MAS NMR on a Red/Far-Red Photochromic Cyanobacteriochrome All2699 from Nostoc

 , , , , and
, , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. NMR Spectroscopic Analyses
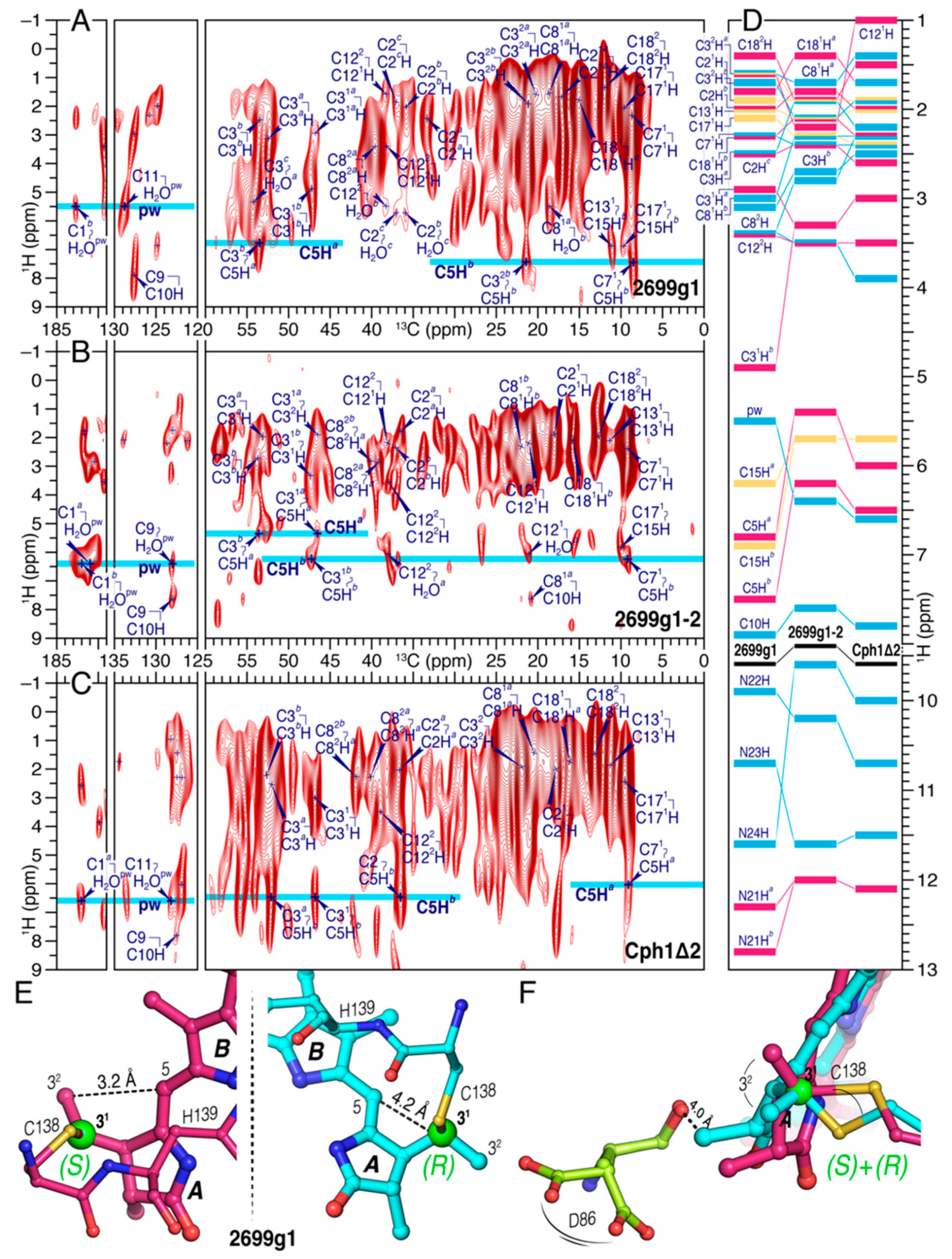
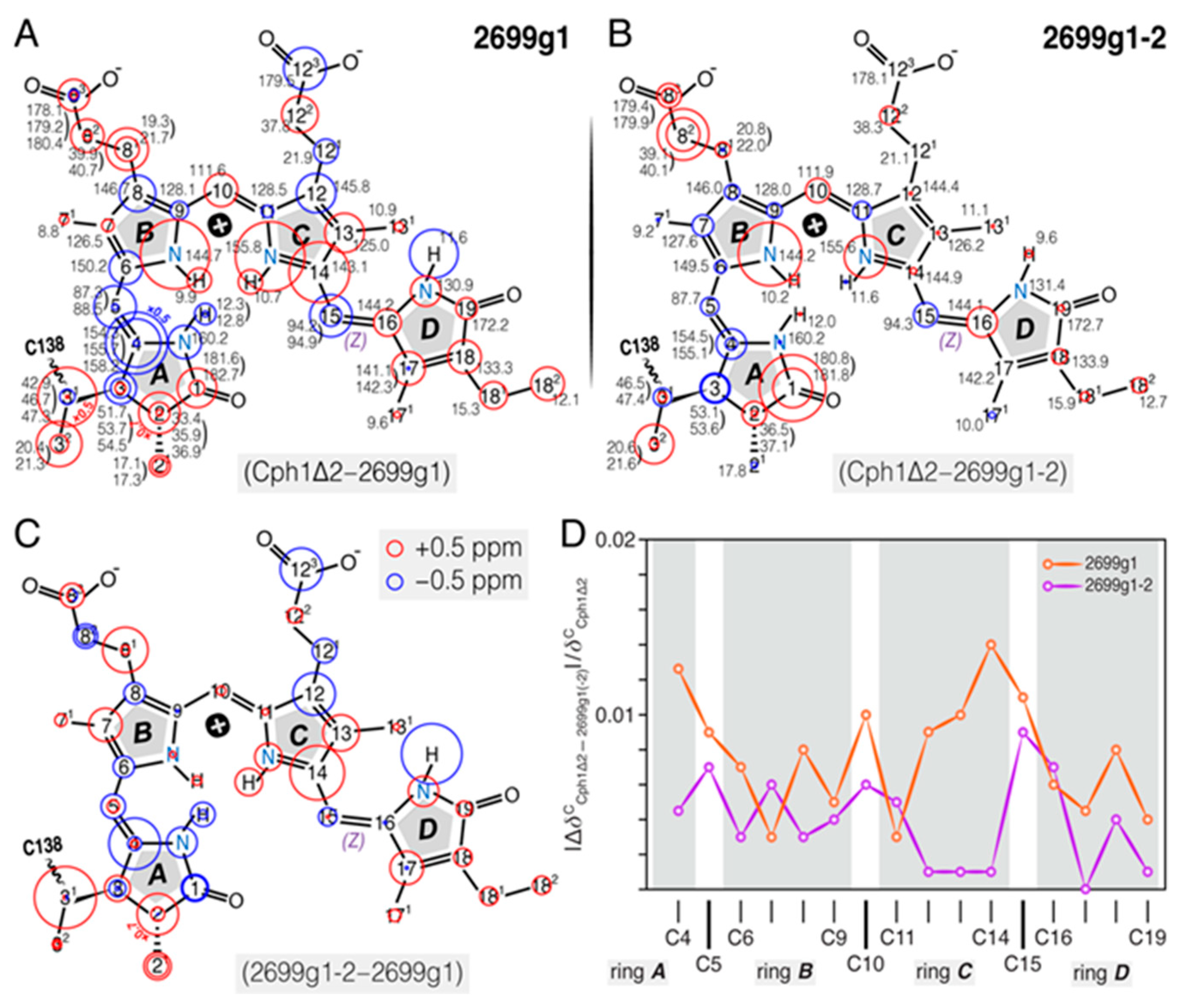
2.1.1. 13C Chemical Shift Assignments of the Chromophore in the Two all2699 Constructs
2.1.2. 1H–13C Correlation Spectra for Cph1Δ2 and the Two all2699 Protein Samples
2.1.3. 15N Assignments of Four Pyrrole Nitrogens in the Two all2699 Proteins
2.1.4. Comparison of 1H, 13C, and 15N Chemical Shifts of the Bilin in all2699g1 with all2699g1-2 and Cph1Δ2
2.2. Characterization of all2699g1-2 proteins
3. Discussion
3.1. Bilin Chromophore in the Pr dark States
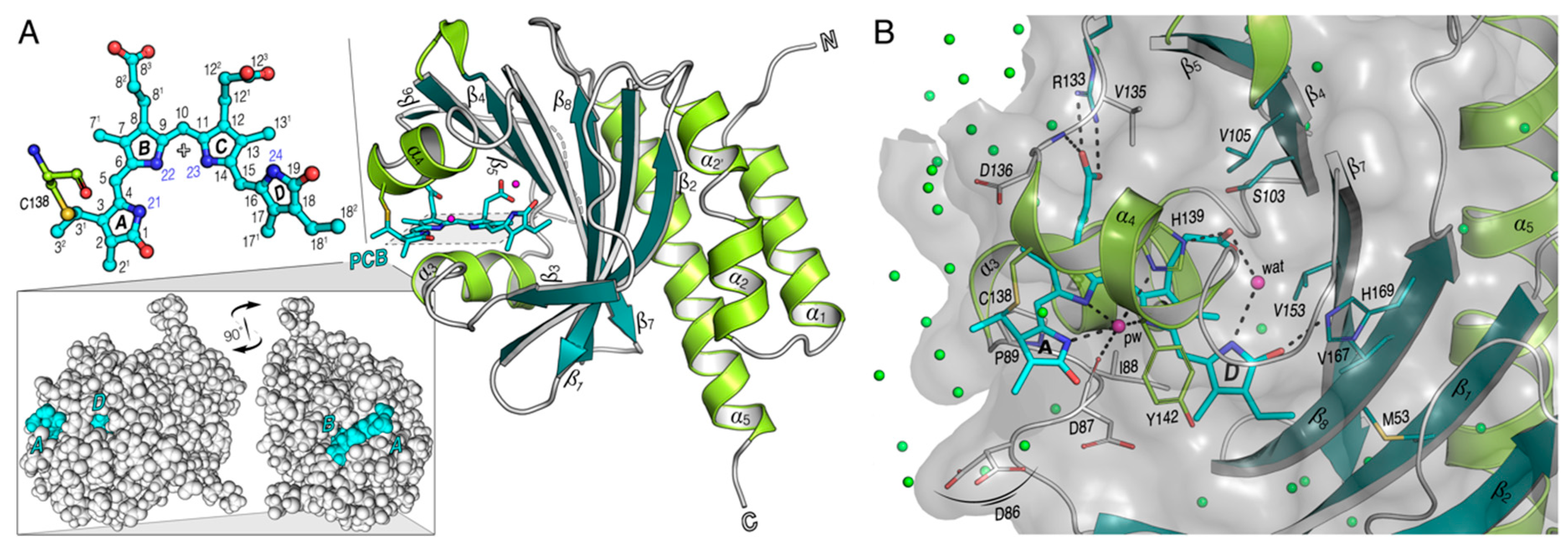
3.2. Heterogeneous Microenvironment of the GAF1-Bound PCB Chromophore in all2699 Proteins
3.3. Stereoselectivity of all2699 Apoproteins for PCB Chromophore
3.4. The Pyrrole Water
3.5. The Tongue
4. Materials and Methods
4.1. Sample Preparation
4.1.1. Cloning and Expression
4.1.2. Protein Production for NMR
4.1.3. UV-vis Spectroscopic Analyses
4.2. NMR Spectroscopic Analyses
4.3. Construction of Structural Model for all2699g1-2 Pr dark State
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rockwell, N.C.; Lagarias, J.C. A brief history of phytochromes. ChemPhysChem 2010, 11, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Anders, K.; Essen, L.-O. The family of phytochrome-like photoreceptors: Diverse, complex and multi-colored, but very useful. Curr. Opin. Struct. Biol. 2015, 35, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Burgie, E.S.; Walker, J.M.; Phillips, G.N., Jr.; Vierstra, R.D. A photo-labile thioether linkage to phycoviolobilin provides the foundation for the blue/green photocycles in DXCF-cyanobacteriochromes. Structure 2013, 21, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Mroginski, M.A.; Lang, C.; Kopycki, J.; Gärtner, W.; Matysik, J.; Hughes, J. 3D structures of plant phytochrome A as Pr and Pfr from solid-state NMR: Implications for molecular function. Front. Plant. Sci. 2018, 9, 498. [Google Scholar] [CrossRef] [PubMed]
