The Effects of Different Dynamic Culture Systems on Cell Proliferation and Osteogenic Differentiation in Human Mesenchymal Stem Cells

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Cell Proliferation
2.2. Mitochondrial Activity
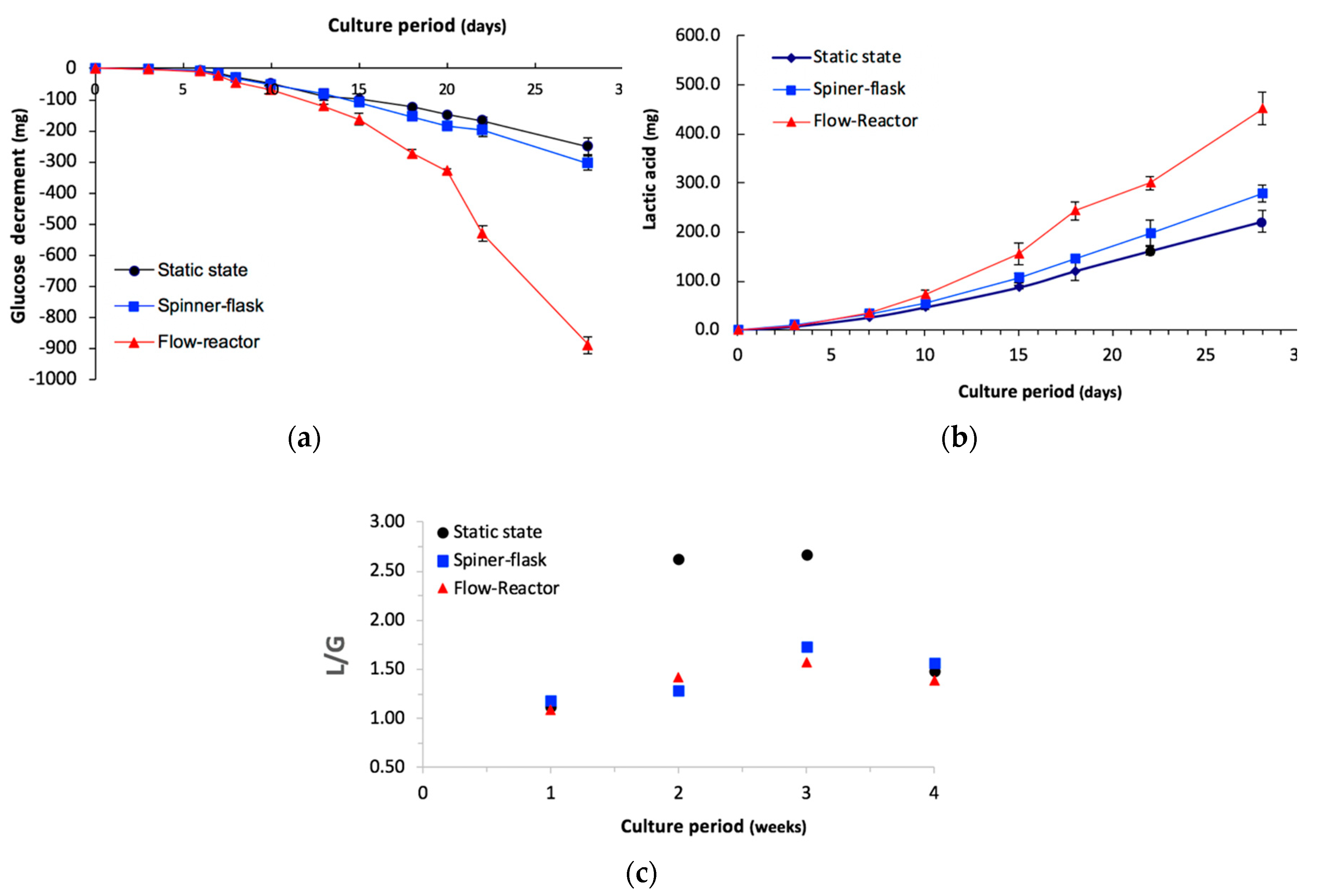
2.3. Metabolite Assay
2.4. Examination of Osteogenic Differentiation
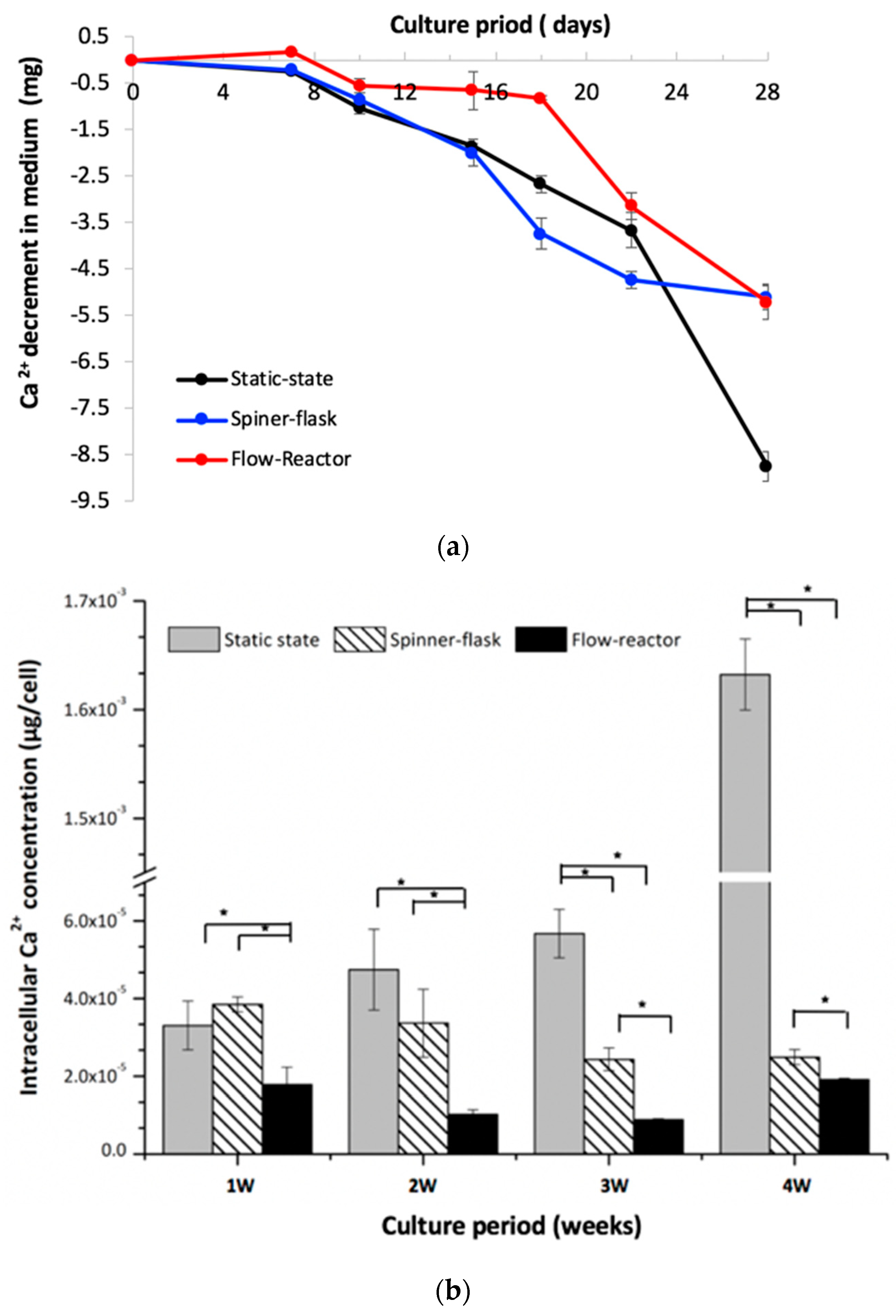
2.5. Medium and Intracellular Calcium Content
2.6. Histological Inspection
3. Discussion
4. Materials and Methods
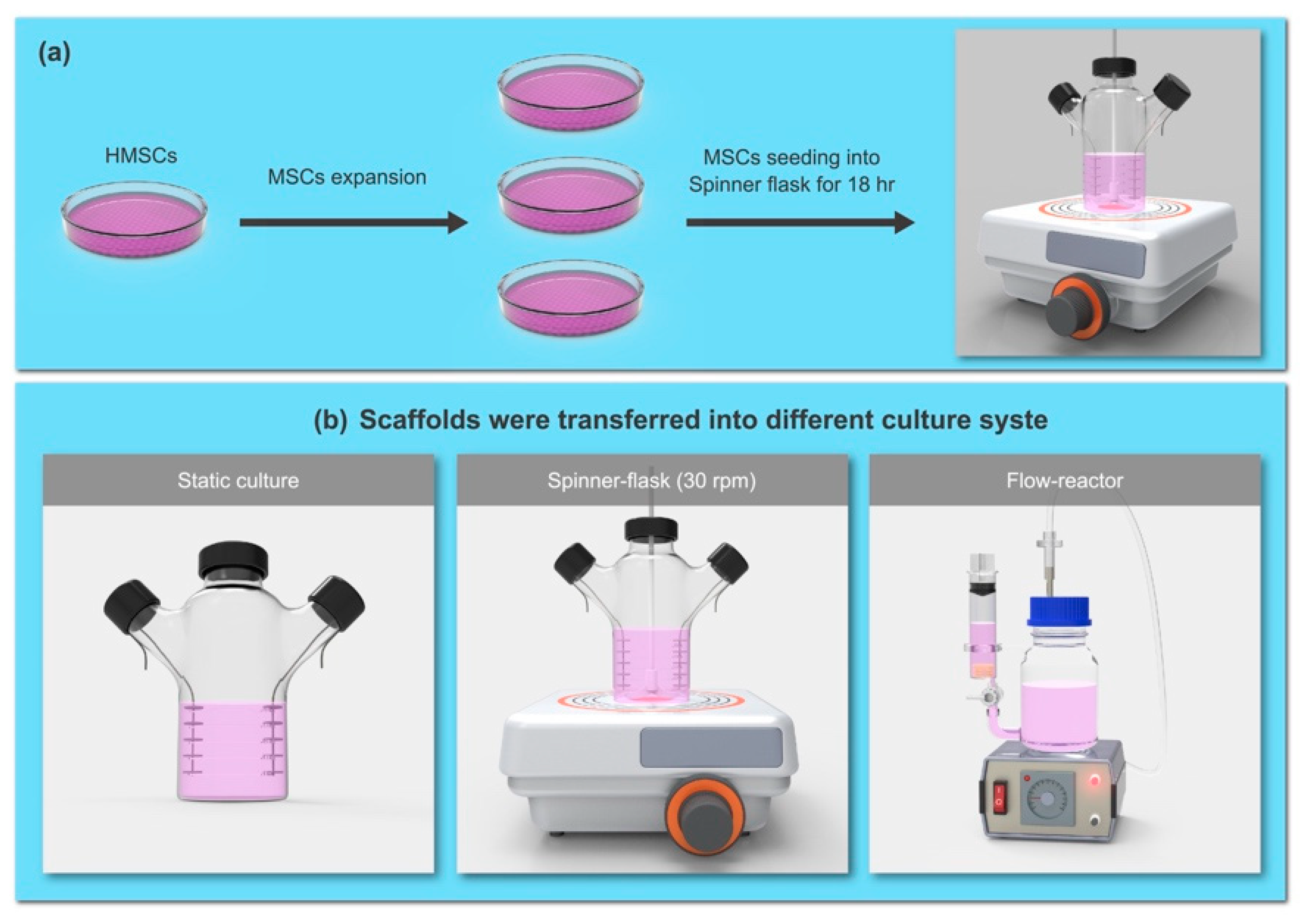
4.1. Cultivation of Human Bone Marrow MSCs in Bioreactors
4.2. Total DNA Quantification for Cell Proliferation
4.3. Mitochondrial Activity
4.4. Metabolite Assay
4.5. Examination of Osteogenic Differentiation
4.6. Histological Inspection
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heathman, T.R.; Nienow, A.W.; McCall, M.J.; Coopman, K.; Kara, B.; Hewitt, C.J. The translation of cell-based therapies: Clinical landscape and manufacturing challenges. Regen. Med. 2015, 10, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.R.; Chang, J.H.; Yang, K.C.; Lu, H.K.; Senatov, F.S.; Wu, C.C.; Tsai, M.H. The influence of vancomycin on extracellular matrix and pro-inflammatory cytokine expressions to human articular chondrocytes. Process Biochem. 2018, 65, 178–185. [Google Scholar] [CrossRef]
