Enhanced Hepatogenic Differentiation of Human Wharton’s Jelly–Derived Mesenchymal Stem Cells by Using Three-Step Protocol


Abstract
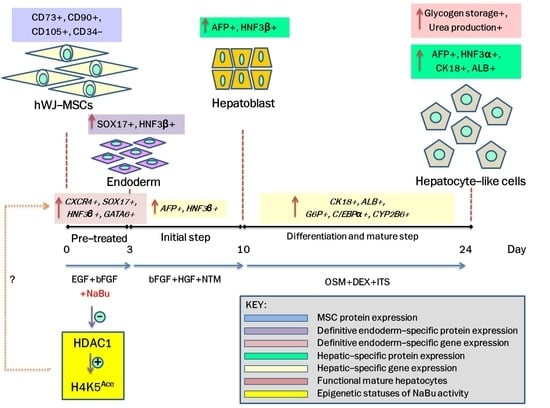
:
1. Introduction
2. Results
2.1. Isolation and Characterization of hWJ-MSCs
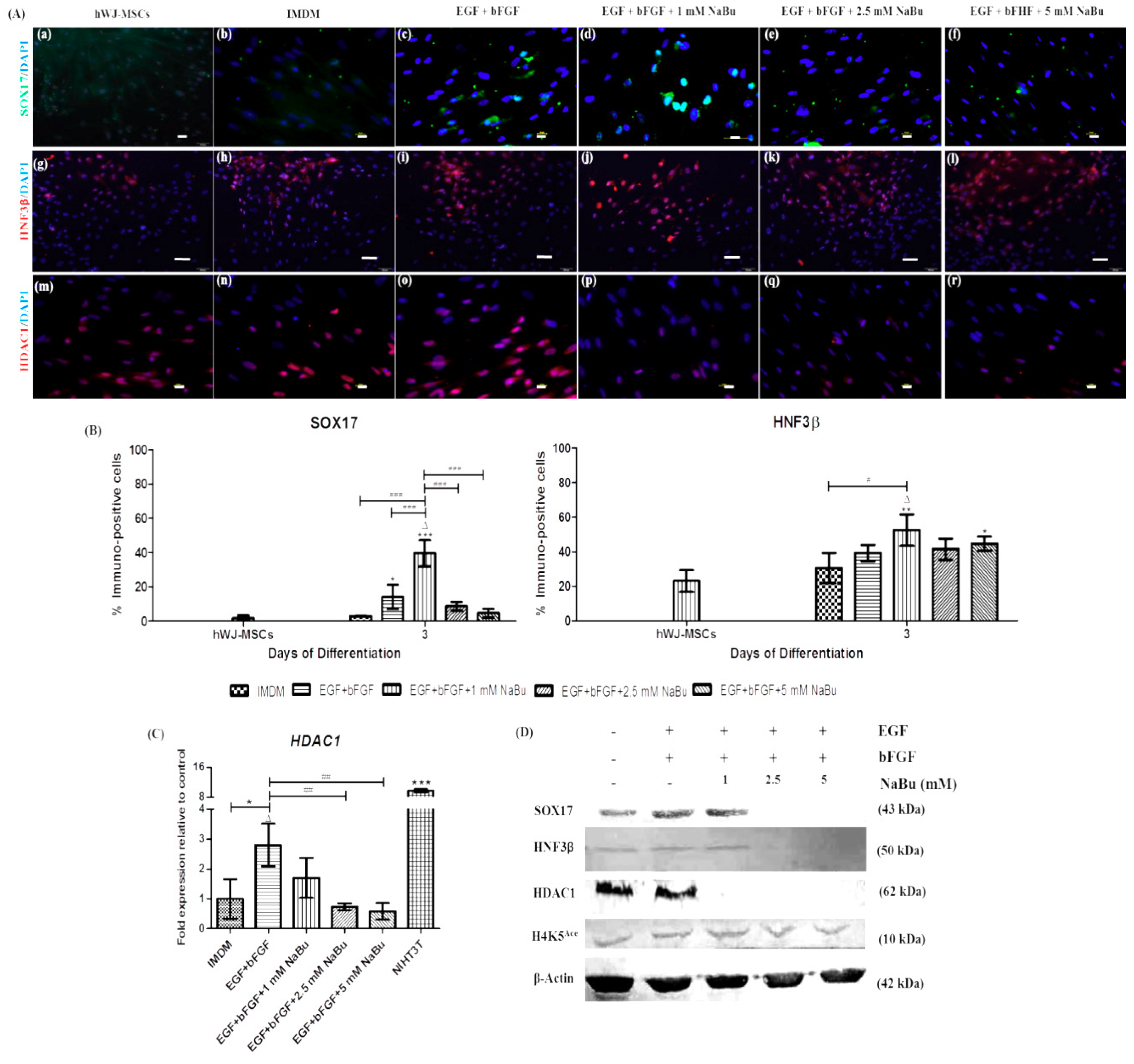
2.2. Effect of NaBu Treatment on hWJ-MSC Viability
2.3. Effect of NaBu on Epigenetic Statuses and Endodermal Differentiation of hWJ-MSCs
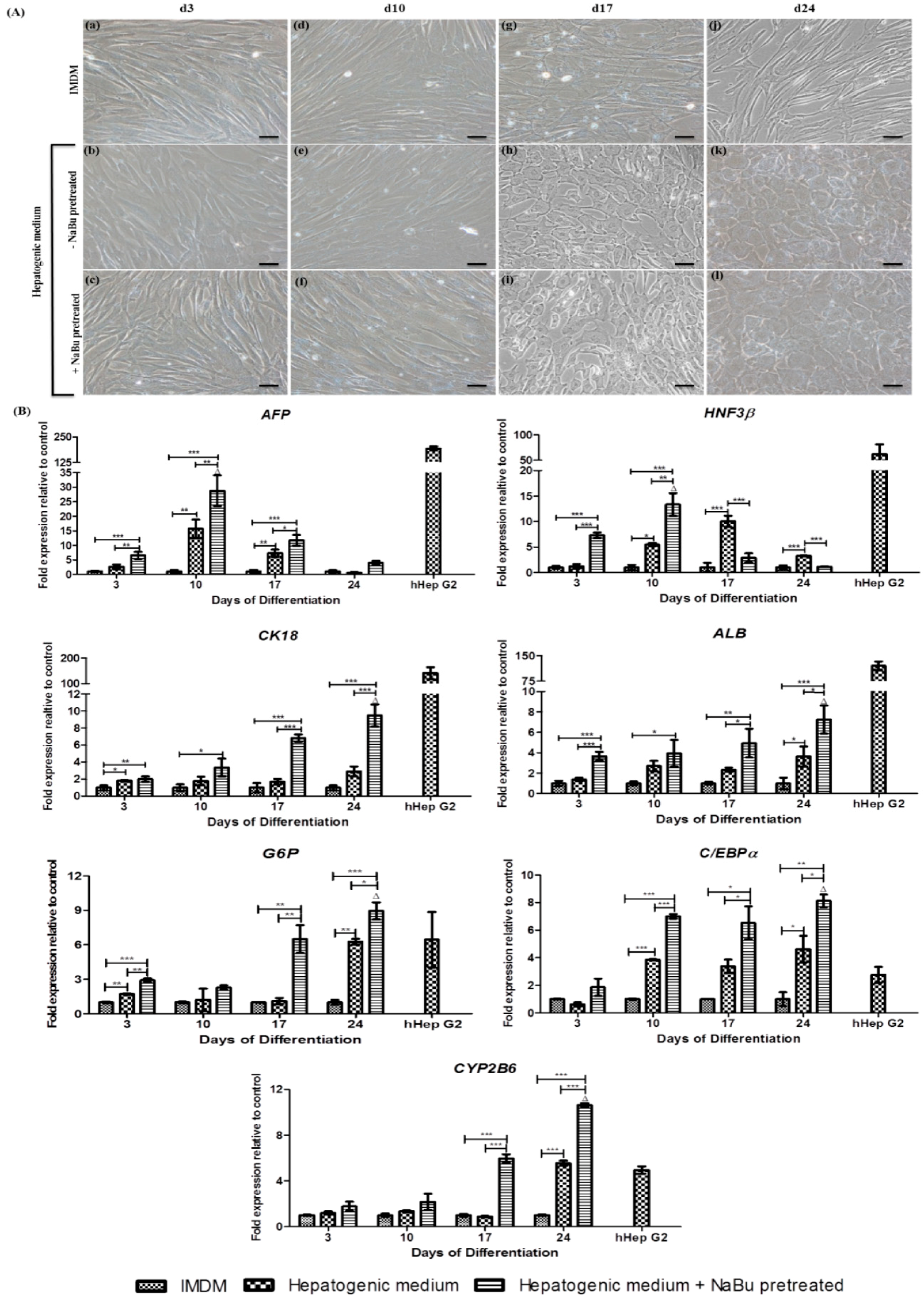
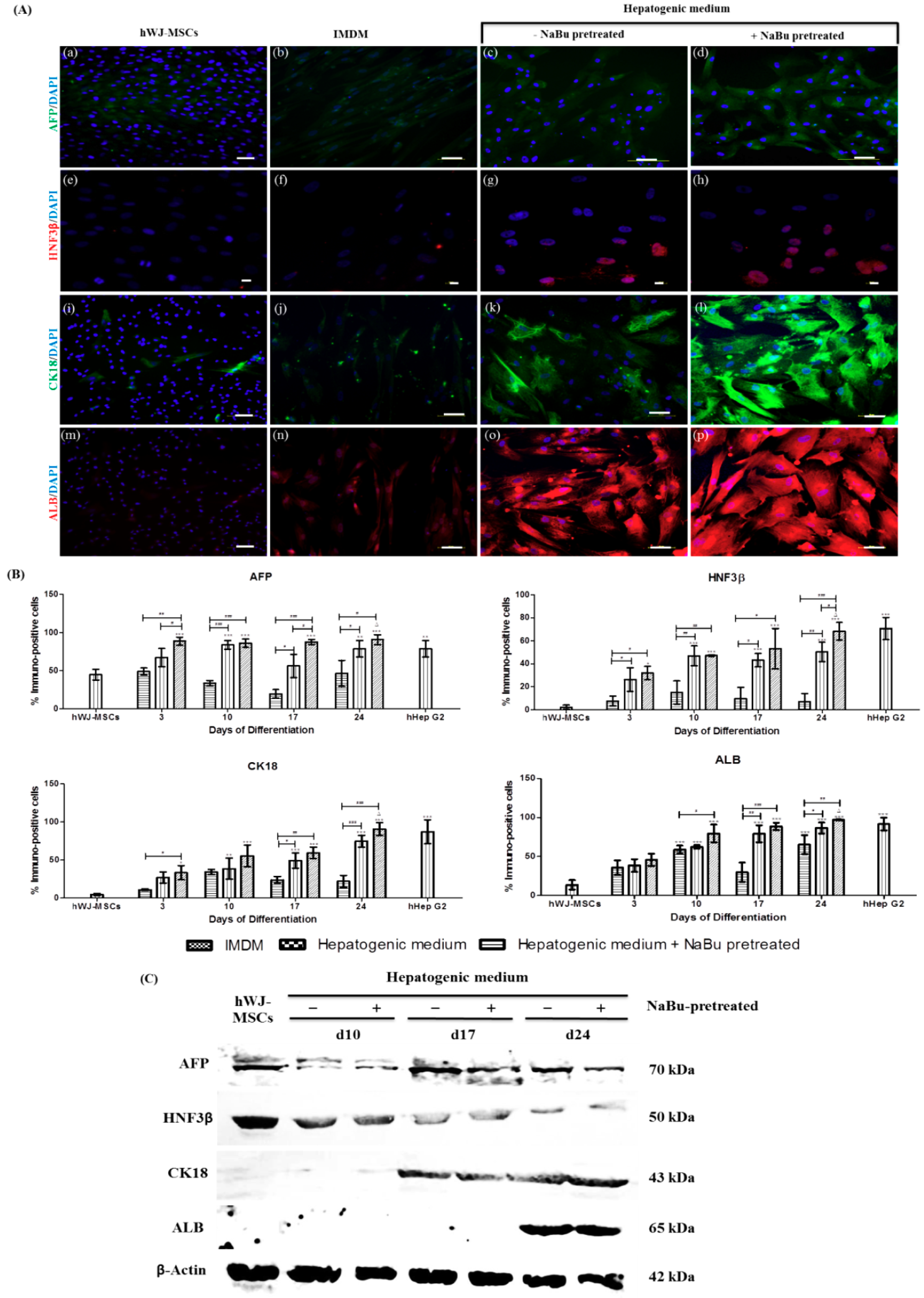
2.4. Effect of NaBu on Hepatogenic Differentiation of hWJ-MSCs
3. Discussion
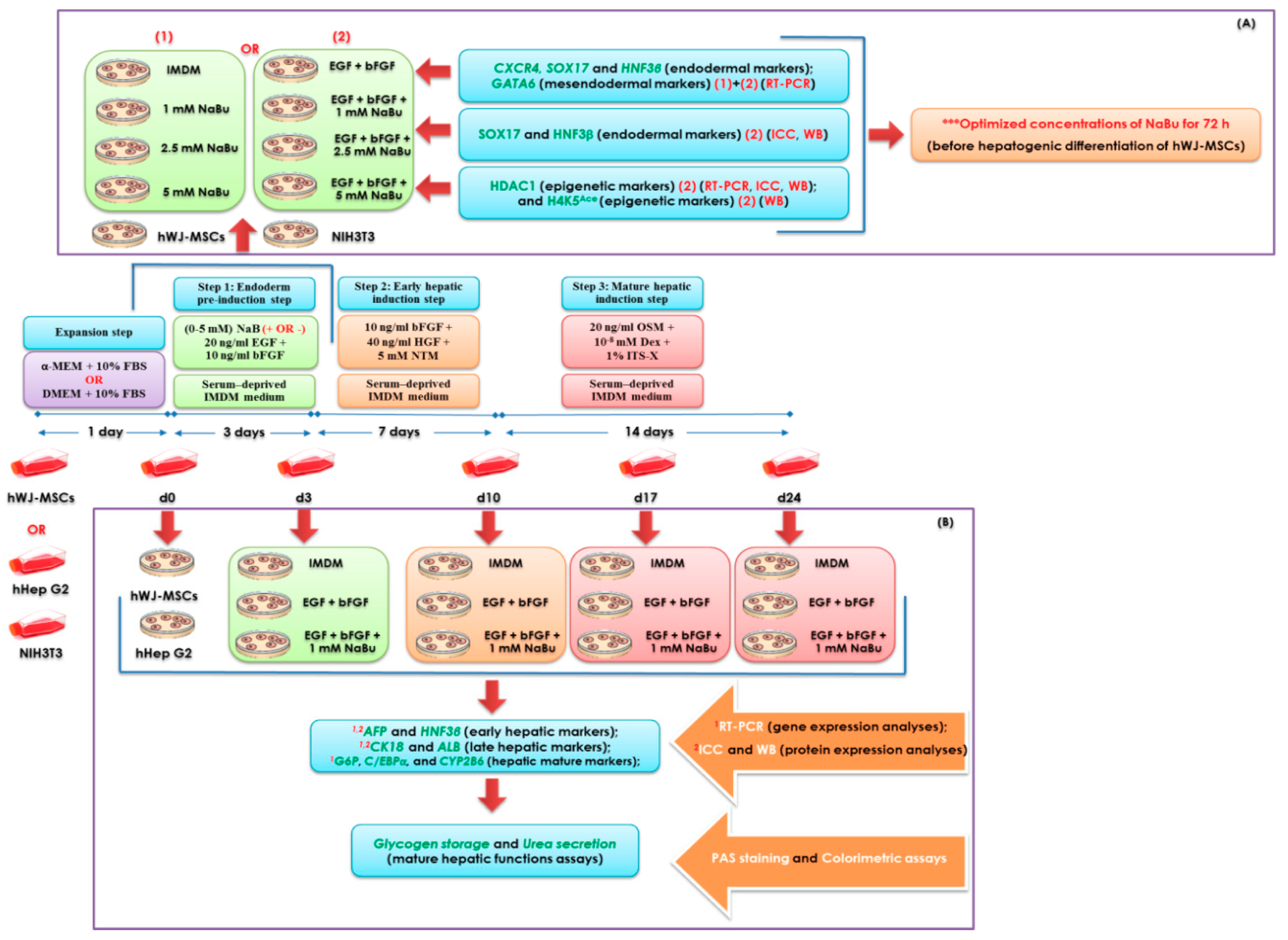
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Human Hepatocarcinoma (hHep G2) and NIH3T3 Cell Culture
4.3. Isolation and Culture of hWJ-MSCs
4.4. Characterizations of hWJ-MSCs
4.5. Analysis of Cytotoxicity
4.6. Treatment with NaBu
4.7. Hepatogenic Differentiation
4.8. Gene Expression Analysis
4.9. Immunofluorescence Staining
4.10. SDS-PAGE and Western Blot Analysis
4.11. Periodic Acid-Schiff Staining
4.12. Analysis of Urea Production
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MSCs | Mesenchymal stem cells |
| ESCs | Embryonic stem cells |
| hWJ-MSCs | Human Wharton’s jelly-derived mesenchymal stem cells |
| NaBu | Sodium butyrate |
| EGF | Epidermal growth factor |
| bFGF | Basic fibroblast growth factor |
| HDACi | Histone deactylase inhibitor |
| HGF | Hepatocyte growth factor |
| OSM | Oncostatin M |
| ITS | Insulin-transferrin-selenium-ethanolamine |
| hHepG2 | Human hepatocarcinoma cell line |
| DMEM | Dulbecco’s modified Eagle’s medium |
| FBS | Fetal bovine serum |
| α-MEM | Alpha-Minimum Essential medium |
| IMDM | Iscove’s modified Dulbecco’s medium |
| DMSO | Dimethyl sulfoxide |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide |
| H4K5Ace | Acetylated histone H4 (Lys5) |
| RT-PCR | Real-time reverse transcriptase-polymerase chain reaction |
| PFA | Paraformaldehyde |
| BSA | Bovine serum albumin |
References