- Ihalainen, J.A.; Gustavsson, E.; Schroeder, L.; Donnini, S.; Lehtivuori, H.; Isaksson, L.; Thöing, C.; Modi, V.; Berntsson, O.; Stucki-Buchli, B.; et al. Chromophore–protein interplay during the phytochrome photocycle revealed by step-scan FTIR spectroscopy. J. Am. Chem. Soc. 2018, 140, 12396–12404. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Yu, Q.; Gottlieb, S.M.; Chang, C.-W.; Rockwell, N.C.; Martin, S.S.; Madsen, D.; Lagarias, J.C.; Larsen, D.S.; Ames, J.B. Correlating structural and photochemical heterogeneity in cyanobacteriochrome NpR6012g4. Proc. Natl. Acad. Sci. USA 2018, 115, 4387–4392. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Sauthof, L.; Szczepek, M.; Fernandez Lopez, M.; Velazquez Escobar, F.; Qureshi, B.M.; Michael, N.; Buhrke, D.; Stevens, T.; Kwiatkowski, D.; et al. Structural snapshot of a bacterial phytochrome in its functional intermediate state. Nat. Commun. 2018, 9, 4912. [Google Scholar] [CrossRef]
- Yang, X.; Ren, Z.; Kuk, J.; Moffat, K. Temperature-scan cryocrystallography reveals reaction intermediates in bacteriophytochrome. Nature 2011, 479, 428–432. [Google Scholar] [CrossRef]
- Yang, X.; Kuk, J.; Moffat, K. Crystal structure of Pseudomonas aeruginosa bacteriophytochrome: Photoconversion and signal transduction. Proc. Natl. Acad. Sci. USA 2008, 105, 14715–14720. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ishizuka, T. Cyanobacteriochromes: A new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria. Photochem. Photobiol. Sci. 2008, 7, 1159–1167. [Google Scholar] [CrossRef]
- Fushimi, K.; Narikawa, R. Cyanobacteriochromes: Photoreceptors covering the entire UV-to-visible spectrum. Curr. Opin. Struct. Biol. 2019, 57C, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Hagen, K.D.; Chen, R.; Risser, D.D.; Ferreira, D.P.; Meeks, J.C. Genetic analysis reveals the identity of the photoreceptor for phototaxis in Hormogonium Filaments of Nostoc punctiforme. J. Bacteriol. 2015, 197, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Wiltbank, L.B.; Kehoe, D.M. Diverse light responses of cyanobacteria mediated by phytochrome superfamily photoreceptors. Nat. Rev. Microbiol. 2019, 17, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, J.; Luo, J.; Tu, J.-M.; Zeng, X.-L.; Xie, J.; Zhou, M.; Zhao, J.-Q.; Scheer, H.; Zhao, K.-H. Photophysical diversity of two novel cyanobacteriochromes with phycocyanobilin chromophores: Photochemistry and dark reversion kinetics. FEBS J. 2012, 279, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Martin, S.S.; Lagarias, J.C. Red/green cyanobacteriochromes: Sensors of color and power. Biochemistry 2012, 51, 9667–9677. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Martin, S.S.; Gulevich, A.G.; Lagarias, J.C. Phycoviolobilin formation and spectral tuning in the DXCF cyanobacteriochrome subfamily. Biochemistry 2012, 51, 1449–1463. [Google Scholar] [CrossRef] [PubMed]
