Fibronectin Promotes Cell Growth and Migration in Human Renal Cell Carcinoma Cells


Abstract
: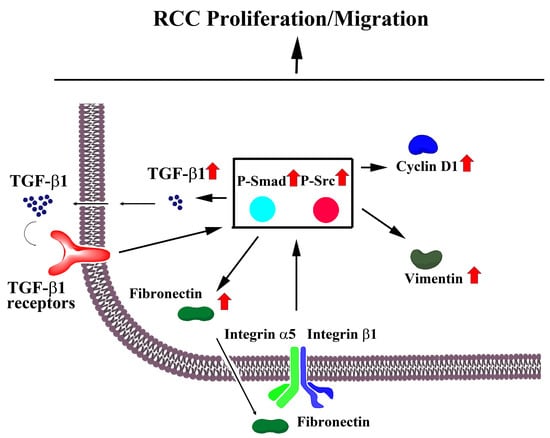
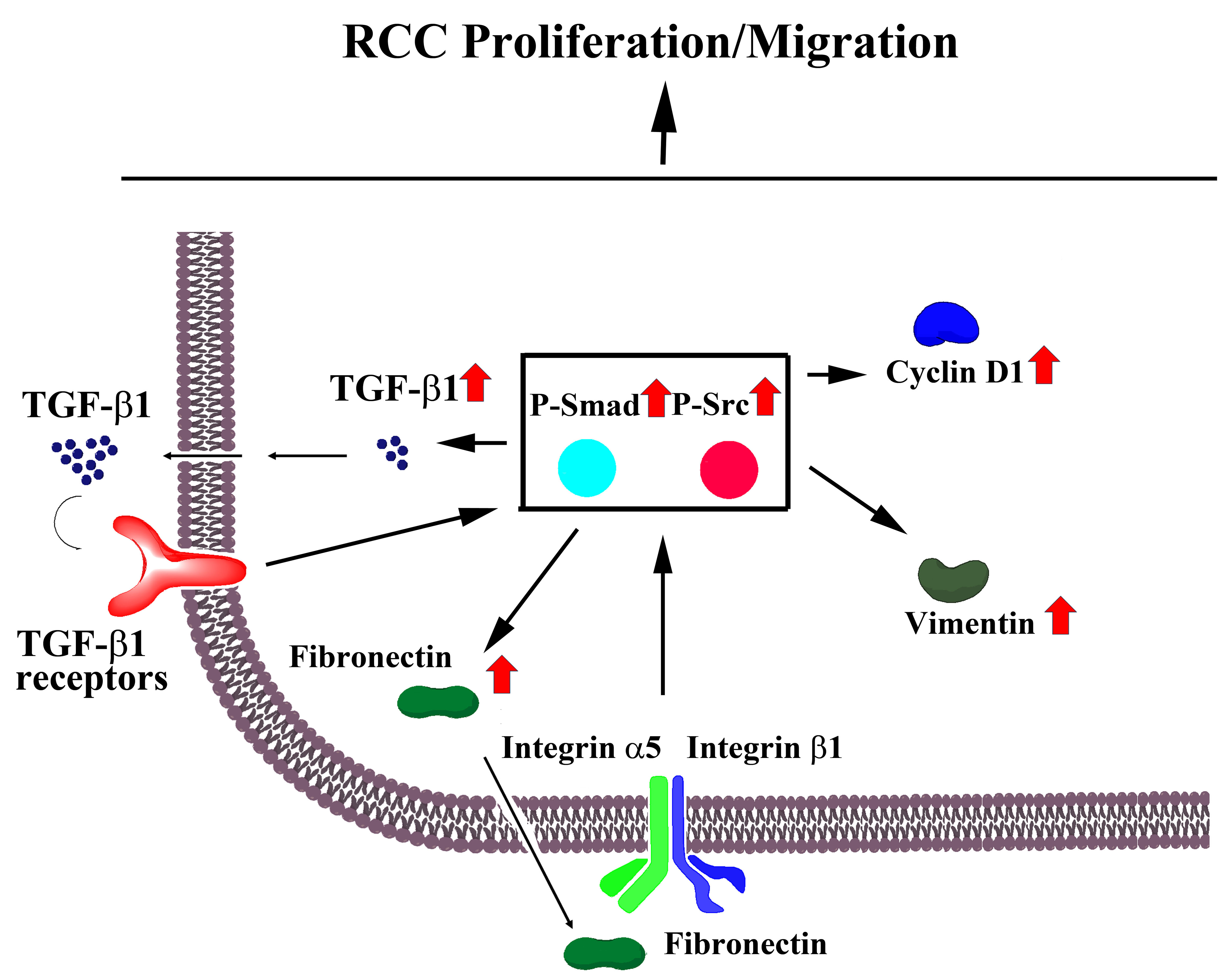{kind=link}
{kind=link}
{kind=link}
{kind=link}
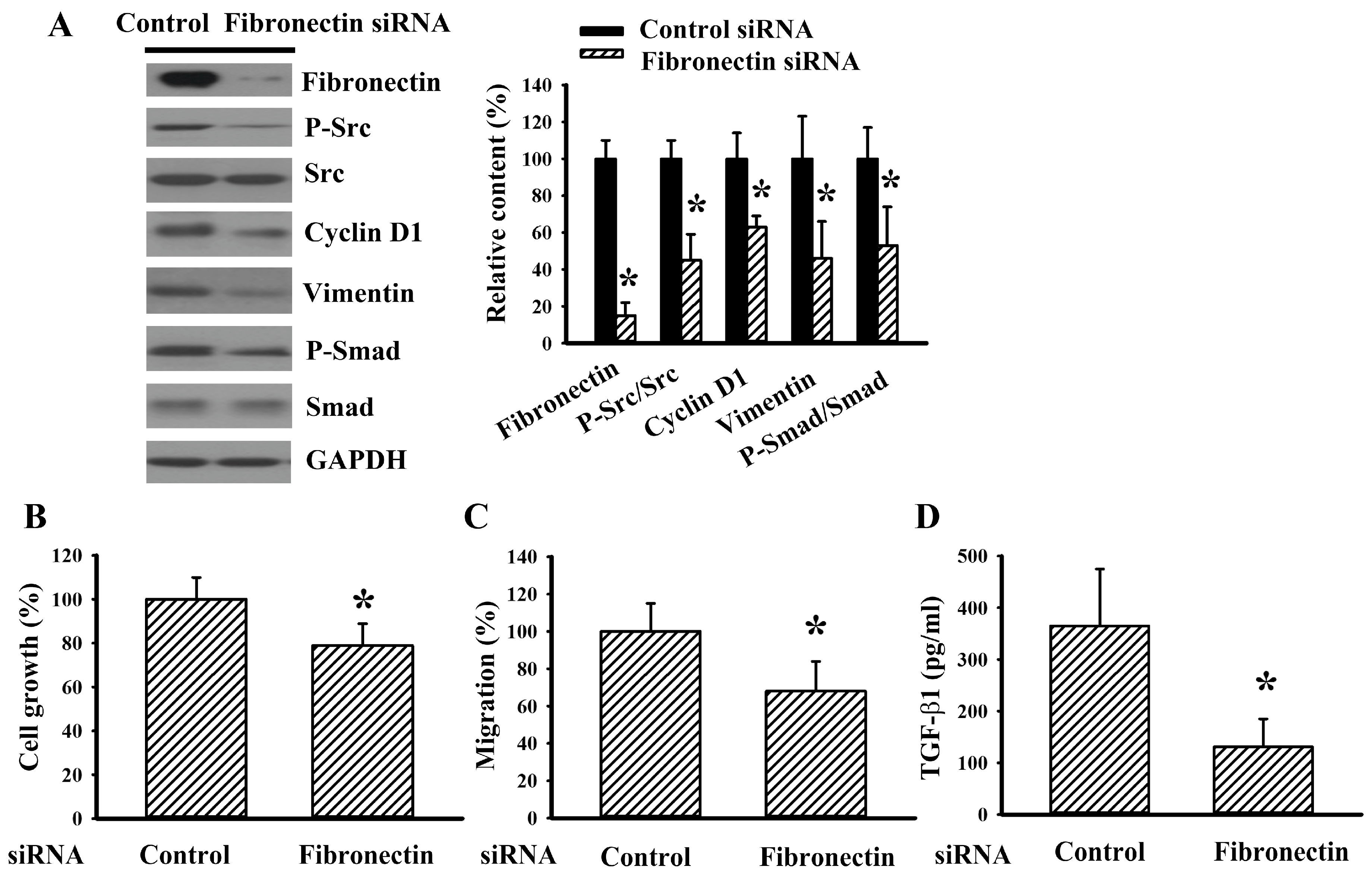{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Fibronectin Silencing Decreased Cell Growth and Migration
2.2. Integrin α5 and Integrin β1 Silencing Alleviated Fibronectin Effects
2.3. Fibronectin Silencing Decreased Intracellular Src Signaling
2.4. Fibronectin Silencing Decreased Cell Growth and Migration in Caki-1 Cells
2.5. Cell Growth and Migration Potential of RCC Cells Were Associated with Fibronectin
2.6. Fibronectin Increased Src and TGFβ-1 Signaling
2.7. Hypoxia Increased RCC Cell Migration
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Cell Growth
4.3. Colony Formation Assay
4.4. Wound Healing Assay
4.5. Cell Migration Assay
4.6. Small Interfering RNA (siRNA) Transfection
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Western Blot
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| RCC | Renal cell carcinoma |
| TGF-β1 | Transforming growth factor-β1 |
| EMT | Epithelial–mesenchymal transition |
| FAK | Focal adhesion kinase |
| ERK | Extracellular signal-regulated kinase |
| FBS | Fetal bovine serum |
| DMEM | Dulbecco’s modified Eagle medium |
| BSA | Bovine serum albumin |
| ELISA | Enzyme-linked immunosorbent assay |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| ECL | Enhanced chemiluminescence |
References
- Najjar, Y.G.; Rini, B.I. Novel agents in renal carcinoma: A reality check. Ther. Adv. Med. Oncol. 2012, 4, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Stadler, W.M. Targeted agents for the treatment of advanced renal cell carcinoma. Cancer 2005, 104, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Miller, J.D.; Li, J.Z.; Russell, M.W.; Charbonneau, C. Epidemiologic and socioeconomic burden of metastatic renal cell carcinoma (mRCC): A literature review. Cancer Treat. Rev. 2008, 34, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deep, G.; Kumar, R.; Jain, A.K.; Agarwal, C.; Agarwal, R. Silibinin inhibits fibronectin induced motility, invasiveness and survival in human prostate