- Gordillo, M.; Evans, T.; Gouon-Evans, V. Orchestrating liver development. Development 2015, 142, 2094–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaret, K.S.; Grompe, M. Generation and regeneration of cells of the liver and pancreas. Science 2008, 322, 1490–1494. [Google Scholar] [CrossRef]
- Dhawan, A.; Puppi, J.; Hughes, R.D.; Mitry, R.R. Human hepatocyte transplantation: Current experience and future challenges. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marin, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Romanov, Y.A.; Darevskaya, A.N.; Merzlikina, N.V.; Buravkova, B. Mesenchymal stem cells from human bone marrow and adipose tissue: Isolation, characterization and differentiation potentialities. Bull. Exp. Biol. Med. 2005, 140, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Troyer, D.L.; Weiss, M.L. Wharton’s jelly-derived cells are a primitive stromal cell population. Stem Cells 2008, 26, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Witkowska-Zimny, M.; Wrobel, E. Perinatal sources of mesenchymal stem cells: Wharton’s jelly, amnion and chorion. Cell. Mol. Biol. Lett. 2011, 16, 493–514. [Google Scholar] [CrossRef] [PubMed]
- Karahuseyinoglu, S.; Cinar, O.; Kilic, E.; Kara, F.; Akay, G.G.; Demiralp, D.Ö.; Tukun, A.; Uckan, D.; Can, A. 2007. Biology of stem cells in human umbilical cord stroma: In situ and in vitro surveys. Stem Cells 2007, 25, 319–331. [Google Scholar] [CrossRef]
- Wang, H.S.; Hung, S.C.; Peng, S.T.; Huang, C.C.; Wei, H.M.; Guo, Y.J.; Fu, Y.S.; Lai, M.C.; Chen, C.C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar] [CrossRef]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E.M. Native umbilical cord matrix stem cells express hepatic markers and differentiate into hepatocyte-like cells. Gastroenterol 2008, 134, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Lie, P.C.; Wei, X. Differentiation of mesenchymal stromal cells derived from umbilical cord Wharton’s jelly into hepatocyte-like cells. Cytotherapy 2009, 11, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Ren, H.; Li, X.; Chen, Z.; Zhang, X.; Gong, W.; Liu, Y.; Pang, T.; Han, Z.C. Differentiation