- Narikawa, R.; Fukushima, Y.; Ishizuka, T.; Itoh, S.; Ikeuchi, M. A novel photoactive GAF domain of cyanobacteriochrome AnPixJ that shows reversible green/red photoconversion. J. Mol. Biol. 2008, 380, 844–855. [Google Scholar] [CrossRef]
- Xu, X.-L.; Gutt, A.; Mechelke, J.; Raffelberg, S.; Tang, K.; Miao, D.; Valle, L.; Borsarelli, C.D.; Zhao, K.-H.; Gärtner, W. Combined mutagenesis and kinetics characterization of the bilin-binding GAF domain of the protein Slr1393 from the cyanobacterium Synechocystis PCC6803. ChemBioChem 2014, 15, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Lamparter, T.; Mittmann, F.; Hartmann, E.; Gärtner, W.; Wilde, A.; Börner, T. A prokaryotic phytochrome. Nature 1997, 386, 663. [Google Scholar] [CrossRef]
- Yeh, K.-C.; Wu, S.-H.; Murphy, J.T.; Lagarias, J.C. A cyanobacterial phytochrome two-component light sensory system. Science 1997, 277, 1505–1508. [Google Scholar] [CrossRef]
- Hu, P.-P.; Guo, R.; Zhou, M.; Gärtner, W.; Zhao, K.-H. The red-/green-switching GAF3 of cyanobacteriochrome Slr1393 from Synechocystis sp. PCC6803 regulates the activity of an adenylyl cyclase. ChemBioChem 2018, 19, 1887–1895. [Google Scholar] [CrossRef] [PubMed]
- Blain-Hartung, M.; Rockwell, N.C.; Moreno, M.V.; Martin, S.S.; Gan, F.; Bryant, D.A.; Lagarias, J.C. Cyanobacteriochrome-based photoswitchable adenylyl cyclases (cPACs) for broad spectrum light regulation of cAMP levels in cells. J. Biol. Chem. 2018, 293, 8473–8483. [Google Scholar] [CrossRef] [PubMed]
- Essen, L.-O.; Mailliet, J.; Hughes, J. The structure of a complete phytochrome sensory module in the Pr ground state. Proc. Natl. Acad. Sci. USA 2008, 105, 14709–14714. [Google Scholar] [CrossRef] [PubMed]
- Burgie, E.S.; Bussell, A.N.; Walker, J.M.; Dubiel, K.; Vierstra, R.D. Crystal structure of the photosensing module from a red/far-red light-absorbing plant phytochrome. Proc. Natl. Acad. Sci. USA 2014, 111, 10179–10184. [Google Scholar] [CrossRef] [PubMed]
- Anders, K.; Daminelli-Widany, G.; Mroginski, M.A.; von Stetten, D.; Essen, L.-O. Structure of the cyanobacterial phytochrome 2 photosensor implies a tryptophan switch for phytochrome signaling. J. Biol. Chem. 2013, 288, 35714–35725. [Google Scholar] [CrossRef] [PubMed]
