A Novel Transcription Factor CaSBP12 Gene Negatively Regulates the Defense Response against Phytophthora capsici in Pepper (Capsicum annuum L.)


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. CaSBP12 Protein is Localized in the Nucleus
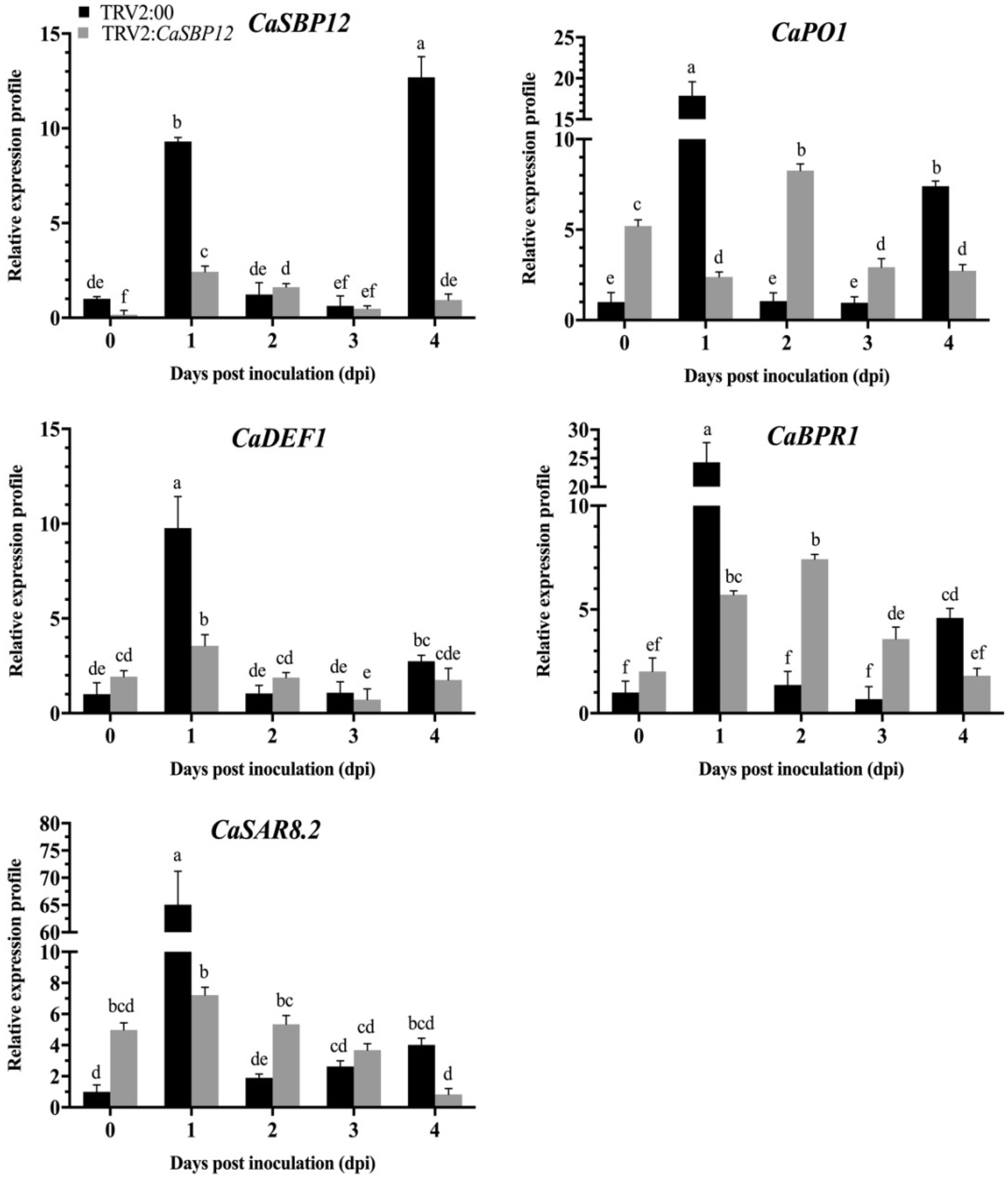
2.2. Silencing the CaSBP12 Gene Enhanced Pepper Plant Resistance to P. capsici Infection
2.2.1. Phenotypic Observation and Silencing Efficiency of CaSBP12 Gene Silencing Plants
2.2.2. Identification of CaSBP12 Gene Involvement in P. capsici Resistance
2.2.3. Determination of Root Activity of Gene-Silencing Plants
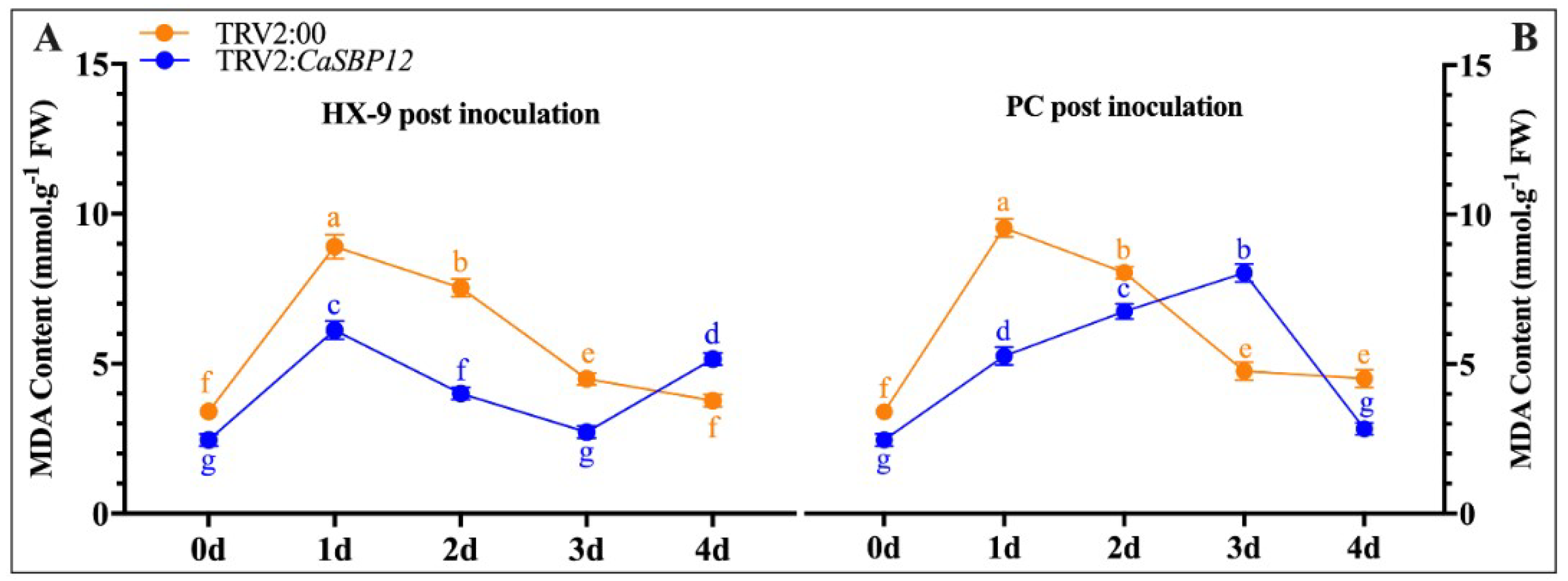
2.2.4. Determination of Malondialdehyde (MDA) Content
2.2.5. Disease Index Percent Statistics
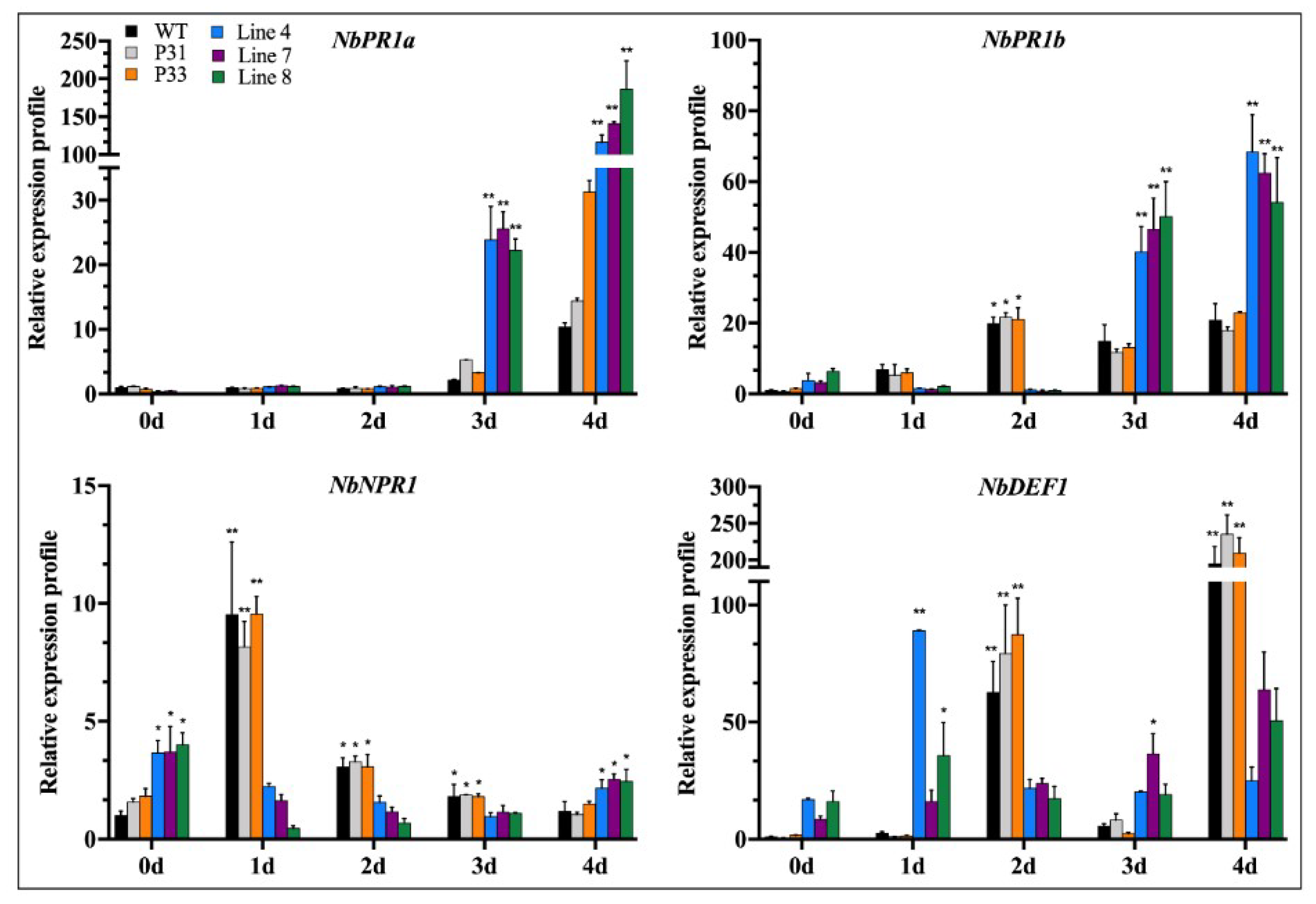
2.3. Overexpression of CaSBP12 in Nicotiana Benthamiana Enhanced Susceptibility to P. capsici Infection
2.3.1. Disease Resistance Identification of CaSBP12 Overexpression in Plants
2.3.2. Determination of Biochemical Indexes
2.3.3. Disease Index Percent Statistics
3. Discussion
4. Materials and Methods
4.1. Plant Material and Pathogen Preparation
4.2. Subcellular Localization of CaSBP12
4.3. Production of CaSBP12-silenced Pepper Plants by Virus-Induced Gene Silencing (VIGS)
4.4. Nicotiana Benthamiana Transformation
4.5. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.6. Determination of Root Activity
4.7. Determination of Malondialdehyde (MDA) Content
4.8. Determination of Ion Conductivity and Histochemical Staining
4.9. Determination of Antioxidant Activity
4.10. The Disease Index Percent Statistics
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SBP-box | Squamosa-promoter binding protein |
| MDA | Malondialdehyde |
| CaPO1 | Pepper peroxidase-like gene |
| CaBPR1 | Pepper pathogenesis-related (PR)-1 protein |
| CaDEF1 | Pepper defensin gene |
| EC | Electrical conductivity |
| NbDEF | Nicotiana benthamiana defensin gene |
| NbNPR1 | Nicotiana benthamiana non-expressor of PR genes |
| NbPR1a | Nicotiana benthamiana pathogenesis-related protein PR1a |
| NbPR1b | Nicotiana benthamiana pathogenesis-related protein PR1b |
References
