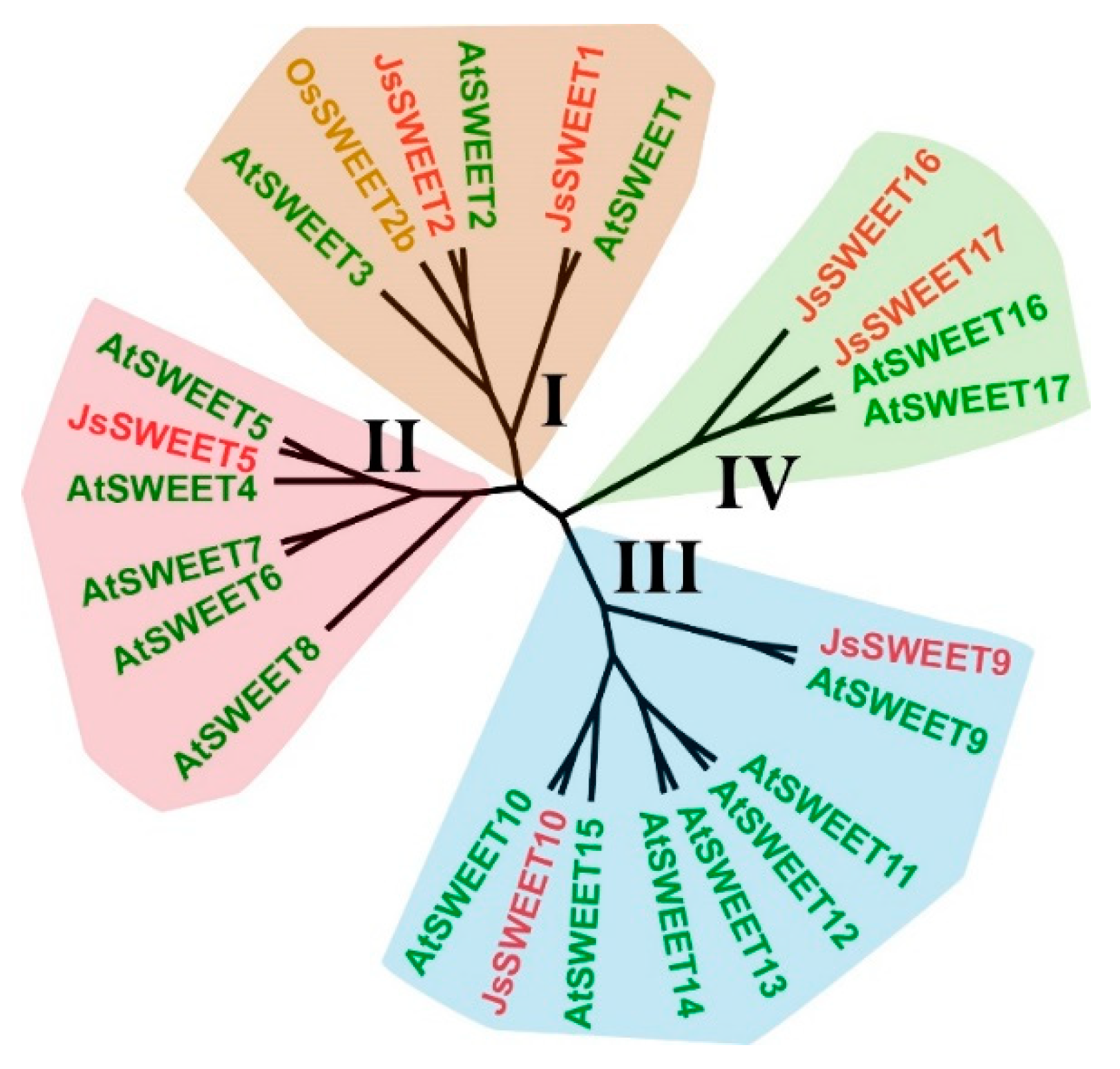
2.1. Identification of Seven Flower-Expressed SWEET Genes Representative of the Four-Clade Family
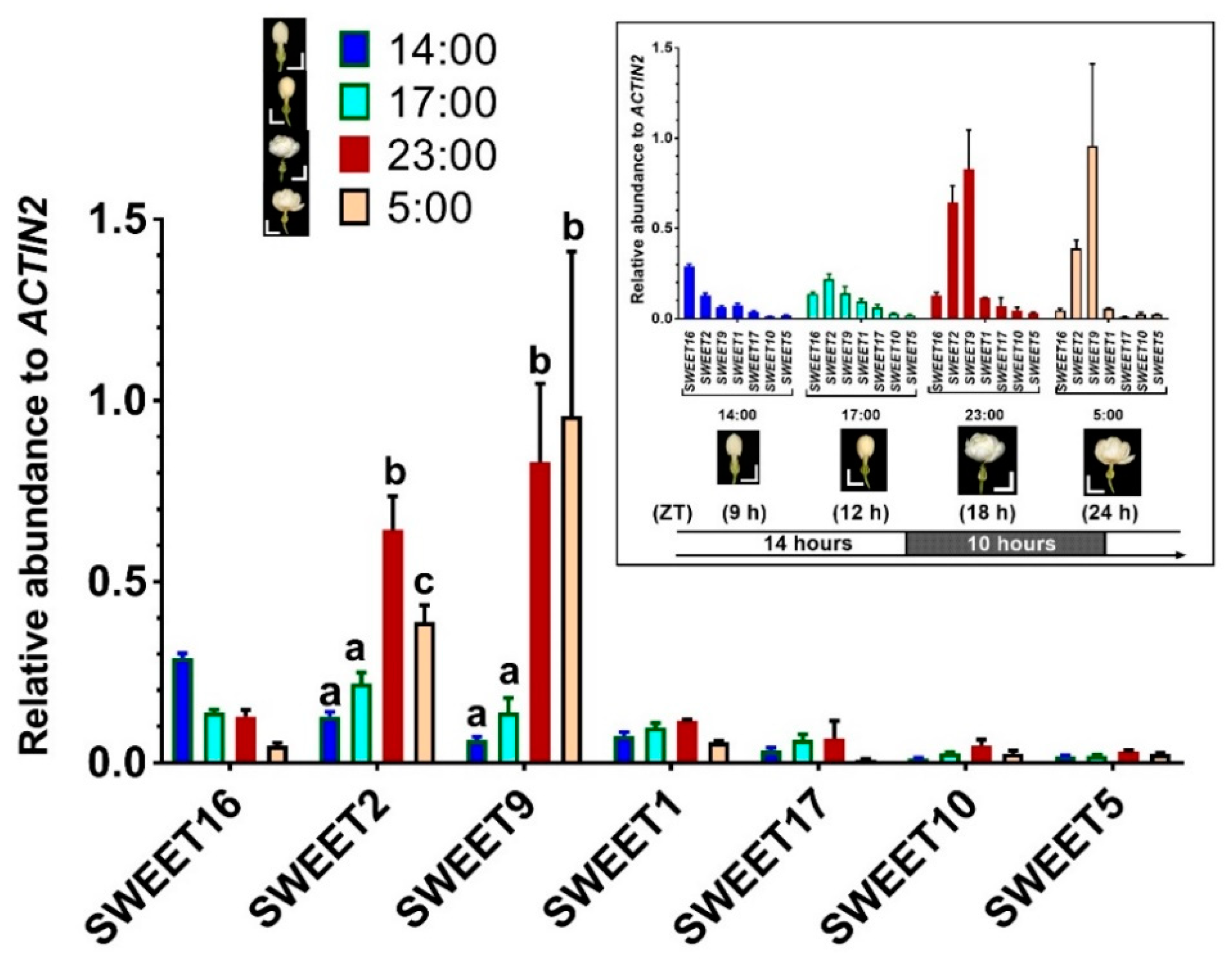
2.2. Expression of the SWEET Genes during Flower Opening Stages
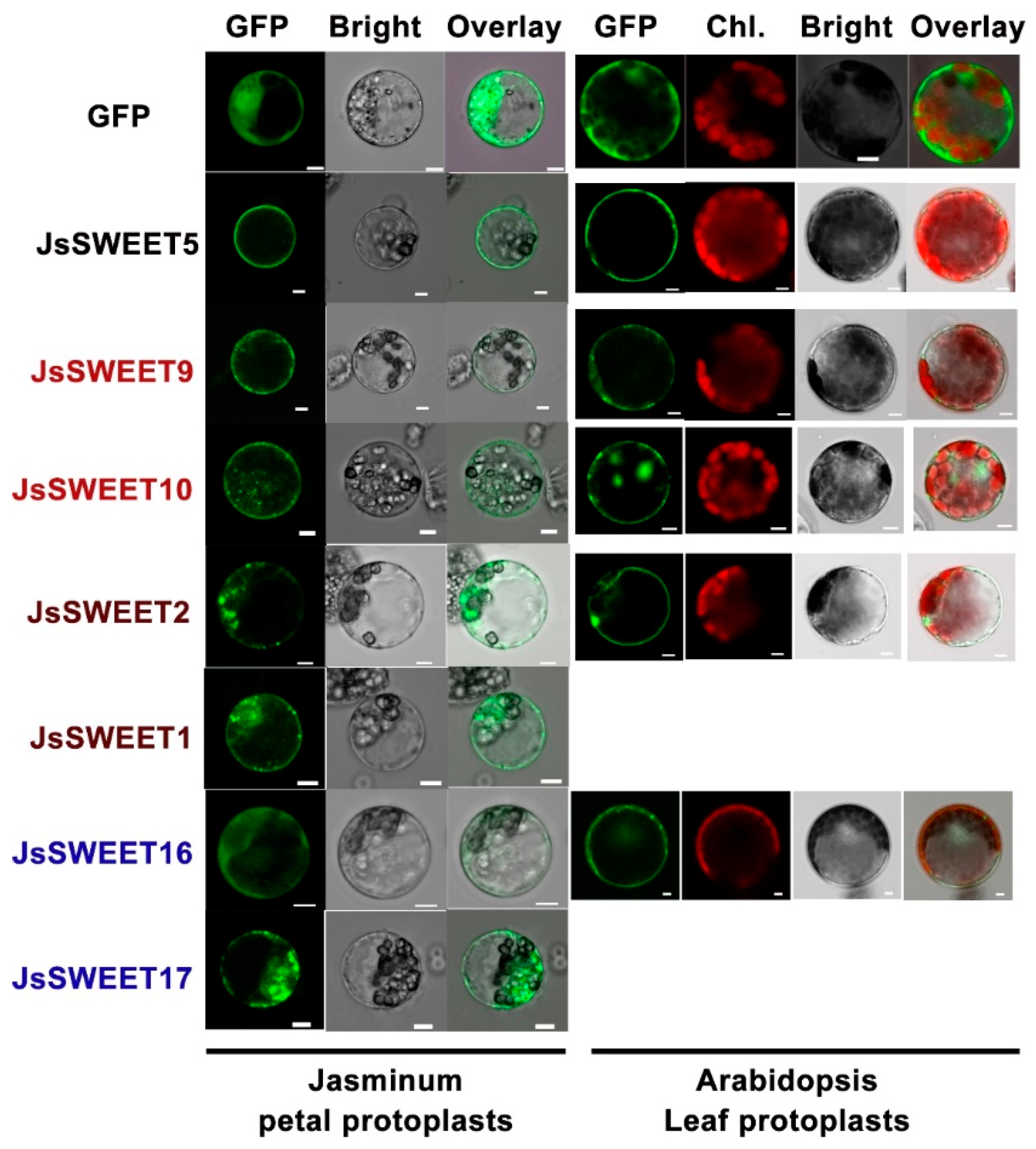
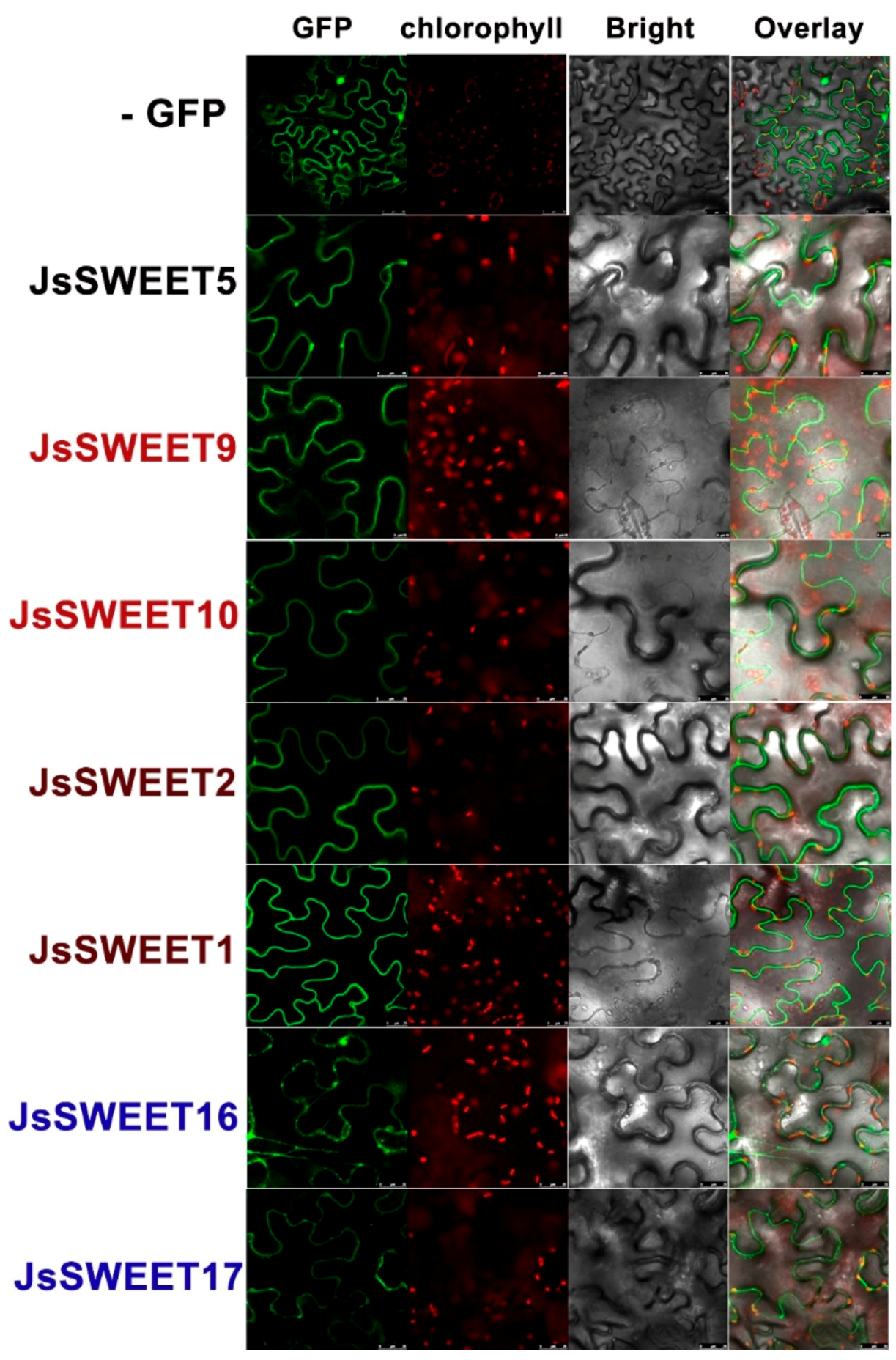
2.3. Subcellular Localization of the Seven SWEET Proteins
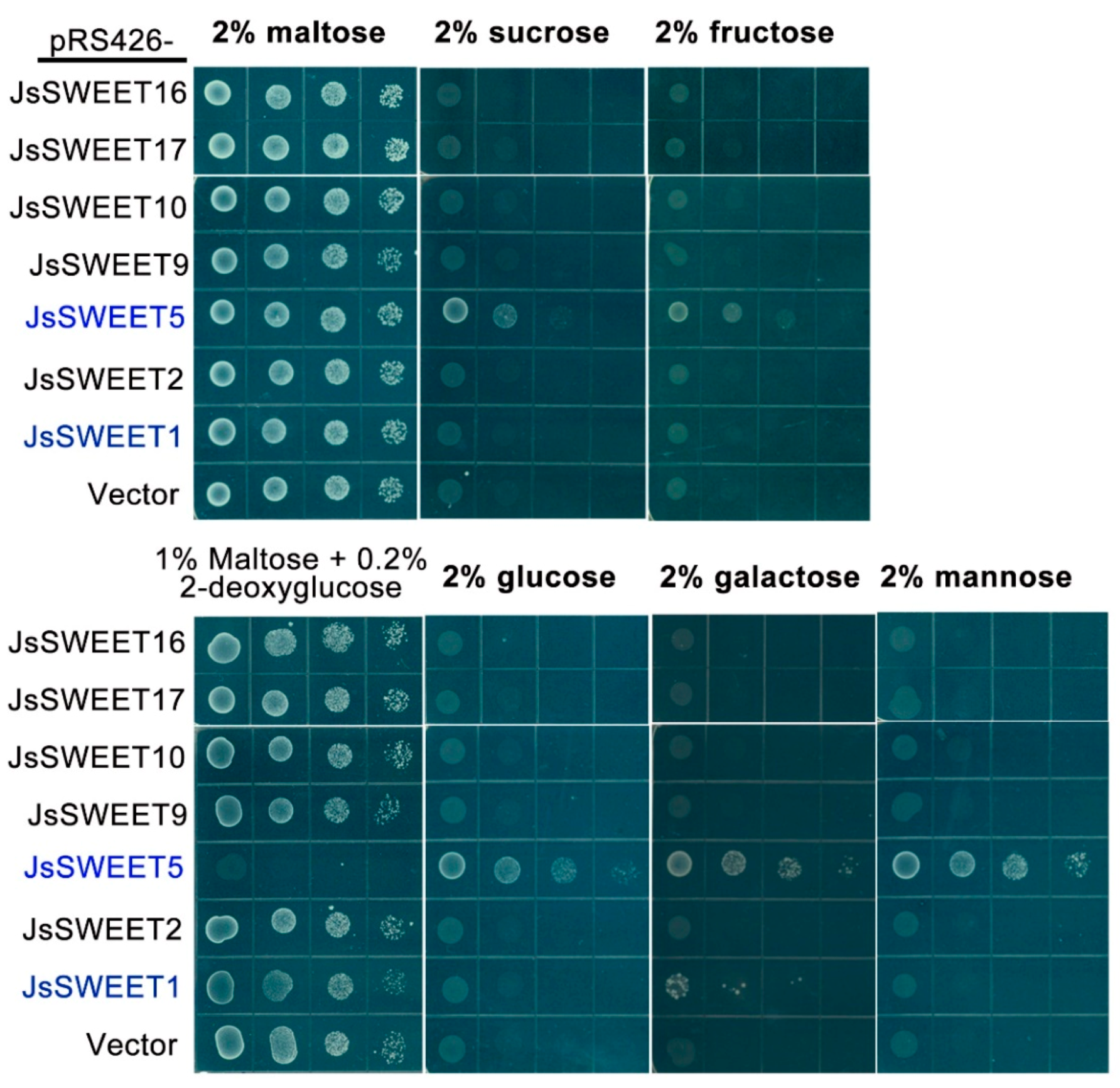
2.4. The JsSWEET5 Is a Multi-Substrates Transporter Revealed by a Yeast Complementation Assay
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA-seq Library Preparation and Transcriptome Generation
4.3. Full-Length cDNA Cloning and Sequence Analysis
4.4. Reverse Transcription Quantitative PCR (RT-PCR)
4.5. Transient Expression and Subcellular Localizaton Analysis
4.6. Yeast Sugar Uptake Complementation Assay
4.7. Genebank Accession
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, S.; Frommer, W.B. Sut sucrose and mst monosaccharide transporter inventory of the selaginella genome. Front. Plant. Sci. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.A.; Thomas, M.A. The monosaccharide transporter gene family in arabidopsis and rice: A history of duplications, adaptive evolution, and functional divergence. Mol. Biol. Evol. 2007, 24, 2412–2423. [Google Scholar] [CrossRef] [PubMed]
- Buttner, M. The monosaccharide transporter(-like) gene family in arabidopsis. FEBS Lett. 2007, 581, 2318–2324. [Google Scholar] [CrossRef] [PubMed]
- Rottmann, T.; Fritz, C.; Sauer, N.; Stadler, R. Glucose uptake via stp transporters inhibits in vitro pollen tube growth in a hexokinase1-dependent manner in arabidopsis thaliana. Plant. Cell 2018, 30, 2057–2081. [Google Scholar] [CrossRef]