- Takala, H.; Björling, A.; Berntsson, O.; Lehtivuori, H.; Niebling, S.; Hoernke, M.; Kosheleva, I.; Henning, R.; Menzel, A.; Ihalainen, J.A.; et al. Signal amplification and transduction in phytochrome photosensors. Nature 2014, 509, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Stojković, E.A.; Toh, K.C.; Alexandre, M.T.A.; Baclayon, M.; Moffat, K.; Kennis, J.T.M. FTIR spectroscopy revealing light-dependent refolding of the conserved tongue region of bacteriophytochrome. J. Phys. Chem. Lett. 2014, 5, 2512–2515. [Google Scholar] [CrossRef]
- Song, C.; Rohmer, T.; Tiersch, M.; Zaanen, J.; Hughes, J.; Matysik, J. Solid-state NMR spectroscopy to probe photoactivation in canonical phytochromes. Photochem. Photobiol. 2013, 89, 259–273. [Google Scholar] [CrossRef]
- Rohmer, T.; Lang, C.; Hughes, J.; Essen, L.-O.; Gartner, W.; Matysik, J. Light-induced chromophore activity and signal transduction in phytochromes observed by 13C and 15N magic-angle spinning NMR. Proc. Natl. Acad. Sci. USA 2008, 105, 15229–15234. [Google Scholar] [CrossRef]
- Song, C.; Psakis, G.; Lang, C.; Mailliet, J.; Gärtner, W.; Hughes, J.; Matysik, J. Two ground state isoforms and a chromophore D-ring photoflip triggering extensive intramolecular changes in a canonical phytochrome. Proc. Natl. Acad. Sci. USA 2011, 108, 3842–3847. [Google Scholar] [CrossRef]
- Stöppler, D.; Song, C.; van Rossum, B.-J.; Geiger, M.-A.; Lang, C.; Mroginski, M.-A.; Jagtap, A.P.; Sigurdsson, S.T.; Matysik, J.; Hughes, J.; et al. Dynamic nuclear polarization provides new insights into chromophore structure in phytochrome photoreceptors. Angew. Chem. Int. Ed. Engl. 2016, 55, 16017–16020. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Velazquez Escobar, F.; Xu, X.-L.; Narikawa, R.; Ikeuchi, M.; Siebert, F.; Gärtner, W.; Matysik, J.; Hildebrandt, P. A red/green cyanobacteriochrome sustains its color despite a change in the bilin chromophore’s protonation state. Biochemistry 2015, 54, 5839–5848. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Strauss, H.M.; Schmieder, P. Heteronuclear NMR investigation on the structure and dynamics of the chromophore binding pocket of the cyanobacterial phytochrome Cph1. J. Am. Chem. Soc. 2008, 130, 11170–11178. [Google Scholar] [CrossRef] [PubMed]
- Huster, D.; Yao, L.; Hong, M. Membrane protein topology probed by 1H spin diffusion from lipids using solid-state NMR spectroscopy. J. Am. Chem. Soc. 2002, 124, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Narikawa, R.; Ishizuka, T.; Muraki, N.; Shiba, T.; Kurisu, G.; Ikeuchi, M. Structures of cyanobacteriochromes from phototaxis regulators AnPixJ and TePixJ reveal general and specific photoconversion mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 918–923. [Google Scholar] [CrossRef]