- Yang, S.H.; Hu, M.H.; Lo, W.Y.; Sun, Y.H.; Lin, F.H.; Yang, K.C. The influence of oxygen concentration on the extracellular matrix production of human nucleus pulposus cells during isolation-expansion process. J. Biomed. Mater. Res. A 2017, 105, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Bara, J.J.; Richards, R.G.; Alini, M.; Stoddart, M.J. Concise review: Bone marrow-derived mesenchymal stem cells change phenotype following in vitro culture: Implications for basic research and the clinic. Stem Cells 2014, 32, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Estes, B.T.; Diekman, B.O.; Guilak, F. Monolayer cell expansion conditions affect the chondrogenic potential of adipose-derived stem cells. Biotechnol. Bioeng. 2008, 99, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Tsai, A.C.; Liu, Y. Biomanufacturing of human mesenchymal stem cells in cell therapy: Influence of microenvironment on scalable expansion in bioreactors. Biochem. Eng. J. 2016, 108, 44–50. [Google Scholar] [CrossRef]
- Chen, X.; Xu, H.; Wan, C.; McCaigue, M.; Li, G. Bioreactor expansion of human adult bone marrow-derived mesenchymal stem cells. Stem Cells 2006, 24, 2052–2059. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tang, Y.; Xu, Y.; Xiao, Z.; Zhao, Y.; Li, J.; Han, S.; Chen, L.; Dai, B.; Wang, L.; Chen, B.; et al. The combination of three-dimensional and rotary cell culture system promotes the proliferation and maintains the differentiation potential of rat BMSCs. Sci. Rep. 2017, 7, 192. [Google Scholar] [CrossRef]
- Wang, T.W.; Wu, H.C.; Wang, H.Y.; Lin, F.H.; Sun, J.S. Regulation of adult human mesenchymal stem cells into osteogenic and chondrogenic lineages by different bioreactor systems. J Biomed. Mater. Res. A 2009, 88, 935–946. [Google Scholar] [CrossRef]
