Expression of Chemerin and Its Receptors in the Porcine Hypothalamus and Plasma Chemerin Levels during the Oestrous Cycle and Early Pregnancy

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
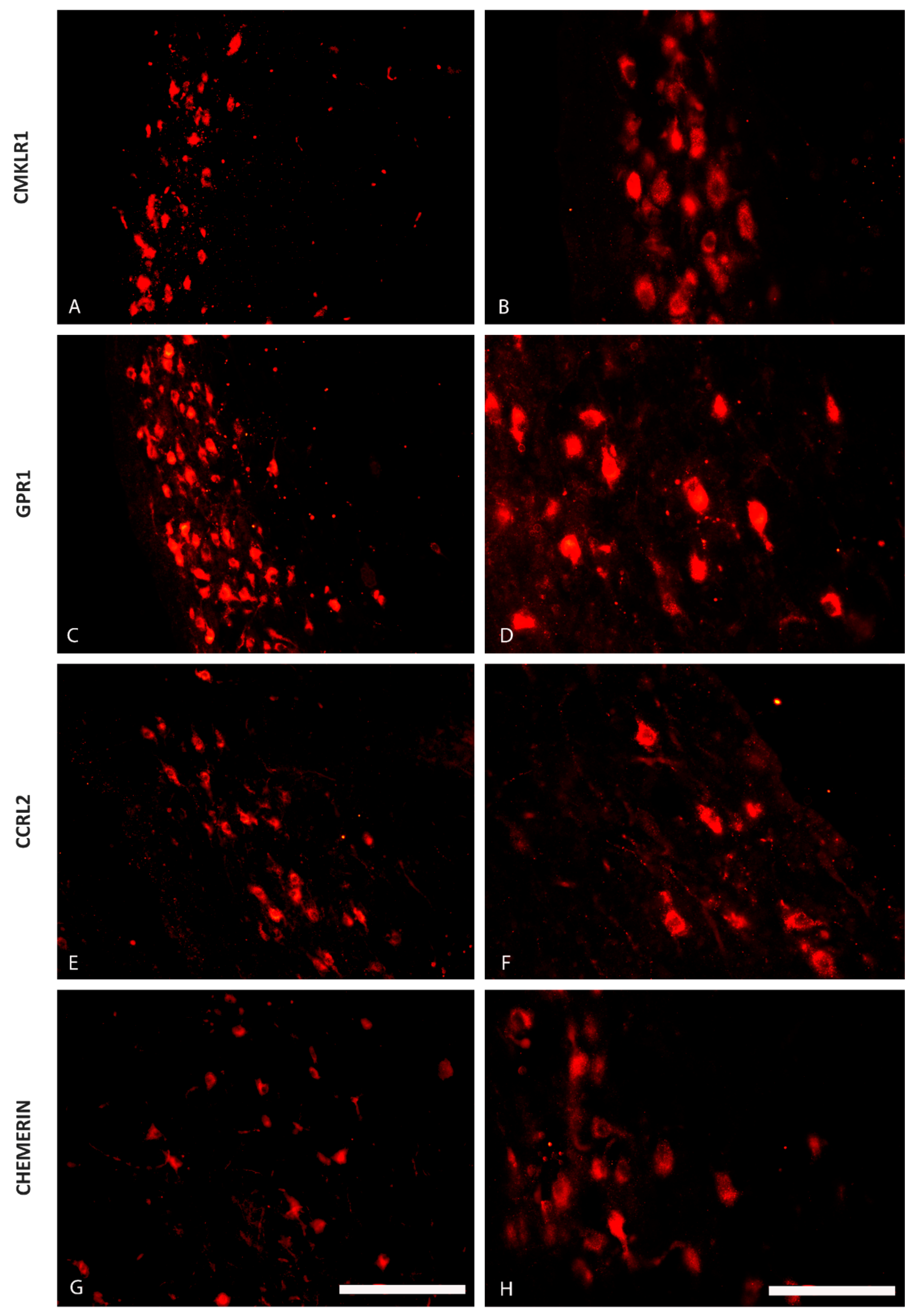
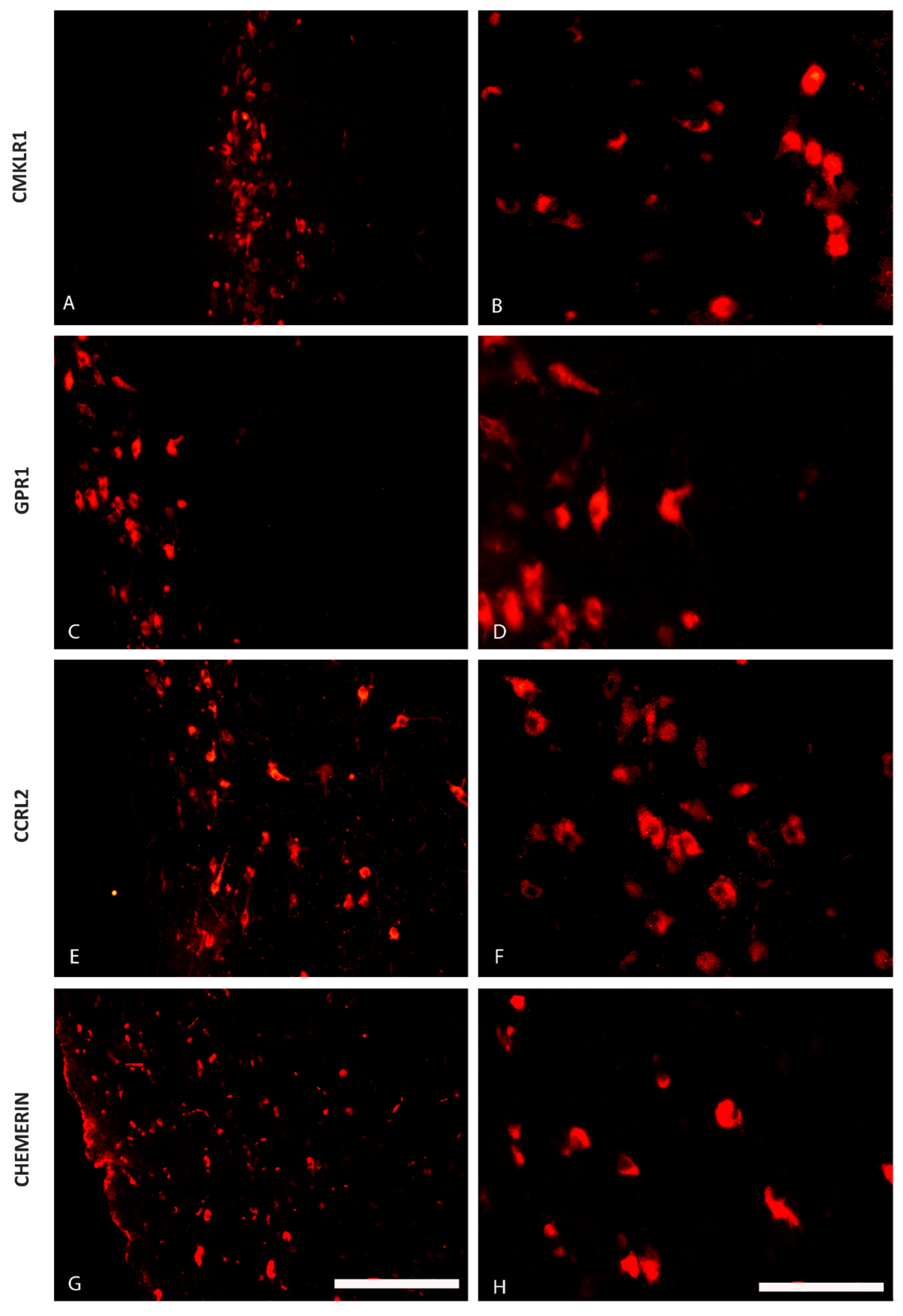
2.1. The Distribution of CHEM, CMKLR1, GPR1 and CCRL2 in the Porcine Hypothalamus
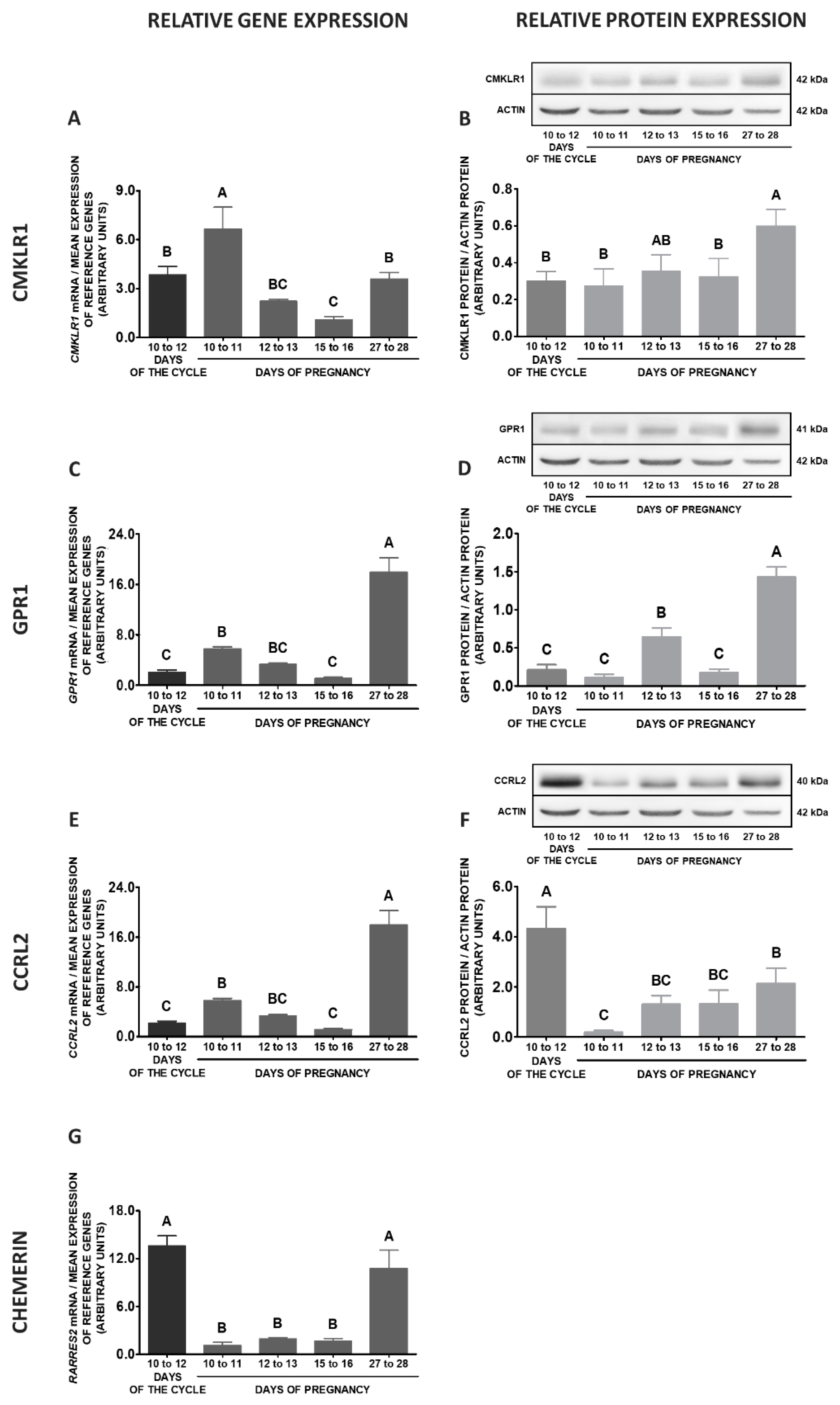
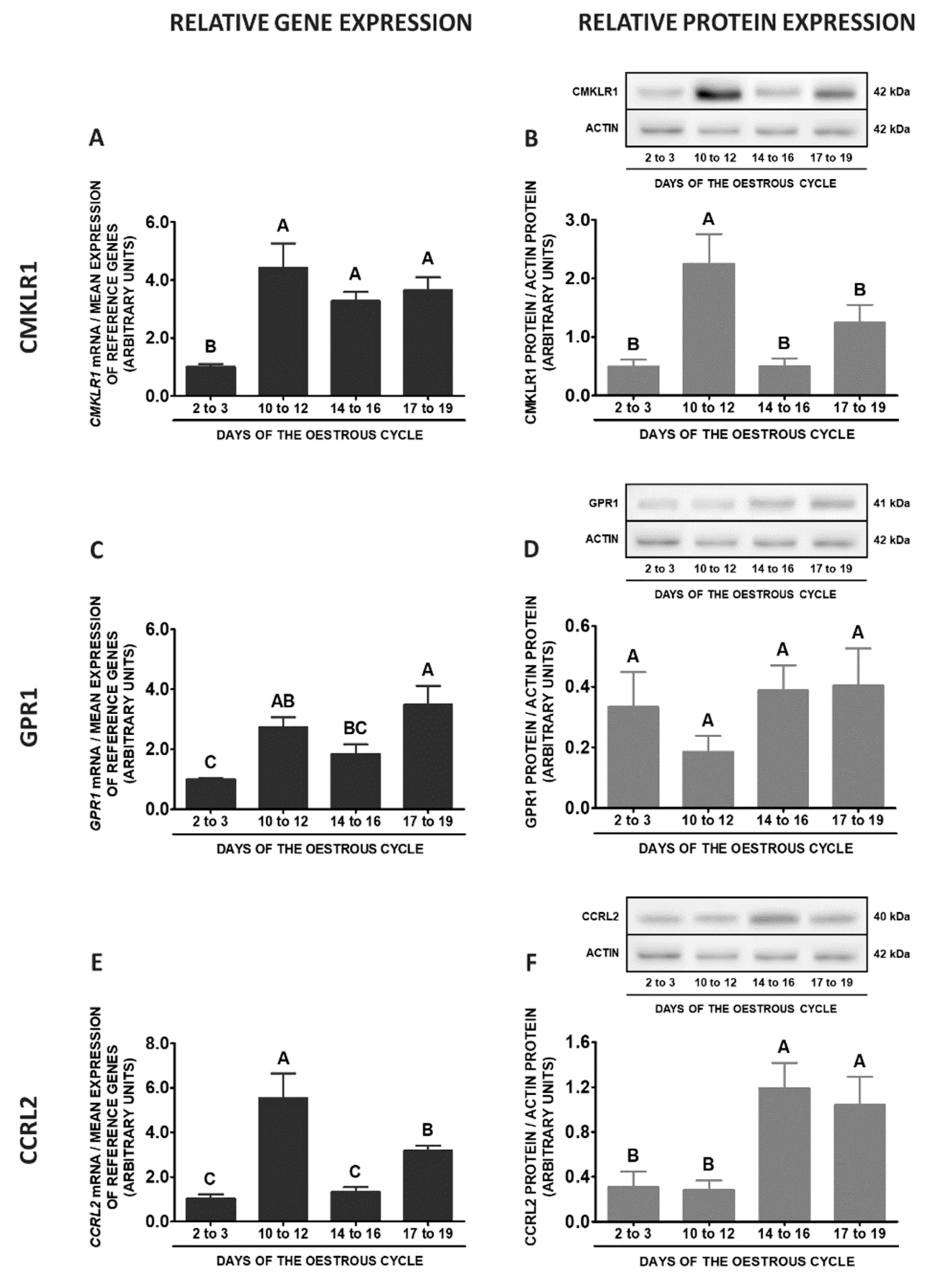
2.2. CHEM System Gene and Protein Expression in MBH during the Oestrous Cycle
2.3. CHEM System Gene and Protein Expression in MBH during Pregnancy
2.4. CHEM System Gene and Protein Expression in MBH—Pregnancy vs. Oestrous Cycle
2.5. CHEM System Gene and Protein Expression in POA during the Oestrous Cycle
2.6. CHEM System Gene and Protein Expression in POA during Pregnancy
2.7. CHEM System Gene and Protein Expression in POA—Pregnancy vs. Oestrous Cycle
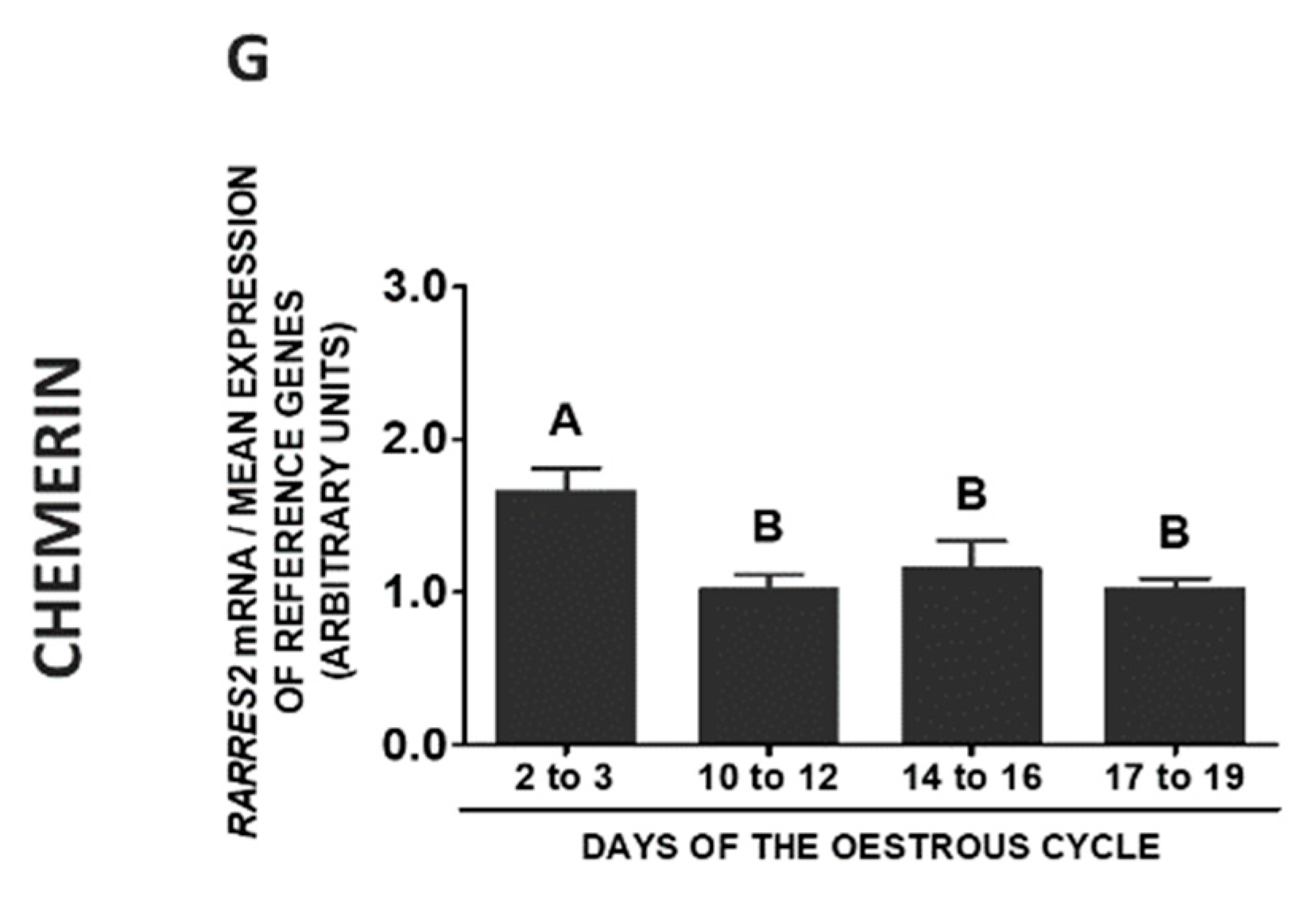
2.8. CHEM System Gene and Protein Expression in SME during the Oestrous Cycle
2.9. CHEM System Gene and Protein Expression in SME during Pregnancy
2.10. CHEM System Gene and Protein Expression in SME—Pregnancy vs. Oestrous Cycle
2.11. CHEM Concentrations in the Blood Plasma
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Tissue Collection
4.2. Fluorescent Immunohistochemistry of the Porcine Hypothalamus
4.3. Quantitative Real-Time PCR
4.4. Western Blotting
4.5. Enzyme-Linked Immune-Sorbent Assay (ELISA) of CHEM
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ernst, M.C.; Sinal, C.J. Chemerin: At the crossroads of inflammation and obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Goralski, K.B.; McCarthy, T.C.; Hanniman, E.A.; Zabel, B.A.; Butcher, E.C.; Parlee, S.D.; Muruganandan, S.; Sinal, C.J. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J. Biol. Chem. 2007, 282, 28175–28188. [Google Scholar] [CrossRef] [PubMed]