- Rockwell, N.C.; Martin, S.S.; Lim, S.; Lagarias, J.C.; Ames, J.B. Characterization of red/green cyanobacteriochrome NpR6012g4 by solution Nuclear Magnetic Resonance spectroscopy: A hydrophobic pocket for the C15-E,anti chromophore in the photoproduct. Biochemistry 2015, 54, 3772–3783. [Google Scholar] [CrossRef]
- Van Rossum, B.J.; Schulten, E.A.M.; Raap, J.; Oschkinat, H.; de Groot, H.J.M. A 3-D structural model of solid self-assembled chlorophyll a/H2O from multispin labelling and MAS NMR 2-D dipolar correlation spectroscopy in high magnetic field. J. Magn. Reson. 2002, 155, 1–14. [Google Scholar] [CrossRef]
- Lim, S.; Rockwell, N.C.; Martin, S.S.; Dallas, J.L.; Lagarias, J.C.; Ames, J.B. Photoconversion changes bilin chromophore conjugation and protein secondary structure in the violet/orange cyanobacteriochrome NpF2163g3. Photochem. Photobiol. Sci. 2014, 13, 951–962. [Google Scholar] [CrossRef]
- Anders, K.; Gutt, A.; Gärtner, W.; Essen, L.-O. Phototransformation of the red light sensor cyanobacterial phytochrome 2 from Synechocystis species depends on its tongue motifs. J. Biol. Chem. 2014, 289, 25590–25600. [Google Scholar] [CrossRef]
- Yang, X.; Stojković, E.A.; Kuk, J.; Moffat, K. Crystal structure of the chromophore binding domain of an unusual bacteriophytochrome, RpBphP3, reveals residues that modulate photoconversion. Proc. Natl. Acad. Sci. USA 2007, 104, 12571–12576. [Google Scholar] [CrossRef]
- Fischer, A.J.; Lagarias, J.C. Harnessing phytochrome’s glowing potential. Proc. Natl. Acad. Sci. USA 2004, 101, 17334–17339. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Psakis, G.; Kopycki, J.; Lang, C.; Matysik, J.; Hughes, J. The D-ring, not the A-ring, rotates in Synechococcus OS-B’ phytochrome. J. Biol. Chem. 2014, 289, 2552–2562. [Google Scholar] [CrossRef] [PubMed]
- Wiebeler, C.; Rao, A.G.; Gärtner, W.; Schapiro, I. The effective conjugation length is responsible for the red/green spectral tuning in the cyanobacteriochrome Slr1393g3. Angew. Chem. Int. Ed. Engl. 2019, 58, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Essen, L.-O.; Gärtner, W.; Hughes, J.; Matysik, J. Solid-state NMR spectroscopic study of chromophore–protein interactions in the Pr ground state of plant phytochrome A. Mol. Plant 2012, 5, 698–715. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.R.; Zhang, J.; Brunzelle, J.S.; Vierstra, R.D.; Forest, K.T. High resolution structure of Deinococcus bacteriophytochrome yields new insights into phytochrome architecture and evolution. J. Biol. Chem. 2007, 282, 12298–12309. [Google Scholar] [CrossRef] [PubMed]