- Rottmann, T.; Zierer, W.; Subert, C.; Sauer, N.; Stadler, R. Stp10 encodes a high-affinity monosaccharide transporter and is induced under low-glucose conditions in pollen tubes of arabidopsis. J. Exp. Bot. 2016, 67, 2387–2399. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Saijo, Y.; Nakagami, H.; Takano, Y. Regulation of sugar transporter activity for antibacterial defense in arabidopsis. Science 2016, 354, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Lemonnier, P.; Gaillard, C.; Veillet, F.; Verbeke, J.; Lemoine, R.; Coutos-Thevenot, P.; La Camera, S. Expression of arabidopsis sugar transport protein stp13 differentially affects glucose transport activity and basal resistance to botrytis cinerea. Plant. Mol. Biol. 2014, 85, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Riesmeier, J.W.; Willmitzer, L.; Frommer, W.B. Evidence for an essential role of the sucrose transporter in phloem loading and assimilate partitioning. EMBO J. 1994, 13, 1–7. [Google Scholar] [CrossRef]
- Ayre, B.G. Membrane-transport systems for sucrose in relation to whole-plant carbon partitioning. Mol. Plant. 2011, 4, 377–394. [Google Scholar] [CrossRef]
- Kuhn, C.; Grof, C.P. Sucrose transporters of higher plants. Curr. Opin. Plant. Biol. 2010, 13, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Mainson, D.; Porcheron, B.; Maurousset, L.; Lemoine, R.; Pourtau, N. Carbon source–sink relationship in arabidopsis thaliana: The role of sucrose transporters. Planta 2018, 247, 587–611. [Google Scholar] [CrossRef] [PubMed]
- Latorraca, N.R.; Fastman, N.M.; Venkatakrishnan, A.J.; Frommer, W.B.; Dror, R.O.; Feng, L. Mechanism of substrate translocation in an alternating access transporter. Cell 2017, 169, 96–107.e12. [Google Scholar] [CrossRef] [PubMed]
- Eom, J.S.; Chen, L.Q.; Sosso, D.; Julius, B.T.; Lin, I.W.; Qu, X.Q.; Braun, D.M.; Frommer, W.B. Sweets, transporters for intracellular and intercellular sugar translocation. Curr. Opin. Plant. Biol. 2015, 25, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by sweet proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Hou, B.H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.Q.; Guo, W.J.; Kim, J.G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Cheung, L.S.; Li, S.; Eom, J.S.; Chen, L.Q.; Xu, Y.; Perry, K.; Frommer, W.B.; Feng, L. Structure of a eukaryotic sweet transporter in a homotrimeric complex. Nature 2015, 527, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.H.; Hu, Y.B.; Chen, L.Q.; Sosso, D.; Ducat, D.C.; Hou, B.H.; Frommer, W.B. Functional role of oligomerization for bacterial and plant sweet sugar transporter family. Proc. Natl. Acad. Sci. USA 2013, 110, E3685–E3694. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhu, Y.; Liu, M.; Zhou, Y.; Lu, G.; Lan, L.; Wang, X.; Zhao, Y.; Zhang, X.C. Molecular mechanism of substrate recognition and transport by the atsweet13 sugar transporter. Proc. Natl. Acad. Sci. USA 2017, 114, 10089–10094. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Tao, Y.; Cheung, L.S.; Fan, C.; Chen, L.Q.; Xu, S.; Perry, K.; Frommer, W.B.; Feng, L. Structures of bacterial homologues of sweet transporters in two distinct conformations. Nature 2014, 515, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J.; et al. Soybean (glycine max) sweet gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genome 2015, 16, 520. [Google Scholar] [CrossRef] [PubMed]
- Gautam, T.; Saripalli, G.; Gahlaut, V.; Kumar, A.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Further studies on sugar transporter (sweet) genes in wheat (Triticum aestivum L.). Mol. Biol. Rep. 2019, 46, 2327–2353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, S.; Yu, F.; Tang, J.; Shan, X.; Bao, K.; Yu, L.; Wang, H.; Fei, Z.; Li, J. Genome-wide characterization and expression profiling of sweet genes in cabbage (Brassica oleracea var. Capitata L.) reveal their roles in chilling and clubroot disease responses. BMC Genom. 2019, 20, 93. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ren, Z.; Wang, Z.; Sun, K.; Pei, X.; Liu, Y.; He, K.; Zhang, F.; Song, C.; Zhou, X.; et al. Evolution and stress responses of gossypium hirsutum sweet genes. Int. J. Mol. Sci. 2018, 19, 769. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.L.; Xiao, X.H.; Qi, J.Y.; Fang, Y.J.; Tang, C.R. The sweet gene family in hevea brasiliensis—Its evolution and expression compared with four other plant species. FEBS Open Biol. 2017, 7, 1943–1959. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Zhu, X.F.; Pu, Z.J.; Duan, Y.X.; Hao, L.J.; Zhang, J.; Chen, L.Q.; Jeon, C.O.; Xuan, Y.H. Integrative view of the diversity and evolution of sweet and semisweet sugar transporters. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef]
- Jeena, G.S.; Kumar, S.; Shukla, R.K. Structure, evolution and diverse physiological roles of sweet sugar transporters in plants. Plant. Mol. Biol. 2019, 100, 351–365. [Google Scholar] [CrossRef]
- Bezrutczyk, M.; Yang, J.; Eom, J.S.; Prior, M.; Sosso, D.; Hartwig, T.; Szurek, B.; Oliva, R.; Vera-Cruz, C.; White, F.F.; et al. Sugar flux and signaling in plant-microbe interactions. Plant. J.: Cell Mol. Biol. 2018, 93, 675–685. [Google Scholar] [CrossRef]
- Bezrutczyk, M.; Hartwig, T.; Horshman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired phloem loading in zmsweet13a,b,c sucrose transporter triple knock-out mutants in zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Zhang, H.; Zhang, Q.; Zhai, H.; Liu, Q.; He, S. The plasma membrane-localized sucrose transporter ibsweet10 contributes to the resistance of sweet potato to fusarium oxysporum. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef]
- Chen, H.-Y.; Huh, J.-H.; Yu, Y.-C.; Ho, L.-H.; Chen, L.-Q.; Tholl, D.; Frommer, W.B.; Guo, W.-J. The arabidopsis vacuolar sugar transporter sweet2 limits carbon sequestration from roots and restricts pythium infection. Plant. J.: Cell Mol. Biol. 2015, 83, 1046–1058. [Google Scholar] [CrossRef]