- King, J.A.; Mille, W.M. Bioreactor development for stem cell expansion and controlled differentiation. Curr. Opin. Chem. Biol. 2007, 11, 394–398. [Google Scholar] [CrossRef]
- Dos Santos, F.; Campbell, A.; Fernandes-Platzgummer, A.; Andrade, P.Z.; Gimble, J.M.; Wen, Y.; Boucher, S.; Vemuri, M.C.; da Silva, C.L.; Cabral, J.M. A xenogeneic-free bioreactor system for the clinical-scale expansion of human mesenchymal stem/stromal cells. Biotechnol. Bioeng. 2014, 111, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Côme, J.; Nissan, X.; Aubry, L.; Tournois, J.; Girard, M.; Perrier, A.L.; Peschanski, M.; Cailleret, M. Improvement of culture conditions of human embryoid bodies using a controlled perfused and dialyzed bioreactor system. Tissue Eng. Part. C Methods 2008, 14, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Stiehler, M.; Bünger, C.B.; Baatrup, A.; Lind, M.; Kassem, M.; Mygind, T. Effect of dynamic 3-D culture on proliferation, distribution, and osteogenic differentiation of human mesenchymal stem cells. J. Biomed. Mater. Res. A 2009, 89, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, W.; Han, B.; Xu, R. The bioreactor: A powerful tool for large-scale culture of animal cells. Curr. Pharm. Biotechnol. 2005, 6, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Tanzeglock, T.; Soos, M.; Stephanopoulos, G.; Morbidelli, M. Induction of mammalian cell death by simple shear and extensional flows. Biotechnol. Bioeng. 2009, 104, 360–370. [Google Scholar] [CrossRef]