- Mailliet, J.; Psakis, G.; Feilke, K.; Sineshchekov, V.; Essen, L.-O.; Hughes, J. Spectroscopy and a high-resolution crystal structure of Tyr263 mutants of cyanobacterial phytochrome Cph1. J. Mol. Biol. 2011, 413, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.R.; Hondred, D.; Walker, J.M.; Vierstra, R.D. Phytochrome requires the 6-kDa N-terminal domain for full biological activity. Proc. Natl. Acad. Sci. USA 1992, 89, 5039–5043. [Google Scholar] [CrossRef] [PubMed]
- Sineshchekov, V.A. Phytochrome A: Functional diversity and polymorphism. Photochem. Photobiol. Sci. 2004, 3, 596–607. [Google Scholar] [CrossRef]
- Sineshchekov, V.; Koppel, L.; Okamoto, H.; Wada, M. Fern Adiantum capillus-veneris phytochrome 1 comprises two native photochemical types similar to seed plant phytochrome A. J. Photochem. Photobiol. B Biol. 2014, 130, 20–29. [Google Scholar] [CrossRef]
- Velazquez Escobar, F.; Lang, C.; Takiden, A.; Schneider, C.; Balke, J.; Hughes, J.; Alexiev, U.; Hildebrandt, P.; Mroginski, M.A. Protonation-dependent structural heterogeneity in the chromophore binding site of cyanobacterial phytochrome Cph1. J. Phys. Chem. B 2017, 121, 47–57. [Google Scholar] [CrossRef]
- Yang, Y.; Linke, M.; von Haimberger, T.; Hahn, J.; Matute, R.; González, L.; Schmieder, P.; Heyne, K. Real-time tracking of phytochrome’s orientational changes during Pr photoisomerization. J. Am. Chem. Soc. 2012, 134, 1408–1411. [Google Scholar] [CrossRef] [PubMed]
- Nieder, J.B.; Brecht, M.; Bittl, R. Dynamic intracomplex heterogeneity of phytochrome. J. Am. Chem. Soc. 2009, 131, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Von Stetten, D.; Günther, M.; Scheerer, P.; Murgida, D.H.; Mroginski, M.A.; Krauß, N.; Lamparter, T.; Zhang, J.; Anstrom, D.M.; Vierstra, R.D.; et al. Chromophore heterogeneity and photoconversion in phytochrome crystals and solution studied by resonance Raman spectroscopy. Angew. Chem. Int. Ed. Engl. 2008, 47, 4753–4755. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Duanmu, D.; Martin, S.S.; Bachy, C.; Price, D.C.; Bhattacharya, D.; Worden, A.Z.; Lagarias, J.C. Eukaryotic algal phytochromes span the visible spectrum. Proc. Natl. Acad. Sci. USA 2014, 111, 3871–3876. [Google Scholar] [CrossRef] [PubMed]
- Nieder, J.B.; Stojković, E.A.; Moffat, K.; Forest, K.T.; Lamparter, T.; Bittl, R.; Kennis, J.T.M. Pigment–protein interactions in phytochromes probed by fluorescence line narrowing spectroscopy. J. Phys. Chem. B 2013, 117, 14940–14950. [Google Scholar] [CrossRef] [PubMed]