- Bozaoglu, K.; Bolton, K.; McMillan, J.; Zimmet, P.; Jowett, J.; Collier, G.; Walder, K.; Segal, D. Chemerin is a novel adipokine associated with obesity and metabolic syndrome. Endocrinology 2007, 148, 4687–4694. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Y.; Leungl, L.L. Proteolytic regulatory mechanism of chemerin bioactivity. Acta Biochim. Biophys. Sin. 2009, 41, 973–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rourke, J.L.; Dranse, H.J.; Sinal, C.J. Towards an integrative approach to understanding the role of chemerin in human health and disease. Obes. Rev. 2013, 14, 245–262. [Google Scholar] [CrossRef] [PubMed]
- Mattern, A.; Zellmann, T.; Beck-Sickinger, A.G. Processing, signaling, and physiological function of chemerin. IUBMB Life 2014, 66, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Chamberland, J.P.; Berman, R.L.; Aronis, K.N.; Mantzoros, C.S. Chemerin is expressed mainly in pancreas and liver, is regulated by energy deprivation, and lacks day/night variation in humans. Eur. J. Endocrinol. 2013, 169, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, M.; Cornuau, M.; Rame, C.; Guerif, F.; Royere, D.; Dupont, J. Chemerin inhibits IGF-1-induced progesterone and estradiol secretion in human granulosa cells. Hum. Reprod. 2012, 27, 1790–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Kim, J.Y.; Xue, K.; Liu, J.-Y.; Leader, A.; Tsang, B.K. Chemerin, a novel regulator of follicular steroidogenesis and its potential involvement in polycystic ovarian syndrome. Endocrinology 2012, 153, 5600–5611. [Google Scholar] [CrossRef]