of human umbilical cord mesenchymal stromal cells into low immunogenic hepatocyte-like cells. Cytotherapy 2009, 11, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.H.; Jung, B.Y.; Seo, Y.K.; Song, K.Y.; Park, J.K. In vitro hepatic differentiation of umbilical cord-derived mesenchymal stem cell. Proc. Chem. 2010, 45, 1857–1864. [Google Scholar] [CrossRef]
- Prasajak, P.; Leeanansaksiri, W. Developing a new two-step protocol to generate functional hepatocytes from Wharton’s jelly-derived mesenchymal stem cells under hypoxic condition. Stem Cells Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed]
- An, S.Y.; Han, J.; Lim, H.J.; Park, S.Y.; Kim, J.H.; Do, B.R.; Kim, J.H. Valproic acid promotes differentiation of hepatocyte-like cells from whole human umbilical cord-derived mesenchymal stem cells. Tissue Cell 2014, 46, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.K.; Perrine, S.P.; Williams, R.M.; Faller, D.V. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents. Blood 2012, 119, 1008–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Ghazawi, F.M.; Bakkar, W.; Li, Q. Valproic acid and butyrate induce apoptosis in human cancer cells through inhibition of gene expression of Akt/protein kinase B. Mol. Cancer 2006, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Vrba, J.; Trtkova, K.; Ulrichova, J. HDAC inhibitors sodium butyrate and sodium valproate do not affect human ncor1 and ncor2 gene expression in HL-60 cells. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2011, 155, 259–262. [Google Scholar] [CrossRef]
- Hay, D.C.; Zhao, D.; Fletcher, J.; Hewitt, Z.A.; McLean, D.; Urruticoechea-Uriguen, A.; Black, J.R.; Elcombe, C.; Ross, J.A.; Wolf, R.; et al. Efficient differentiation of hepatocytes from human embryonic stem cells exhibiting markers recapitulating liver development in vivo. Stem Cells 2008, 26, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Shang, C.Z.; Lu, L.H.; Qiu, D.C.; Ren, M.; Chen, Y.J.; Min, J. Differentiation of embryonic stem cells into hepatocytes that coexpress coagulation factors VIII and IX. Acta Pharmacol. Sin. 2010, 31, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Li, P.; Tan, L.; Qu, S.; Ying, Q.L.; Song, H. Differentiation of mouse embryonic stem cells into hepatocytes induced by a combination of cytokines and sodium butyrate. J. Cell. Biochem. 