- Kirpich, J.S.; Mix, L.T.; Martin, S.S.; Rockwell, N.C.; Lagarias, J.C.; Larsen, D.S. Protonation heterogeneity modulates the ultrafast photocycle initiation dynamics of phytochrome Cph1. J. Phys. Chem. Lett. 2018, 9, 3454–3462. [Google Scholar] [CrossRef]
- Ihalainen, J.A.; Takala, H.; Lehtivuori, H. Fast photochemistry of prototypical phytochromes—A species vs. subunit specific comparison. Front. Mol. Biosci. 2015, 2, 75. [Google Scholar] [CrossRef]
- Kim, P.W.; Rockwell, N.C.; Freer, L.H.; Chang, C.-W.; Martin, S.S.; Lagarias, J.C.; Larsen, D.S. Unraveling the primary isomerization dynamics in cyanobacterial phytochrome Cph1 with multi-pulse manipulations. J. Phys. Chem. Lett. 2013, 4, 2605–2609. [Google Scholar] [CrossRef][Green Version]
- Kim, P.W.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Larsen, D.S. Dynamic inhomogeneity in the photodynamics of cyanobacterial phytochrome Cph1. Biochemistry 2014, 53, 2818–2826. [Google Scholar] [CrossRef]
- Gottlieb, S.M.; Kim, P.W.; Chang, C.-W.; Hanke, S.J.; Hayer, R.J.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Larsen, D.S. Conservation and diversity in the primary forward photodynamics of red/green cyanobacteriochromes. Biochemistry 2015, 54, 1028–1042. [Google Scholar] [CrossRef]
- Kim, P.W.; Freer, L.H.; Rockwell, N.C.; Martin, S.S.; Lagarias, J.C.; Larsen, D.S. Second-chance initiation dynamics of the cyanobacterial photocycle in the NpR6012 GAF4 domain of Nostoc punctiforme. J. Am. Chem. Soc. 2012, 134, 130–133. [Google Scholar] [CrossRef][Green Version]
- Scarbath-Evers, L.K.; Jähnigen, S.; Elgabarty, H.; Song, C.; Narikawa, R.; Matysik, J.; Sebastiani, D. Structural heterogeneity in a parent ground-state structure of AnPixJg2 revealed by theory and spectroscopy. Phys. Chem. Chem. Phys. 2017, 19, 13882–13894. [Google Scholar] [CrossRef] [PubMed]
- Rockwell, N.C.; Su, Y.-S.; Lagarias, J.C. Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Remberg, A.; Schmidt, P.; Braslavsky, S.E.; Gärtner, W.; Schaffner, K. Differential effects of mutations in the chromophore pocket of recombinant phytochrome on chromoprotein assembly and Pr-to-Pfr photoconversion. Eur. J. Biochem. 1999, 266, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Peersen, O.B.; Wu, X.; Smith, S.O. Enhancement of CP-MAS signals by variable-amplitude cross polarization. Compensation for inhomogeneous B1 fields. J. Magn. Reson. Ser. A 1994, 106, 127–131. [Google Scholar] [CrossRef]