- Miranda-Angulo, A.L.; Byerly, M.S.; Mesa, J.; Wang, H.; Blackshaw, S. Rax regulates hypothalamic tanycyte differentiation and barrier function in mice. J. Comp. Neurol. 2014, 522, 876–899. [Google Scholar] [CrossRef]
- Helfer, G.; Ross, A.W.; Thomson, L.M.; Mayer, C.D.; Stoney, P.N.; McCaffery, P.J.; Morgan, P.J. A neuroendocrine role for chemerin in hypothalamic remodelling and photoperiodic control of energy balance. Sci. Rep. 2016, 6, 26830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Fu, Y.; Xu, Y.; Weng, S.; Liu, D.; Cui, D.; Yu, S.; Liu, X.; Jiang, K.; Dong, Y. Chronic mild restraint stress rats decreased CMKLR1 expression in distinct brain region. Neurosci. Lett. 2012, 524, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Soda, Y.; Kanbe, K.; Liu, H.-Y.; Jinno, A.; Kitamura, T.; Hoshino, H. An orphan G protein-coupled receptor, GPR1, acts as a coreceptor to allow replication of human immunodeficiency virus types 1 and 2 in brain-derived cells. J. Virol. 1999, 73, 5231–5239. [Google Scholar]
- Banas, M.; Zegar, A.; Kwitniewski, M.; Zabieglo, K.; Marczynska, J.; Kapinska-Mrowiecka, M.; LaJevic, M.; Zabel, B.A.; Cichy, J. The expression and regulation of chemerin in the epidermis. PLoS One 2015, 10, e0117830. [Google Scholar] [CrossRef]
- Brunetti, L.; Orlando, G.; Ferrante, C.; Recinella, L.; Leone, S.; Chiavaroli, A.; Nisio, C.D.; Shohreh, R.; Manippa, F.; Ricciuti, A.; et al. Peripheral chemerin administration modulates hypothalamic control of feeding. Peptides 2014, 51, 115–121. [Google Scholar] [CrossRef]