2010, 109, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Yan, L.; Shang, Z.C.; Cao, J.; Lu, L.H.; Min, J.; Cheng, H. Effects of sodium butyrate on the differentiation of pancreatic and hepatic progenitor cells from mouse embryonic stem cells. J. Cell. Biochem. 2010, 109, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zhong, B.; Qi, W.; Song, Y.; Wu, Y.; Fan, Y.; Wang, F. In vitro differentiation of mouse embryonic stem cells into functional hepatocytes by sodium butyrate, hepatocyte growth factor and dexamethasone under chemically defined conditions. Afr. J. Biotechnol. 2011, 10, 9493–9500. [Google Scholar] [Green Version]
- Tanthaisong, P.; Imsoonthornruksa, S.; Ngernsoungnern, A.; Ngernsoungnern, P.; Ketudat-Cairns, M.; Parnpai, R. Enhanced chondrogenic differentiation of human umbilical cord Wharton’s jelly derived mesenchymal stem cells by GSK-3 inhibitors. PLoS ONE 2017, 12, e0168059. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, F.S. Stem cell therapies for liver failures and cirrhosis. J. Hepatol. 2013, 59, 183–185. [Google Scholar] [CrossRef]
- Chen, Y.; Pan, R.L.; Zhang, X.L.; Shao, J.Z.; Xiang, L.X.; Dong, X.J.; Zhang, G.O. Induction of hepatic differentiation of mouse bone marrow stromal stem cells by the histone deacetylase inhibitor VPA. J. Cell. Mol. Med. 2009, 13, 2582–2592. [Google Scholar] [CrossRef]
- Dong, X.; Pan, R.; Zhang, H.; Yang, C.; Shao, J.; Xiang, L. Modification of histone acetylation facilitates hepatic differentiation of human bone marrow mesenchymal stem cells. PLoS ONE 2013, 8, e63405. [Google Scholar] [CrossRef]