carcinoma PC3 cells via targeting integrin signaling. Mutat. Res. 2014, 768, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.R.; Lee, C.T.; Chang, K.Y.; Chang, W.C.; Liu, Y.W.; Lee, J.C.; Chen, B.K. Down-regulation of ARNT promotes cancer metastasis by activating the fibronectin/integrin β1/FAK axis. Oncotarget 2015, 6, 11530–11546. [Google Scholar]
- Ramos, G.; De, O.; Bernardi, L.; Lauxen, I.; Sant’Ana Filho, M.; Horwitz, A.R.; Lamers, M.L. Fibronectin modulates cell adhesion and signaling to promote single cell migration of highly invasive oral squamous cell carcinoma. PLoS ONE 2016, 11, e0151338. [Google Scholar] [CrossRef]
- Steffens, S.; Schrader, A.J.; Vetter, G.; Eggers, H.; Blasig, H.; Becker, J.; Kuczyk, M.A.; Serth, J. Fibronectin 1 protein expression in clear cell renal cell carcinoma. Oncol. Lett. 2012, 3, 787–790. [Google Scholar]
- Wang, W.Y.; Twu, C.W.; Liu, Y.C.; Lin, H.H.; Chen, C.J.; Lin, J.C. Fibronectin promotes nasopharyngeal cancer cell motility and proliferation. Biomed. Pharmacother. 2019, 109, 1772–1784. [Google Scholar] [CrossRef]
- Chaves, K.C.; Turaça, L.T.; Pesquero, J.B.; Mennecier, G.; Dagli, M.L.; Chammas, R.; Schor, N.; Bellini, M.H. Fibronectin expression is decreased in metastatic renal cell carcinoma following endostatin gene therapy. Biomed. Pharmacother. 2012, 66, 464–468. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, Y.; Sun, Y.; Chen, J.; Chang, C.; Wang, X.; Yeh, S. ERβ-mediated alteration of circATP2B1 and miR-204-3p signaling promotes invasion of clear cell renal cell carcinoma. Cancer Res. 2018, 78, 2550–2563. [Google Scholar] [CrossRef] [PubMed]
- Hegele, A.; Hofmann, R.; Kosche, B.; Kropf, J. Evaluation of cellular fibronectin plasma levels as a useful staging tool in different stages of transitional cell carcinoma of the bladder and renal cell carcinoma. Biomark. Insights 2007, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Knowles, L.M.; Gurski, L.A.; Maranchie, J.K.; Pilch, J. Fibronectin matrix formation is a prerequisite for colonization of kidney tumor cells in fibrin. J. Cancer 2015, 6, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Kondisetty, S.; Menon, K.N.; Pooleri, G.K. Fibronectin protein expression in renal cell carcinoma