- Ernst, M.C.; Haidl, I.D.; Zuniga, L.A.; Dranse, H.J.; Rourke, J.L.; Zabel, B.A.; Butcher, E.C.; Sinal, C.J. Disruption of the chemokine-like receptor 1 (CMKLR1) gene is associated with reduced adiposity and glucose intolerance. Endocrinology 2012, 153, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Xue, K.; Cao, M.; Wang, Q.; Liu, J.-Y.; Leder, A.; Han, J.Y.; Tsang, B.K. Chemerin suppresses ovarian follicular development and its potential involvement in follicular arrest in rats treated chronically with dihydrotestosterone. Endocrinology 2013, 154, 2912–2923. [Google Scholar] [CrossRef]
- Wang, Q.; Leader, A.; Tsang, B.K. Inhibitory roles of prohibitin and chemerin in FSH-induced rat granulosa cell steroidogenesis. Endocrinology 2013, 154, 956–967. [Google Scholar] [CrossRef]
- Tan, B.K.; Chen, J.; Farhatullah, S.; Adya, R.; Kaur, J.; Heutling, D.; Lewandowski, K.C.; O’Hare, J.P.; Lehnert, H.; Randeva, H.S. Insulin and metformin regulate circulating and adipose tissue chemerin. Diabetes 2009, 58, 1971–1977. [Google Scholar] [CrossRef]
- Duan, D.M.; Niu, J.M.; Lei, Q.; Lin, X.H.; Chen, X. Serum levels of the adipokine chemerin in preeclampsia. J. Perinat. Med. 2011, 40, 121–127. [Google Scholar] [CrossRef]
- Jin, C.H.; Yi, K.W.; Ha, Y.R.; Shin, J.H.; Park, H.T.; Kim, T.; Hur, J.Y. Chemerin expression in the peritoneal fluid, serum, and ovarian endometrioma of women with endometriosis. Am. J. Reprod. Immunol. 2015, 74, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Carlino, C.; Trotta, E.; Stabile, H.; Morrone, S.; Bulla, R.; Soriani, A.; Iannitto, M.L.; Agostinis, C.; Mocci, C.; Minozzi, M.; et al. Chemerin regulates NK cell accumulation and endothelial cell morphogenesis in the decidua during early pregnancy. J. Clin. Endocrinol. Metab. 2012, 97, 3603–3612. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Alvarez, R.; Garza-Rodriguez Mde, L.; Delgado-Enciso, I.; Trevino-Alvarado, V.M.; Canales-Del-Castillo, R.; Martinez-De-Villarreal, L.E.; Lugo-Trampe, A.; Tejero, M.E.; Schlabritz-Loutsevitch, N.E.; Rocha-Pizana Mdel, R.; et al. Molecular evolution and expression profile of the chemerine encoding gene RARRES2 in baboon and chimpanzee. Biol. Res. 2015, 48, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Yamaguchi, Y.; Sharif, S.; Du, X.Y.; Song, J.J.; Lee, D.M.; Recht, L.D.; Robinson, W.H.; Morser, J.; Leung, L.L. Chemerin158K protein is the dominant chemerin isoform in synovial and cerebrospinal fluids but not in plasma. J. Biol. Chem. 2011, 286, 39520–39527. [Google Scholar] [CrossRef] [PubMed]
- Bozaoglu, K.; Curran, J.E.; Stocker, C.J.; Zaibi, M.S.; Segal, D.; Konstantopoulus, N.; Morrison, N.; Carless, M.; Dyer, T.D.; Cole, S.A.; et al. Chemerin, a novel adipokine in the regulation of angiogenesis. J. Clin Endocrinol Metab. 2010, 95, 2476–2485. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, M.A.; Montes-Nieto, R.; Fernadez-Duran, E.; Insenser, M.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Evidence for masculinization of adipokine gene expression in visceral and subcutaneous adipose tissue of obese women with polycystic ovary syndrome (PCOS). J. Clin. Endocrinol. Metab. 2013, 98, E388–E396. [Google Scholar] [CrossRef] [PubMed]
- Garces, M.F.; Sanchez, E.; Ruiz-Parra, A.I.; Rubio-Romero, J.A.; Angel-Muller, A.; Suarez, M.A.; Bohorquez, L.F.; Bravo, S.B.; Nogueiras, R.; Dieguez, C.; et al. Serum chemerin levels during normal human pregnancy. Peptides 2013, 42, 138–143. [Google Scholar] [CrossRef]