- Lee, S.; Park, J.R.; Seo, M.S.; Roh, K.H.; Park, S.B.; Hwang, J.W.; Sun, B.; Seo, K.; Lee, Y.S.; Kang, S.K.; et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009, 42, 711–720. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Wu, Y.; Ni, B.; Liang, Z. Sodium butyrate promotes the differentiation of rat bone marrow mesenchymal stem cells to smooth muscle cells through histone acetylation. PLoS ONE 2014, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Fu, Y.; Zhang, X.; Dai, L.; Zhu, J.; Bi, Z.; Ao, Y.; Zhou, C. Histone deacetylase inhibitor sodium butyrate promotes the osteogenic differentiation of rat adipose-derived stem cells. Dev. Growth Differ. 2014, 56, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.D.; Kuo, T.K.; Whang-Peng, J.; Chung, Y.F.; Lin, C.T.; Chou, S.H.; Chen, J.R.; Chen, Y.P.; Lee, O.K. In vitro hepatic differentiation of human mesenchymal stem cells. Hepatology 2004, 40, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Talèns-Visconti, R.; Bonora, A.; Jover, R.; Mirabet, V.; Carbonell, F.; Castell, J.V.; Gómez-Lechón, M.J. Hepatogenic differentiation of human mesenchymal stem cells from adipose tissue in comparison with bone marrow mesenchymal stem cells. World J. Gastroenterol. 2006, 12, 5834–5845. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Zheng, M.; Goldfarb, M.; Zaret, K.S. Initiation of mammalian liver development from endoderm by fibroblast growth factors. Science 1999, 284, 1998–2003. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Sekiguchi, T.; Xu, M.J.; Ito, Y.; Kamiya, A.; Tsuji, K. Hepatic differentiation induced by oncostatin M attenuates fetal liver hematopoiesis. Proc. Natl. Acad. Sci. USA 1999, 96, 7265–7270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamiya, A.; Kinoshita, T.; Miyajima, A. Oncostatin M and hepatocyte growth factor induce hepatic maturation via distinct signaling pathways. FEBS Lett. 2001, 492, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zheng, F. Directed hepatic differentiation from embryonic stem cells. Protein Cell 2011, 2, 180–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, F.; Mitaka, T.; Mizuguchi, T.; Mochizuki, Y.; Hirata, K. Effects of nicotinamide related agents on the growth