in correlation with clinical stage of tumour. Biomark. Res. 2018, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatino, M.; Kim-Schulze, S.; Panelli, M.C.; Stroncek, D.; Wang, E.; Taback, B.; Kim, D.W.; Deraffele, G.; Pos, Z.; Marincola, F.M.; et al. Serum vascular endothelial growth factor and fibronectin predict clinical response to high-dose interleukin-2 therapy. J. Clin. Oncol. 2009, 27, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, S.; Atschekzei, F.; Kramer, M.W.; Hennenlotter, J.; Vetter, G.; Becker, J.U.; Stenzl, A.; Merseburger, A.S.; Schrader, A.J.; Kuczyk, M.A.; et al. Fibronectin 1 mRNA expression correlates with advanced disease in renal cancer. BMC Cancer 2010, 10, 503. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, A.; Takakura, M.; Kanai, Y.; Sakuma, T.; Matsubara, J.; Honda, K.; Naito, S.; Yamada, T.; Ono, M. Use of quantitative shotgun proteomics to identify fibronectin 1 as a potential plasma biomarker for clear cell carcinoma of the kidney. Cancer Biomark. 2012, 10, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breuksch, I.; Prosinger, F.; Baehr, F.; Engelhardt, F.P.; Bauer, H.K.; Thüroff, J.W.; Heimes, A.S.; Hasenburg, A.; Prawitt, D.; Brenner, W. Integrin α5 triggers the metastatic potential in renal cell carcinoma. Oncotarget 2017, 8, 107530–107542. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Jia, D.; Wong, C.M.; Oi-Lin Ng, I.; Zhang, Z.; Liu, L.; Wang, Q.; Zhao, F.; Li, J.; Yao, M.; et al. SERPINA5 inhibits tumor cell migration by modulating the fibronectin-integrin β1 signaling pathway in hepatocellular carcinoma. Mol. Oncol. 2014, 8, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.G.; Gumbiner, B.M. Adhesion to fibronectin regulates Hippo signaling via the FAK-Src-PI3K pathway. J. Cell Biol. 2015, 210, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Knowles, L.M.; Gurski, L.A.; Engel, C.; Gnarra, J.R.; Maranchie, J.K.; Pilch, J. Integrin αvβ3 and fibronectin upregulate Slug in cancer cells to promote clot invasion and metastasis. Cancer Res. 2013, 73, 6175–6184. [Google Scholar] [CrossRef] [PubMed]