- Thakur, R.S.; Kurur, N.D.; Madhu, P.K. Swept-frequency two-pulse phase modulation for heteronuclear dipolar decoupling in solid-state NMR. Chem. Phys. Lett. 2006, 426, 459–463. [Google Scholar] [CrossRef]
- Vinogradov, E.; Madhu, P.K.; Vega, S. Phase modulated Lee–Goldburg magic angle spinning proton nuclear magnetic resonance experiments in solid state: A bimodal Floquet theoretical treatment. J. Chem. Phys. 2001, 115, 8983–9000. [Google Scholar] [CrossRef]
- Paul, S.; Thakur, R.S.; Goswami, M.; Sauerwein, A.C.; Mamone, S.; Concistrè, M.; Förster, H.; Levitt, M.H.; Madhu, P.K. Supercycled homonuclear dipolar decoupling sequences in solid-state NMR. J. Magn. Reson. 2009, 197, 14–19. [Google Scholar] [CrossRef]
- Coelho, C.; Rocha, J.; Madhu, P.K.; Marfa, L. Practical aspects of Lee–Goldburg based CRAMPS techniques for high-resolution 1H NMR spectroscopy in solids: Implementation and applications. J. Magn. Reson. 2008, 194, 264–282. [Google Scholar] [CrossRef]
- Biasini, M.; Bienert, S.; Waterhouse, A.; Arnold, K.; Studer, G.; Schmidt, T.; Kiefer, F.; Gallo Cassarino, T.; Bertoni, M.; Bordoli, L.; et al. SWISS-MODEL: Modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 2014, 42, W252–W258. [Google Scholar] [CrossRef]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Q.-Z.; Bielytskyi, P.; Otis, J.; Lang, C.; Hughes, J.; Zhao, K.-H.; Losi, A.; Gärtner, W.; Song, C. MAS NMR on a Red/Far-Red Photochromic Cyanobacteriochrome All2699 from Nostoc. Int. J. Mol. Sci. 2019, 20, 3656. https://doi.org/10.3390/ijms20153656
Xu Q-Z, Bielytskyi P, Otis J, Lang C, Hughes J, Zhao K-H, Losi A, Gärtner W, Song C. MAS NMR on a Red/Far-Red Photochromic Cyanobacteriochrome All2699 from Nostoc. International Journal of Molecular Sciences. 2019; 20(15):3656. https://doi.org/10.3390/ijms20153656
Chicago/Turabian StyleXu, Qian-Zhao, Pavlo Bielytskyi, James Otis, Christina Lang, Jon Hughes, Kai-Hong Zhao, Aba Losi, Wolfgang Gärtner, and Chen Song. 2019. "MAS NMR on a Red/Far-Red Photochromic Cyanobacteriochrome All2699 from Nostoc" International Journal of Molecular Sciences 20, no. 15: 3656. https://doi.org/10.3390/ijms20153656
APA StyleXu, Q.-Z., Bielytskyi, P., Otis, J., Lang, C., Hughes, J., Zhao, K.-H., Losi, A., Gärtner, W., & Song, C. (2019). MAS NMR on a Red/Far-Red Photochromic Cyanobacteriochrome All2699 from Nostoc. International Journal of Molecular Sciences, 20(15), 3656. https://doi.org/10.3390/ijms20153656