of primary rat hepatocytes and formation of small hepatocyte colonies. Liver 1999, 19, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, J.S.; Liu, F.; Omiecinski, C.J. Phenobarbital responsiveness as a uniquely sensitive indicator of hepatocyte differentiation status: Requirement of dexamethasone and extracellular matrix in establishing the functional integrity of cultured primary rat hepatocytes. Exp. Cell Res. 2004, 292, 252–264. [Google Scholar] [CrossRef]
- Allahbakhshi, E.; Hashemitabar, M.; Shariati, M.; Tabandeh, M.R.; Solgi, G. Differentiation of the definitive endoderm from Wharton’s Jelly Mesenchymal Stem Cells (WJMSC). J. Biol. Res. 2013, 20, 217–227. [Google Scholar]
- Schwartz, R.E.; Reyes, M.; Koodie, L.; Jiang, Y.; Blackstad, M.; Lund, T.; Lenvik, T.; Johnson, S.; Hu, W.S.; Verfaillie, C.M. Multipotent adult progenitor cells from bone marrow differentiate into functional hepatocyte-like cells. J. Clin. Investig. 2002, 109, 1291–1302. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, X.; Yao, L.; Lehman, D.M.; Wang, P. Improvement of cell survival during human pluripotent stem cell definitive endoderm differentiation. Stem Cells Dev. 2015, 24, 2536–2546. [Google Scholar] [CrossRef] [PubMed]
- Si-Tayeb, K.; Lemaigre, F.P.; Duncan, S.A. Organogenesis and development of the liver. Dev. Cell 2010, 18, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Kaestner, K.H. The FoxA factors in organogenesis and differentiation. Curr. Opin. Genet. Dev. 2010, 20, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacini, S.; Fazzi, R.; Montali, M.; Carnicelli, V.; Lazzarini, E.; Petrini, M. Specific integrin expression is associated with podosome-like structures on mesodermal progenitor cells. Stem Cells Dev. 2013, 22, 1830–1838. [Google Scholar] [CrossRef] [PubMed]