- Bagatur, Y.; Ilter Akulke, A.Z.; Bihorac, A.; Erdem, M.; Telci, D. Tissue transglutaminase expression is necessary for adhesion, metastatic potential and cancer stemness of renal cell carcinoma. Cell Adhes. Migr. 2018, 12, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.H.; Wang, F.; Wang, F.L.; Liu, Q.; Zhou, J. Regulation of transforming growth factor β-mediated epithelial-mesenchymal transition of lens epithelial cells by c-Src kinase under high glucose conditions. Exp. Ther. Med. 2018, 16, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Cseh, B.; Fernandez-Sauze, S.; Grall, D.; Schaub, S.; Doma, E.; Van Obberghen-Schilling, E. Autocrine fibronectin directs matrix assembly and crosstalk between cell-matrix and cell-cell adhesion in vascular endothelial cells. J. Cell Sci. 2010, 123, 3989–3999. [Google Scholar] [CrossRef] [PubMed]
- Wasil, L.R.; Shair, K.H. Epstein-Barr virus LMP1 induces focal adhesions and epithelial cell migration through effects on integrin-α5 and N-cadherin. Oncogenesis 2015, 4, e171. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.R.; Barcellos-de-Souza, P.; Sousa-Squiavinato, A.C.M.; Fernandes, P.V.; de Oliveira, I.M.; Boroni, M.; Morgado-Diaz, J.A. Annexin A2 overexpression associates with colorectal cancer invasiveness and TGF- ß induced epithelial mesenchymal transition via Src/ANXA2/STAT3. Sci. Rep. 2018, 8, 11285. [Google Scholar] [CrossRef]
- Lima, M.S.; Pereira, R.A.; Costa, R.S.; Tucci, S.; Dantas, M.; Muglia, V.F.; Ravinal, R.C.; Barros-Silva, G.E. The prognostic value of cyclin D1 in renal cell carcinoma. Int. Urol. Nephrol. 2014, 46, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, C.; Suh, J.H.; Chae, J.Y.; Moon, K.C. Nuclear expression of Smad proteins and its prognostic significance in clear cell renal cell carcinoma. Hum. Pathol. 2013, 44, 2047–2054. [Google Scholar] [CrossRef]
- Qayyum, T.; McArdle, P.A.; Lamb, G.W.; Jordan, F.; Orange, C.; Seywright, M.; Horgan, P.G.; Jones, R.J.; Oades, G.; Aitchison, M.A.; et al. Expression and prognostic significance of Src family members in renal clear cell carcinoma. Br. J. Cancer 2012, 107, 856–863. [Google Scholar] [CrossRef]
- Shi, Z.G.; Li, S.Q.; Li, Z.J.; Zhu, X.J.; Xu, P.; Liu, G. Expression of vimentin and survivin in clear cell renal cell carcinoma and correlation with p53. Clin. Transl. Oncol. 2015, 17, 65–73. [Google Scholar] [CrossRef]