- Garces, M.F.; Sanchez, E.; Acosta, B.J.; Angel, E.; Ruiz, A.I.; Rubio-Romero, J.A.; Dieguez, C.; Nogueiras, R.; Caminos, J.E. Expression and regulation of chemerin during rat pregnancy. Placenta 2012, 33, 373–378. [Google Scholar] [CrossRef]
- Dobrzyn, K.; Smolinska, N.; Szeszko, K.; Kiezun, M.; Maleszka, A.; Rytelewska, E.; Kaminski, T. Effect of progesterone on adiponectin system in the porcine uterus during early pregnancy. J. Anim. Sci. 2017, 95, 338–352. [Google Scholar] [CrossRef]
- Luque-Ramírez, M.; Martínez-García, M.Á.; Montes-Nieto, R.; Fernández-Durán, E.; Insenser, M.; Alpañés, M.; Escobar-Morreale, H.F. Sexual dimorphism in adipose tissue function as evidenced by circulating adipokine concentrations in the fasting state and after an oral glucose challenge. Hum. Reprod. 2013, 28, 1908–1918. [Google Scholar] [CrossRef]
- Bobjer, J.; Katrinaki, M.; Dermitzaki, E.; Margioris, A.N.; Giwercman, A.; Tsatsanis, C. Serum chemerin levels are negatively associated with male fertility and reproductive hormones. Hum. Reprod. 2018, 33, 2168–2174. [Google Scholar] [CrossRef] [PubMed]
- Brunetti, L.; Nisio, C.D.; Recinella, L.; Chiavaroli, A.; Leone, S.; Ferrante, C.; Orlando, G.; Vacca, M. Effects of vaspin, chemerin and omentin-1 on feeding behavior and hypotalamic peptidegene expression in the rat. Peptides 2011, 32, 1866–1871. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, M.; Bertoldo, M.J.; Rame, C.; Froment, P.; Dupont, J. Chemerin (RARRES2) decreases in vitro granulosa cell steroidogenesis and blocks oocyte meiotic progression in bovine species. Biol. Reprod. 2014, 90, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Akins, E.L.; Morrissette, M.C. Gross ovarian changes during estrous cycle of swine. Am. J. Vet. Res. 1968, 29, 1953–1957. [Google Scholar] [PubMed]