- Jin, J.H.; Zhang, H.X.; Tan, J.Y.; Yan, M.J.; Li, D.W.; Khan, A.; Gong, Z.H. A new ethylene-responsive factor CaPTI1 gene of pepper (Capsicum annuum L.) involved in the regulation of defense response to Phytophthora capsici. Front Plant Sci. 2016, 6, 1217. [Google Scholar] [CrossRef] [PubMed]
- Granke, L.L.; Quesada-Ocampo, L.; Lamour, K.; Hausbeck, M.K. Advances in research on Phytophthora capsici on vegetable crops in the united states. Plant Dis. 2012, 96, 1588–1600. [Google Scholar] [CrossRef]
- Biles, C.L.; Brunton, B.D.; Wall, M.M.; Rivas, M. Phytophthora capsici zoospore infection of pepper fruit in various physical environments. Proc. Okla. Acad. Sci. 1995, 75, 1–5. [Google Scholar]
- Mou, S.L.; Liu, Z.Q.; Gao, F.; Yang, S.; Su, M.X.; Shen, L.; Wu, Y.; He, S.L. CaHDZ27, a Homeodomain-Leucine Zipper I (HD-Zip I) protein, positively regulates the resistance to Ralstonia solanacearum Infection in pepper. Mol. Plant-Microbe Interact. 2017, 30, 960–973. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Ma, Y.; Du, H.; Yang, Q.; Yu, D. Identification of transcriptional regulatory nodes in soybean defense networks using transient co-transactivation assays. Front. Plant Sci. 2015, 6, 915. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Mertens, J.; Goossens, A. Role and functioning of bHLH transcription factors in jasmonate signalling. J. Exp. Bot. 2017, 68, 1333–1347. [Google Scholar] [CrossRef]
- Xu, Z.S.; Chen, M.; Li, L.C.; Ma, Y.Z. Functions and application of the AP2/ERF transcription factor family in crop improvement. J. Integr. Plant Biol. 2011, 53, 570–585. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.; Saedler, H.; Huijser, P. A new family of DNA binding proteins includes putative transcriptional regulators of the Antirrhinum majus floral meristem identity gene SQUAMOSA. Mol. Gen. Genet. 1996, 250, 7–16. [Google Scholar] [PubMed]
- Lannenpaa, M.; Janonen, I.; Holtta-Vuori, M.; Gardemeister, M.; Porali, I.; Sopanen, T. A new SBP-box gene BpSPL1 in silver birch (Betulapendula). Physiol. Plant. 2004, 120, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.M.; Liang, X.; Nekl, E.R.; Stiers, J.J. Arabidopsis AtSPL14, a plant-specific SBP-domain transcription factor, participates in plant development and sensitivity to fumonisin B1. Plant J. 2005, 41, 744–754. [Google Scholar] [CrossRef]