- Yang, S.D.; Sun, R.C.; Mu, H.J.; Xu, Z.Q.; Zhou, Z.Y. The expression and clinical significance of TGF-beta1 and MMP2 in human renal clear cell carcinoma. Int. J. Surg. Pathol. 2010, 18, 85–93. [Google Scholar] [CrossRef]
- Wu, S.; Jiang, F.; Wu, H.; Wang, Y.; Xu, R.; Cao, J.; Lu, Q.; Zhu, X.; Zhong, Z.; Zhao, X. Prognostic significance of hypoxia inducible factor-1α expression in patients with clear cell renal cell carcinoma. Mol. Med. Rep. 2018, 17, 4846–4852. [Google Scholar] [CrossRef]
- Bluyssen, H.A.; Lolkema, M.P.; van Beest, M.; Boone, M.; Snijckers, C.M.; Los, M.; Gebbink, M.F.; Braam, B.; Holstege, F.C.; Giles, R.H.; et al. Fibronectin is a hypoxia-independent target of the tumor suppressor VHL. FEBS Lett. 2004, 556, 137–142. [Google Scholar] [CrossRef]
- Ou, Y.C.; Li, J.R.; Kuan, Y.H.; Raung, S.L.; Wang, C.C.; Hung, Y.Y.; Pan, P.H.; Lu, H.C.; Chen, C.J. Luteolin sensitizes human 786-O renal cell carcinoma cells to TRAIL-induced apoptosis. Life Sci. 2014, 100, 110–117. [Google Scholar] [CrossRef]
- Ou, Y.C.; Li, J.R.; Wang, J.D.; Chen, W.Y.; Kuanm, Y.H.; Yang, C.P.; Liao, S.L.; Lu, H.C.; Chen, C.J. Aspirin restores ABT-737-mediated apoptosis in human renal carcinoma cells. Biochem. Biophys. Res. Commun. 2018, 502, 187–193. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, Y.-C.; Li, J.-R.; Wang, J.-D.; Chang, C.-Y.; Wu, C.-C.; Chen, W.-Y.; Kuan, Y.-H.; Liao, S.-L.; Lu, H.-C.; Chen, C.-J. Fibronectin Promotes Cell Growth and Migration in Human Renal Cell Carcinoma Cells. Int. J. Mol. Sci. 2019, 20, 2792. https://doi.org/10.3390/ijms20112792
Ou Y-C, Li J-R, Wang J-D, Chang C-Y, Wu C-C, Chen W-Y, Kuan Y-H, Liao S-L, Lu H-C, Chen C-J. Fibronectin Promotes Cell Growth and Migration in Human Renal Cell Carcinoma Cells. International Journal of Molecular Sciences. 2019; 20(11):2792. https://doi.org/10.3390/ijms20112792
Chicago/Turabian StyleOu, Yen-Chuan, Jian-Ri Li, Jiaan-Der Wang, Cheng-Yi Chang, Chih-Cheng Wu, Wen-Ying Chen, Yu-Hsiang Kuan, Su-Lan Liao, Hsi-Chi Lu, and Chun-Jung Chen. 2019. "Fibronectin Promotes Cell Growth and Migration in Human Renal Cell Carcinoma Cells" International Journal of Molecular Sciences 20, no. 11: 2792. https://doi.org/10.3390/ijms20112792