- Sesti, L.A.; Britt, J.H. Relationship of secretion of GnRH in vitro to changes in pituitary concentrations of LH and FSH and serum concentrations of LH during lactation in sows. J. Reprod. Fertil. 1993, 98, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolinska, N.; Dobrzyn, K.; Kiezun, M.; Szeszko, K.; Maleszka, A.; Kaminski, T. Effect of adiponectin on the steroidogenic acute regulatory protein, P450 side chain cleavage enzyme and 3β-hydroxysteroid dehydrogenase genes expression, progesterone and androstenedione production by the porcine uterus during early pregnancy. J. Physiol. Pharmacol. 2016, 67, 443–456. [Google Scholar] [PubMed]
- Martyniak, M.; Zglejc, K.; Franczak, A.; Kotwica, G. Expression of 3-hydroxysteroid dehydrogenase and P450 aromatase in porcine oviduct during the estrous cycle. J. Anim. Feed Sci. 2016, 25, 235–243. [Google Scholar] [CrossRef]
- Martyniak, M.; Franczak, A.; Kotwica, G. Interleukin-1β system in the oviducts of pigs during the oestrous cycle and early pregnancy. Theriogenology 2017, 96, 31–41. [Google Scholar] [CrossRef]
- Smolinska, N.; Kaminski, T.; Siawrys, G.; Przala, J. Long form of leptin receptor gene and protein expression in the porcine ovary during the estrous cycle and early pregnancy. Reprod. Biol. 2007, 7, 17–39. [Google Scholar]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers Sequences | Accession Number | Amplicon Length, bp | Primer, nM | Reference |
|---|---|---|---|---|---|
| RARRES2 | F: 5′-TGGAGGAGTTCCACAAGCAC-3′ | EU660865 | 154 | 500 | [The present study] |
| R: 5′-GCTTTCTTCCAGTCCCTCTTC-3′ | 500 | ||||
| CCRL2 | F: 5′-GAGCAGCAGCTACTTACTTCC-3′ | NM_001001617.1 | 196 | 200 | [The present study] |
| R: 5′-CTGCCCACTGACCGAGTTC-3′ | 200 | ||||