- Walls, P.L.L.; McRae, O.; Natarajan, V.; Johnson, C.; Antoniou, C.; Bird, J.C. Quantifying the potential for bursting bubbles to damage suspended cells. Sci. Rep. 2017, 7, 15102. [Google Scholar] [CrossRef]
- Liu, Y.; Li, F.; Hu, W.; Wiltberger, K.; Ryll, T. Effects of bubble–liquid two-phase turbulent hydrodynamics on cell damage in sparged bioreactor. Biotechnol. Prog. 2014, 30, 48–58. [Google Scholar] [CrossRef]
- Sorg, R.; Tanzeglock, T.; Soos, M.; Morbidelli, M.; Périlleux, A.; Solacroup, T.; Broly, H. Minimizing hydrodynamic stress in mammalian cell culture through the lobed Taylor-Couette bioreactor. Biotechnol. J. 2011, 6, 1504–1515. [Google Scholar] [CrossRef]
- Lock, L.T.; Tzanakakis, E.S. Expansion and differentiation of human embryonic stem cellsto endoderm progeny in a microcarrier stirred-suspension culture. Tissue Eng. Part A 2009, 15, 2051–2063. [Google Scholar] [CrossRef]
- Chen, A.K.L.; Reuveny, S.; Oh, S.K.W. Application of human mesenchymal and pluripotent stem cell microcarrier cultures in cellular therapy: Achievements and future direction. Biotechnol. Adv. 2013, 15, 1032–1046. [Google Scholar] [CrossRef]
- Strathearn, K.E.; Pardo, A.M.P. Parameters to Consider When Expanding Cells on Corning® Microcarriers. Corning Application Note CLS-AN-243. Available online: https://www.corning.com/worldwide/en/products/life-sciences/ products/bioprocess/microcarriers.html. (accessed on 17 August 2019).