- Hou, H.M.; Yan, Q.; Wang, X.P.; Xu, H. A SBP-Box gene VpSBP5 from Chinese wild vitis species responds to erysiphe necator and defense signaling molecules. Plant Mol. Biol. Rep. 2013, 31, 1261–1270. [Google Scholar] [CrossRef]
- Jung, C.; Yeu, S.Y.; Koo, Y.J.; Kim, M.; Choi, Y.D.; Cheong, J.J. Transcript Profile of Transgenic Arabidopsis Constitutively Producing Methyl Jasmonate. J. Plant Biol. 2007, 50, 12–17. [Google Scholar] [CrossRef]
- Song, A.; Gao, T.; Wu, D.; Xin, J.; Chen, S.; Guan, Z.; Wang, H.; Jin, L.; Chen, F. Transcriptome-wide identification and expression analysis of chrysanthemum SBP-like transcription factors. Plant Physiol. Biochem. 2016, 102, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Song, X.M.; Duan, W.K.; Wang, Y.; Hou, X.L. Genome-wide analysis of the SBP-box gene family in Chinese cabbage (Brassica rapasub sp. pekinensis). Genome Res. 2015, 58, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Li, R.J.; Sun, J.T.; Ma, F.; Zhang, H.X.; Jin, J.H.; Ali, M.; Haq, S.; Wang, J.; Gong, Z. Genome-wide analysis of dirigent gene family in pepper (Capsicum annuum L.) and characterization of CaDIR7 in biotic and abiotic stresses. Sci. Rep. 2018, 8, 5500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Jia, Q.L.; Li, D.W.; Wang, J.E.; Yin, Y.X.; Gong, Z.H. Characteristic of the pepper CaRGA2 gene in defense responses against Phytophthora capsici Leonian. Int. J. Mol. Sci. 2013, 14, 8985–9004. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, S.; Xiao, H.; Zhang, H.; Zhang, Z.; Jing, H. Overexpression of the CaTIP1-1 pepper gene in tobacco enhances resistance to osmotic stresses. Int. J. Mol. Sci. 2014, 15, 20101–20116. [Google Scholar] [CrossRef]
- Zhang, H.X.; Jin, J.H.; He, Y.M.; Lu, B.Y.; Li, D.W.; Chai, W.G.; Khan, A.; Gong, Z.H. Genome-wide identification and analysis of the SBP-Box family genes under Phytophthora capsici stress in pepper (Capsicum annuum L.). Front. Plant Sci. 2016, 7, 504. [Google Scholar] [CrossRef]
- Ma, N.N.; Zuo, Y.Q.; Liang, X.Q.; Yin, B.; Wang, G.D.; Meng, Q.W. The multiple stress-responsive transcription factor SlNAC1 improves the chilling tolerance of tomato. Physiol. Plant. 2013, 149, 474–486. [Google Scholar] [CrossRef]