- Lin, I.W.; Sosso, D.; Chen, L.Q.; Gase, K.; Kim, S.G.; Kessler, D.; Klinkenberg, P.M.; Gorder, M.K.; Hou, B.H.; Qu, X.Q.; et al. Nectar secretion requires sucrose phosphate synthases and the sugar transporter sweet9. Nature 2014, 508, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Sosso, D.; Luo, D.; Li, Q.B.; Sasse, J.; Yang, J.; Gendrot, G.; Suzuki, M.; Koch, K.E.; McCarty, D.R.; Chourey, P.S.; et al. Seed filling in domesticated maize and rice depends on sweet-mediated hexose transport. Nat. Genet. 2015, 47, 1489–1493. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.L.; Luo, D.P.; Yang, B.; Frommer, W.B.; Eom, J.S. Sweet11 and 15 as key players in seed filling in rice. New Phytol. 2018, 218, 604–615. [Google Scholar] [CrossRef]
- Zhang, Z.; Zou, L.; Ren, C.; Ren, F.; Wang, Y.; Fan, P.; Li, S.; Liang, Z. Vvsweet10 mediates sugar accumulation in grapes. Genes 2019, 10, 255. [Google Scholar] [CrossRef] [PubMed]
- Borghi, M.; Fernie, A.R. Floral metabolism of sugars and amino acids: Implications for pollinators’ preferences and seed and fruit set. Plant. Physiol. 2017, 175, 1510–1524. [Google Scholar] [CrossRef]
- Guan, Y.F.; Huang, X.Y.; Zhu, J.; Gao, J.F.; Zhang, H.X.; Yang, Z.N. Ruptured pollen grain1, a member of the mtn3/saliva gene family, is crucial for exine pattern formation and cell integrity of microspores in arabidopsis. Plant. Physiol. 2008, 147, 852–863. [Google Scholar] [CrossRef]
- Ge, Y.X.; Angenent, G.C.; Dahlhaus, E.; Franken, J.; Peters, J.; Wullems, G.J.; Creemers-Molenaar, J. Partial silencing of the nec1 gene results in early opening of anthers in petunia hybrida. Mol. Genet. Genom.~Mgg 2001, 265, 414–423. [Google Scholar]