- Osiecki, M.J.; McElwain, S.D.L.; Lott, W.B. Modelling mesenchymal stromal cell growth in a packed bed bioreactor with a gas permeable wall. PLoS ONE 2018, 13, e0202079. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.K.; Zhang, Z.Y.; Chen, A.K.; Reuveny, S.; Choolani, M.; Chan, J.K.; Oh, S.K. Microcarrier culture for efficient expansion and osteogenic differentiation of human fetal mesenchymal stem cells. Biores Open Access 2013, 2, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.C.; Chen, Y.J.; Yew, T.L.; Chen, L.L.; Wang, J.Y.; Chiu, C.H.; Hung, S.C. Hypoxia inhibits senescence and maintains mesenchymal stem cell properties through down-regulation of E2A-p21 by HIF-TWIST. Blood 2011, 117, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.S.; Ryu, H.W. Lactic acid production and carbon catabolite repression from single and mixed sugars using Enterococcus faecalis RKY1. Process Biochem. 2001, 37, 235–240. [Google Scholar] [CrossRef]
- Nishimura, I.; Hisanaga, R.; Sato, T.; Arano, T.; Nomoto, S.; Ikada, Y.; Yoshinari, M. Effect of osteogenic differentiation medium on proliferation and differentiation of human mesenchymal stem cells in three-dimensional culture with radial flow bioreactor. Regenerative Ther. 2015, 2, 24–31. [Google Scholar] [CrossRef]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon Med. Sch. 2010, 77, 4–12. [Google Scholar] [CrossRef]
- Komori, T. Regulation of osteoblast differentiation by Runx2. Adv. Exp. Med. Biol. 2010, 658, 43–49. [Google Scholar]
- Bakker, A.D.; Soejima, K.; Klein-Nulend, J.; Burger, E.H. The production of nitric oxide and prostaglandin E(2) by primary bone cells is shear stress dependent. J. Biomech. 2001, 34, 671–677. [Google Scholar] [CrossRef]
- Li, D.; Tang, T.; Lu, J.; Dai, K. Effects of flow shear stress and mass transport on the construction of a large-scale tissue engineered bone in a perfusion bioreactor. Tissue Eng Part. A 2009, 15, 2773–2783. [Google Scholar] [CrossRef]
- Birru, B.; Mekala, N.K.; Parcha, S.R. Improved osteogenic differentiation of umbilical cord blood MSCs using custom made perfusion bioreactor. Biomed. J. 2018, 41, 290–297. [Google Scholar] [CrossRef]
- Tseng, C.L.; Chang, K.M.; Wu, C.C.; Wang, Y.K.; Wang, I.K. Rabbit mesenchymal stem cells cultured in a dynamic culture system displayed superior cell proliferation and osteogenetic induction. J. Exp. Clin. Med. 2014, 6, 10–15. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Period | Static State | Spinner Flask | Flow Reactor |
|---|---|---|---|
| 1 week | 1.08 ± 0.17 | 1.00 ± 0.19 | 2.70 ± 0.04 *,# |
| 2 weeks | 1.27 ± 0.10 | 1.65 ± 0.10* | 4.64 ± 0.36 *,# |
| 3 weeks | 1.51 ± 0.02 | 2.68 ± 0.64* | 6.08 ± 0.21 *,# |
| 4 weeks | 1.27 ± 0.02 | 3.13 ± 0.23* | 5.85 ± 0.02 *,# |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, H.-H.; Yang, K.-C.; Wu, M.-H.; Chen, J.-C.; Tseng, C.-L. The Effects of Different Dynamic Culture Systems on Cell Proliferation and Osteogenic Differentiation in Human Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 4024. https://doi.org/10.3390/ijms20164024
Tsai H-H, Yang K-C, Wu M-H, Chen J-C, Tseng C-L. The Effects of Different Dynamic Culture Systems on Cell Proliferation and Osteogenic Differentiation in Human Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2019; 20(16):4024. https://doi.org/10.3390/ijms20164024
Chicago/Turabian StyleTsai, Hsiou-Hsin, Kai-Chiang Yang, Meng-Huang Wu, Jung-Chih Chen, and Ching-Li Tseng. 2019. "The Effects of Different Dynamic Culture Systems on Cell Proliferation and Osteogenic Differentiation in Human Mesenchymal Stem Cells" International Journal of Molecular Sciences 20, no. 16: 4024. https://doi.org/10.3390/ijms20164024
APA StyleTsai, H.-H., Yang, K.-C., Wu, M.-H., Chen, J.-C., & Tseng, C.-L. (2019). The Effects of Different Dynamic Culture Systems on Cell Proliferation and Osteogenic Differentiation in Human Mesenchymal Stem Cells. International Journal of Molecular Sciences, 20(16), 4024. https://doi.org/10.3390/ijms20164024