- Wang, J.E. Expression Analysis and Functional Identification of CaRGA1 and CaPOD Genes Induced by Phytophthora capsici in Pepper; Institute of Horticulture, Northwest A&F University: Yangling, China, 2013. [Google Scholar]
- Cai, H.; Yang, S.; Yan, Y.; Xiao, Z.; Cheng, J.; Wu, J.; Qiu, A.; Lai, Y.; Mou, S.; Guan, D.; et al. CaWRKY6 transcriptionally activates CaWRKY40, regulates Ralstonia solanacearum resistance, and confers high-temperature and high-humidity tolerance in pepper. J. Exp. Bot. 2015, 66, 3163–3174. [Google Scholar] [CrossRef]
- Magbanua, Z.V.; De Moraes, C.M.; Brooks, T.D.; Williams, W.P.; Luthe, D.S. Is catalase activity one of the factors associated with maize resistance to Aspergillus flavus? Mol. Plant Microbe Interact. 2007, 20, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Esra, K.; AyşenSülün, Ü.; Cemil, İ.; YelizKaşko, A. Defence responses in leaves of resistant and susceptible pepper (Capsicum annuum L.) cultivars infected with different inoculum concentrations of Phytophthora capsici Leon. Sci. Hortic. 2011, 128, 434–442. [Google Scholar]
- Wu, Z.; Cao, Y.; Yang, R.; Qi, T.; Hang, Y.; Lin, H.; Zhou, G.; Wang, Ze.; Fu, C. Biotechnology for biofuels switchgrass SBP—Box transcription factors PvSPL1 and 2 function redundantly to initiate side tillers and affect biomass yield of energy crop. Biotechnol. Biofuels. 2016, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Kim, Y.J.; Hwang, B.K. The hypersensitive induced reaction and Leucine-Rich Repeat proteins regulate plant cell death associated with disease and plant immunity. Mol. Plant Microbe Interact. 2011, 24, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Hwang, B.K. Identification of the pepper SAR8.2 gene as a molecular marker for pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Planta 2003, 216, 387–396. [Google Scholar] [PubMed]
- Do, H.M.; Hong, J.K.; Jung, H.W.; Sang Hee Kim, S.H.; Ham, J.H.; Hwang, B.K. Expression of peroxidase-like genes, H2O2 production, and peroxidase activity during the hypersensitive response to Xanthomonas campestris pv. Vesicatoria in Capsicum annuum. Mol. Plant Microbe Interact. 2002, 16, 96–205. [Google Scholar]
- Kim, Y.J.; Hwang, B.K. Pepper gene encoding a basic pathogenesis-related 1 protein is pathogen and ethylene inducible. Physiol. Plant. 2000, 108, 51–60. [Google Scholar]