- Deng, Y.M.; Sun, X.B.; Gu, C.S.; Jia, X.P.; Liang, L.J.; Su, J.L. Identification of pre-fertilization reproductive barriers and the underlying cytological mechanism in crosses among three petal-types of Jasminum sambac and their relevance to phylogenetic relationships. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Deng, Y.M.; Jia, X.P.; Liang, L.J.; Gu, C.S.; Sun, X.B. Morphological anatomy, sporogenesis and gametogenesis in flowering process of jasmine (jasminum sambac aiton). Sci. Hortic.-Amst. 2016, 198, 257–266. [Google Scholar] [CrossRef]
- Bera, P.; Mukherjee, C.; Mitra, A. Enzymatic production and emission of floral scent volatiles in jasminum sambac. Plant. Sci 2017, 256, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.X.; Angenent, G.C.; Wittich, P.E.; Peters, J.; Franken, J.; Busscher, M.; Zhang, L.M.; Dahlhaus, E.; Kater, M.M.; Wullems, G.J.; et al. Nec1, a novel gene, highly expressed in nectary tissue of petunia hybrida. Plant. J.: Cell Mol. Biol. 2000, 24, 725–734. [Google Scholar] [CrossRef]
- Wieczorke, R.; Krampe, S.; Weierstall, T.; Freidel, K.; Hollenberg, C.P.; Boles, E. Concurrent knock-out of at least 20 transporter genes is required to block uptake of hexoses in saccharomyces cerevisiae. FEBS Lett. 1999, 464, 123–128. [Google Scholar] [CrossRef]
- Sun, L.; Sui, X.; Lucas, W.J.; Li, Y.; Feng, S.; Ma, S.; Fan, J.; Gao, L.; Zhang, Z. Down-regulation of the sucrose transporter cssut1 causes male sterility by altering carbohydrate supply. Plant. Physiol. 2019, 180, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yokosho, K.; Guo, R.; Whelan, J.; Ruan, Y.L.; Ma, J.F.; Shou, H. The soybean sugar transporter gmsweet15 mediates sucrose export from endosperm to early embryo. Plant. Physiol. 2019, 180, 2133–2141. [Google Scholar] [CrossRef] [PubMed]
- Chardon, F.; Bedu, M.; Calenge, F.; Klemens, P.A.W.; Spinner, L.; Clement, G.; Chietera, G.; Leran, S.; Ferrand, M.; Lacombe, B.; et al. Leaf fructose content is controlled by the vacuolar transporter sweet17 in arabidopsis. Curr. Biol. 2013, 23, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.J.; Nagy, R.; Chen, H.Y.; Pfrunder, S.; Yu, Y.C.; Santelia, D.; Frommer, W.B.; Martinoia, E. Sweet17, a facilitative transporter, mediates fructose transport across the tonoplast of arabidopsis roots and leaves. Plant. Physiol. 2014, 164, 777–789. [Google Scholar] [CrossRef]