| CMKLR1 | F: 5′-GGACTACCACTGGGTGTTCG-3′ | EU660866 | 174 | 200 | [The present study] |
| R: 5′-GCCATGTAAGCCAGTCGGA-3′ | 200 | ||||
| GPR1 | F: 5′-ACCGACTTGGAGGAGAAAGC-3′ | FJ234899.1 | 159 | 200 | [The present study] |
| R: 5′-ATTGAGGAACCAGAGCGTGG-3′ | 200 | ||||
| UBC | F: 5′-GGAGGAATCTACTGGGGCGG-3′ | XM_003483411.3 | 103 | 400 | [37] |
| R: 5′-CAGAAGAAACGCAGGCAAACT-3′ | 400 | ||||
| 18sRNA | F: 5′-TCCAATGGATCCTCGCGGAA-3′ | AY265350.1 | 149 | 400 | [38] |
| R: 5′-GGCTACCACATCCAAGGAAG-3′ | 400 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smolinska, N.; Kiezun, M.; Dobrzyn, K.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Zaobidna, E.; Bogus-Nowakowska, K.; Wyrebek, J.; Bors, K.; et al. Expression of Chemerin and Its Receptors in the Porcine Hypothalamus and Plasma Chemerin Levels during the Oestrous Cycle and Early Pregnancy. Int. J. Mol. Sci. 2019, 20, 3887. https://doi.org/10.3390/ijms20163887
Smolinska N, Kiezun M, Dobrzyn K, Rytelewska E, Kisielewska K, Gudelska M, Zaobidna E, Bogus-Nowakowska K, Wyrebek J, Bors K, et al. Expression of Chemerin and Its Receptors in the Porcine Hypothalamus and Plasma Chemerin Levels during the Oestrous Cycle and Early Pregnancy. International Journal of Molecular Sciences. 2019; 20(16):3887. https://doi.org/10.3390/ijms20163887
Chicago/Turabian StyleSmolinska, Nina, Marta Kiezun, Kamil Dobrzyn, Edyta Rytelewska, Katarzyna Kisielewska, Marlena Gudelska, Ewa Zaobidna, Krystyna Bogus-Nowakowska, Joanna Wyrebek, Kinga Bors, and et al. 2019. "Expression of Chemerin and Its Receptors in the Porcine Hypothalamus and Plasma Chemerin Levels during the Oestrous Cycle and Early Pregnancy" International Journal of Molecular Sciences 20, no. 16: 3887. https://doi.org/10.3390/ijms20163887