- Do, H.M.; Lee, S.C.; Jung, H.W.; Sohn, K.H.; Hwang, B.K. Differential expression and in situ localization of a pepper defensin (CADEF1) gene in response to pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Plant Sci. 2004, 166, 1297–1305. [Google Scholar]
- Zhang, Y.L. Defence Responses of Pepper (Capsicum annuum L.) and Grafting Seedings to Phytophthora capsici Leonia and the Function Analysis of CaRGA2 Gene; Institute of Horticulture, Northwest A&F University: Yangling, China, 2013. [Google Scholar]
- Wu, Y.X.; Zeng, F.H. A preliminary study on the relationship between induced resistance to bacterial blight and defense enzymes in hybrid rice seedlings. Acta Phytophysiol. Sin. 1996, 26, 127–131. [Google Scholar]
- Wang, L.P. Studies on the Relationships of Some Physilogical-Chemical Changes and Disease Resisantce in Cucumber Infected with Fusarium Wilt Disease; College of Horticulture, Shanxi Agricultural University: Taigu, China, 2001. [Google Scholar]
- Koç, E.; Üstün, A.S. Influence of Phytophthora capsici L. inoculation on disease severity, necrosis length, peroxidase and catalase activity, and phenolic content of resistant and susceptible pepper (Capsicum annuum L.) plants. Turk. J. Biol. 2012, 36, 357–371. [Google Scholar]
- Liu, J.Q.; Chen, X.J.; Liang, X.X.; Zhou, X.G.; Yang, F.; Liu, J.; He, S.Y.; Guo, Z.J. Alternative splicing of rice WRKY62 and WRKY76 transcription factor genes in pathogen defense. Plant Physiol. 2016, 01921. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zheng, X.; Huang, X.; Lii, Y.; Shao, C.; Xu, Y.; Chen, F. A novel transcription factor JcNAC1 response to stress in new model woody plant Jatrophacurcas. Planta 2014, 239, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Delessert, C.; Kazan, K.; Wilson, I.W.; Van Der Straeten, D.; Manners, J.; Dennis, E.S.; Dolferus, R. The transcription factor ATAF2 represses the expression of pathogenesis-related genes in Arabidopsis. Plant J. 2005, 43, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, M.M.S.; Burch-Smith, S.; Czymmek, T.M.; Huijser, K.; Dinesh-Kumar, P.; Savithramma, P. Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor-mediated plant innate immunity. PLoS Pathog. 2013, 9, e1003235. [Google Scholar] [CrossRef] [PubMed]