- Klemens, P.A.; Patzke, K.; Deitmer, J.; Spinner, L.; Le Hir, R.; Bellini, C.; Bedu, M.; Chardon, F.; Krapp, A.; Neuhaus, H.E. Overexpression of the vacuolar sugar carrier atsweet16 modifies germination, growth, and stress tolerance in arabidopsis. Plant. Physiol. 2013, 163, 1338–1352. [Google Scholar] [CrossRef]
- Bock, K.W.; Honys, D.; Ward, J.M.; Padmanaban, S.; Nawrocki, E.P.; Hirschi, K.D.; Twell, D.; Sze, H. Integrating membrane transport with male gametophyte development and function through transcriptomics. Plant. Physiol. 2006, 140, 1151–1168. [Google Scholar] [CrossRef]
- Engel, M.L.; Davis, R.H.; McCormick, S. Green sperm. Identification of male gamete promoters in arabidopsis. Plant. Physiol. 2005, 138, 2124–2133. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from rna-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. Degseq: An r package for identifying differentially expressed genes from rna-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for rna-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time pcr data by the comparative c(t) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Inze, D.; Depicker, A. Gateway vectors for agrobacterium-mediated plant transformation. Trends Plant. Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Shen, J.; Fu, J.; Ma, J.; Wang, X.; Gao, C.; Zhuang, C.; Wan, J.; Jiang, L. Isolation, culture, and transient transformation of plant protoplasts. Curr. Protoc. Cell Biol. 2014, 63, 2.8.1–2.8.17. [Google Scholar]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Li, X.; Chanroj, S.; Wu, Z.; Romanowsky, S.M.; Harper, J.F.; Sze, H. A distinct endosomal ca2+/mn2+ pump affects root growth through the secretory process. Plant. Physiol. 2008, 147, 1675–1689. [Google Scholar] [CrossRef]