- King, C.C.; Beattie, G.M.; Lopez, A.D.; Hayek, A. Generation of definitive endoderm from human embryonic stem cells cultured in feeder layer-free conditions. Reg. Med. 2008, 3, 175–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, D.S.; Alawathugoda, T.T.; Ansari, S.A. Fine tuning of hepatocyte differentiation from human embryonic stem cells: Growth factors vs. small molecule based approaches. Stem Cell Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Zhou, F.; Zhang, Y.; Deng, R.; Xu, W.; Bai, M.; Liu, Y.; Shao, L.; Wang, X.; Zhou, L. Butyrate stimulates hepatic gluconeogenesis in mouse primary hepatocytes. Exp. Ther. Med. 2019, 7, 1677–1687. [Google Scholar] [CrossRef]
- Ye, D.; Li, T.; Heraud, P.; Parnpai, R. Effect of chromatin-remodeling agents in hepatic differentiation of rat bone marrow-derived mesenchymal stem cells in vitro and in vivo. Stem Cell Int. 2016. [Google Scholar] [CrossRef] [PubMed]
- Nekanti, U.; Mohanty, L.; Venugopal, P.; Balasubramanian, S.; Totey, S.; Ta, M. Optimization and scale-up of wharton’s jelly-derived mesenchymal stem cells for clinical applications. Stem Cell Res. 2010, 5, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.; Nagamoto, Y.; Mimura, N.; Tashiro, K.; Sakurai, F.; Tachibana, M.; Hayakawa, T.; Kawabata, K.; Mizuguchi, H. Long-term self-renewal of human ES/iPS-derived hepatoblast-like cells on human laminin 111-coated dishes. Stem Cell Rep. 2013, 1, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wang, J.; Jia, J.; Song, N.; Xiang, C.; Xu, J.; Hou, Z.; Su, X.; Liu, B.; Jiang, T.; et al. Human hepatocytes with drug metabolic function induced from fibroblasts by lineage reprogramming. Cell Stem Cell 2014, 14, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Najimi, M.; Khuu, D.N.; Lysy, P.A.; Jazouli, N.; Abarca, J.; Sempoux, C.; Sokal, E.M. Adult-derived human liver mesenchymal-like cells as a potential progenitor reservoir of hepatocytes? Cell Transplant. 2007, 16, 717–728. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′→3′) | Annealing Temperature(°C) | Product Size (bp) | References |
|---|---|---|---|---|
| SOX17 | F: GGCGCAGCAGAATCCAGA | 65 | 61 | [47] |
| R: CCACGACTTGCCCAGCAT | ||||
| CXCR4 | F: ACTACACCGAGGAAATGGGCT | 60 | 133 | NM_003467.2 This study |
| R: CCCACAATGCCAGTTAAGAAGA | ||||