- Cheol Song, G.; Sim, H.J.; Kim, S.G.; Ryu, C.M. Root-mediated signal transmission of systemic acquired resistance against above-ground and below-ground pathogens. Ann. Bot. 2016, 118, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Verhage, A.; Van Wees, S.C.M.; Pieterse, C.M.J. Plant immunity: It’s the hormones talking, but what do they say? Plant Physiol. 2010, 154, 536–540. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Brooks, D.M. Cross talk between signaling pathways in pathogen defense. Curr. Opin. Plant Biol. 2002, 5, 325–331. [Google Scholar] [CrossRef]
- Wu, L.J.; Zhang, H.M.; Zhang, J.; Wu, L.C.; Xi, Z.Y.; Chen, Y.H. Overexpression of Zm-HINT1 in Arabidopsis thaliana enhances resistance to Fusarium graminearum. PCTOC 2015, 121, 511–518. [Google Scholar] [CrossRef]
- Zhang, Z. The Development of ssr Markers in Pepper and the Functional Analysis of CaMYB in Anthocyanin Biosynthetic Pathway of Pepper; Institute of Horticulture, Northwest A&F University: Yangling, China, 2015. [Google Scholar]
- Oh, S.K.; Park, J.M.; Joung, Y.H.; Lee, S.; Chung, E.; Kim, S.Y.; Yu, S.H.; Choi, D. A plant EPF-type zinc-finger protein, CaPIF1, involved in defence against pathogens. Mol. Plant Pathol. 2005, 6, 269–285. [Google Scholar] [CrossRef]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper (Capsicum annuum L.) leaves subjected to chilling stress. Genet. Mol. Res. 2012, 11, 4063–4080. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.X.; Wang, S.B.; Zhang, H.X.; Xiao, H.J.; Jin, J.H.; Ji, J.J.; Jing, H.; Chen, R.G.; Arisha, M.H.; Gong, Z.H. Cloning and expression analysis of CaPIP1-1 gene in pepper (Capsicum annuum L.). Gene 2015, 563, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Du, Y. Phytophthora infestans RXLR Effector AVR1 and Its Host Target Sec5. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2014. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Rampuria, S.; Singh, N.K.; Shukla, P.; Kirti, P.B. Characterization of a vacuolar processing enzyme expressed in Arachis diogoi in resistance responses against late leaf spot pathogen, Phaeoisariopsis personata. Plant Mol. Biol. 2015, 88, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Hwang, B.K. Pepper heat shock protein 70a interacts with the type III effector AvrBsT and triggers plant cell death and immunity. Plant Physiol. 