- Mumberg, D.; Muller, R.; Funk, M. Regulatable promoters of saccharomyces cerevisiae: Comparison of transcriptional activity and their use for heterologous expression. Nucleic Acids Res. 1994, 22, 5767–5768. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Unigene_id | Flowers @11:00 (FPKM) 1 | Flowers @17:00 (FPKM) | Flowers @5:00 (FPKM) | Leaves @17:00 (FPKM) | Leaves @5:00 (FPKM) | Swissprot Evalue |
|---|---|---|---|---|---|---|
| c41111_g1 | 2.775 | 2.435 | 2.035 | 0 | 0 | 1.6 × 10−4 |
| c50045_g1 | 15.92 | 14.29 | 11.59 | 0 | 0 | 1.7 × 10−71 |
| c51046_g2 | 2.655 | 6.975 | 39.445 | 0 | 0 | 6.4 × 10−86 |
| c51507_g1 | 14.165 | 20.89 | 43.425 | 0 | 0 | 9.6 × 10−106 |
| c44769_g1 | 36.935 | 72.54 | 28.14 | 0 | 0.075 | 2.07 × 10−66 |
| c42726_g1 | 4.95 | 11.43 | 9.86 | 13.32 | 2.62 | 1.2 × 10−56 |
| c60775_g2 | 55.56 | 36.72 | 106.135 | 7.59 | 7.755 | 4.8 × 10−87 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Wei, P.; Niu, F.; Liu, X.; Zhang, H.; Lyu, M.; Yuan, Y.; Wu, B. Cloning and Functional Assessments of Floral-Expressed SWEET Transporter Genes from Jasminum sambac. Int. J. Mol. Sci. 2019, 20, 4001. https://doi.org/10.3390/ijms20164001
Wang P, Wei P, Niu F, Liu X, Zhang H, Lyu M, Yuan Y, Wu B. Cloning and Functional Assessments of Floral-Expressed SWEET Transporter Genes from Jasminum sambac. International Journal of Molecular Sciences. 2019; 20(16):4001. https://doi.org/10.3390/ijms20164001
Chicago/Turabian StyleWang, Panpan, Peining Wei, Fangfei Niu, Xiaofeng Liu, Hongliang Zhang, Meiling Lyu, Yuan Yuan, and Binghua Wu. 2019. "Cloning and Functional Assessments of Floral-Expressed SWEET Transporter Genes from Jasminum sambac" International Journal of Molecular Sciences 20, no. 16: 4001. https://doi.org/10.3390/ijms20164001