| GATA6 | F: CCATGACTCCAACTTCCACC | 62 | 214 | [52] |
| R: ACGGAGGACGTGACTTCGGC | ||||
| AFP | F: CTTTGGGCTGCTCGCTATGA | 60 | 131 | NM_001134 This study |
| R: GCATGTTGATTTAACAAGCTGCT | ||||
| HNF3β | F: CCTACTCGTACATCTCGCTCATC | 65 | 69 | [42] |
| R: CGCTCAGCGTCAGCATCTT | ||||
| CK18 | F: TCGCAAATACTGTGGACAATGC | 60 | 171 | NM_199187.1 This study |
| R: GCAGTCGTGTGATATTGGTGT | ||||
| ALB | F: GCACAGAATCCTTGGTGAACAG | 65 | 101 | [53] |
| R: ATGGAAGGTGAATGTTTCAGCA | ||||
| G6P | F: TCAGCTCAGGTGGTCCTCTT | 62 | 291 | [54] |
| R: CCTCCTTAGGCAGCCTTCTT | ||||
| C/EBPα | F: ACAAGAACAGCAACGAGTACCG | 65 | 129 | [53] |
| R: CATTGTCACTGGTCAGCTCCA | ||||
| CYP2B6 | F: GTGATCTTTTGTGTCTGGTTGC | 62 | 138 | NM_000767.4 This study |
| R: GATAGACGGAAGCAGTAGGAAG | ||||
| HDAC1 | F: CCAAGTACCACAGTGATGACTACATT | 65 | 135 | NM_004964.2 This study |
| R: AGAACTCAAACAGGCCATCAAA | ||||
| GAPDH | F: TGCACCACCAACTGCTTAGC | 60 | 87 | [53] |
| R: GGCATGGACTGTGGTCATGAG | ||||
| β-ACTIN | F: TCACCACCACGGCCGAGCG | 60 | 350 | X00351 This study |
| R: TCTCCTTCTGCATCCTGTCG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panta, W.; Imsoonthornruksa, S.; Yoisungnern, T.; Suksaweang, S.; Ketudat-Cairns, M.; Parnpai, R. Enhanced Hepatogenic Differentiation of Human Wharton’s Jelly–Derived Mesenchymal Stem Cells by Using Three-Step Protocol. Int. J. Mol. Sci. 2019, 20, 3016. https://doi.org/10.3390/ijms20123016
Panta W, Imsoonthornruksa S, Yoisungnern T, Suksaweang S, Ketudat-Cairns M, Parnpai R. Enhanced Hepatogenic Differentiation of Human Wharton’s Jelly–Derived Mesenchymal Stem Cells by Using Three-Step Protocol. International Journal of Molecular Sciences. 2019; 20(12):3016. https://doi.org/10.3390/ijms20123016
Chicago/Turabian StylePanta, Wachira, Sumeth Imsoonthornruksa, Ton Yoisungnern, Sanong Suksaweang, Mariena Ketudat-Cairns, and Rangsun Parnpai. 2019. "Enhanced Hepatogenic Differentiation of Human Wharton’s Jelly–Derived Mesenchymal Stem Cells by Using Three-Step Protocol" International Journal of Molecular Sciences 20, no. 12: 3016. https://doi.org/10.3390/ijms20123016