2015, 167, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.W.; Hwang, B.K. The pepper extracellular peroxidase CaPO2 is required for salt, drought and oxidative stress tolerance as well as resistance to fungal pathogens. Planta 2012, 235, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, R.; Nuckles, E.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Zhang, Y.L. Identification of Physiological Race of Phytophthora capsici and Analysis of the Efficiency of Chemical Control; Institute of Horticulture, Northwest A&F University: Yangling, China, 2009. [Google Scholar]












© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-X.; Ali, M.; Feng, X.-H.; Jin, J.-H.; Huang, L.-J.; Khan, A.; Lv, J.-G.; Gao, S.-Y.; Luo, D.-X.; Gong, Z.-H. A Novel Transcription Factor CaSBP12 Gene Negatively Regulates the Defense Response against Phytophthora capsici in Pepper (Capsicum annuum L.). Int. J. Mol. Sci. 2019, 20, 48. https://doi.org/10.3390/ijms20010048
Zhang H-X, Ali M, Feng X-H, Jin J-H, Huang L-J, Khan A, Lv J-G, Gao S-Y, Luo D-X, Gong Z-H. A Novel Transcription Factor CaSBP12 Gene Negatively Regulates the Defense Response against Phytophthora capsici in Pepper (Capsicum annuum L.). International Journal of Molecular Sciences. 2019; 20(1):48. https://doi.org/10.3390/ijms20010048
Chicago/Turabian StyleZhang, Huai-Xia, Muhammad Ali, Xiao-Hui Feng, Jing-Hao Jin, Liu-Jun Huang, Abid Khan, Jing-Gang Lv, Su-Yan Gao, De-Xu Luo, and Zhen-Hui Gong. 2019. "A Novel Transcription Factor CaSBP12 Gene Negatively Regulates the Defense Response against Phytophthora capsici in Pepper (Capsicum annuum L.)" International Journal of Molecular Sciences 20, no. 1: 48. https://doi.org/10.3390/ijms20010048
APA StyleZhang, H.-X., Ali, M., Feng, X.-H., Jin, J.-H., Huang, L.-J., Khan, A., Lv, J.-G., Gao, S.-Y., Luo, D.-X., & Gong, Z.-H. (2019). A Novel Transcription Factor CaSBP12 Gene Negatively Regulates the Defense Response against Phytophthora capsici in Pepper (Capsicum annuum L.). International Journal of Molecular Sciences, 20(1), 48. https://doi.org/10.3390/ijms20010048