Comparative Proteome Analysis Reveals Lipid Metabolism-Related Protein Networks in Response to Rump Fat Mobilization

Abstract
:1. Introduction
2. Results
2.1. Persistent Starvation Experiment
2.2. Changes in Rump Width and Serum Lipid Markers
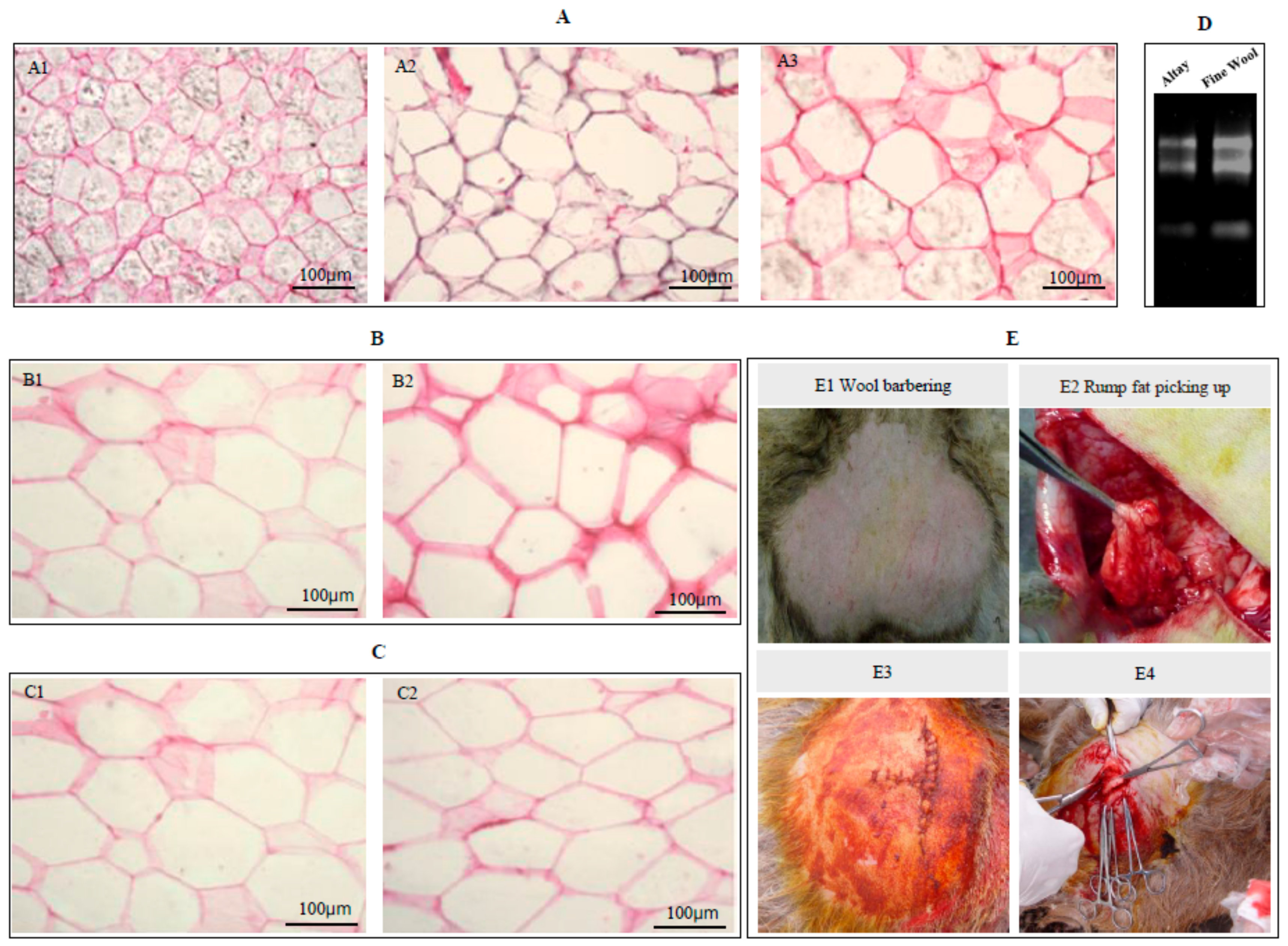
2.3. Cell Volumes of Sheep Adipocytes
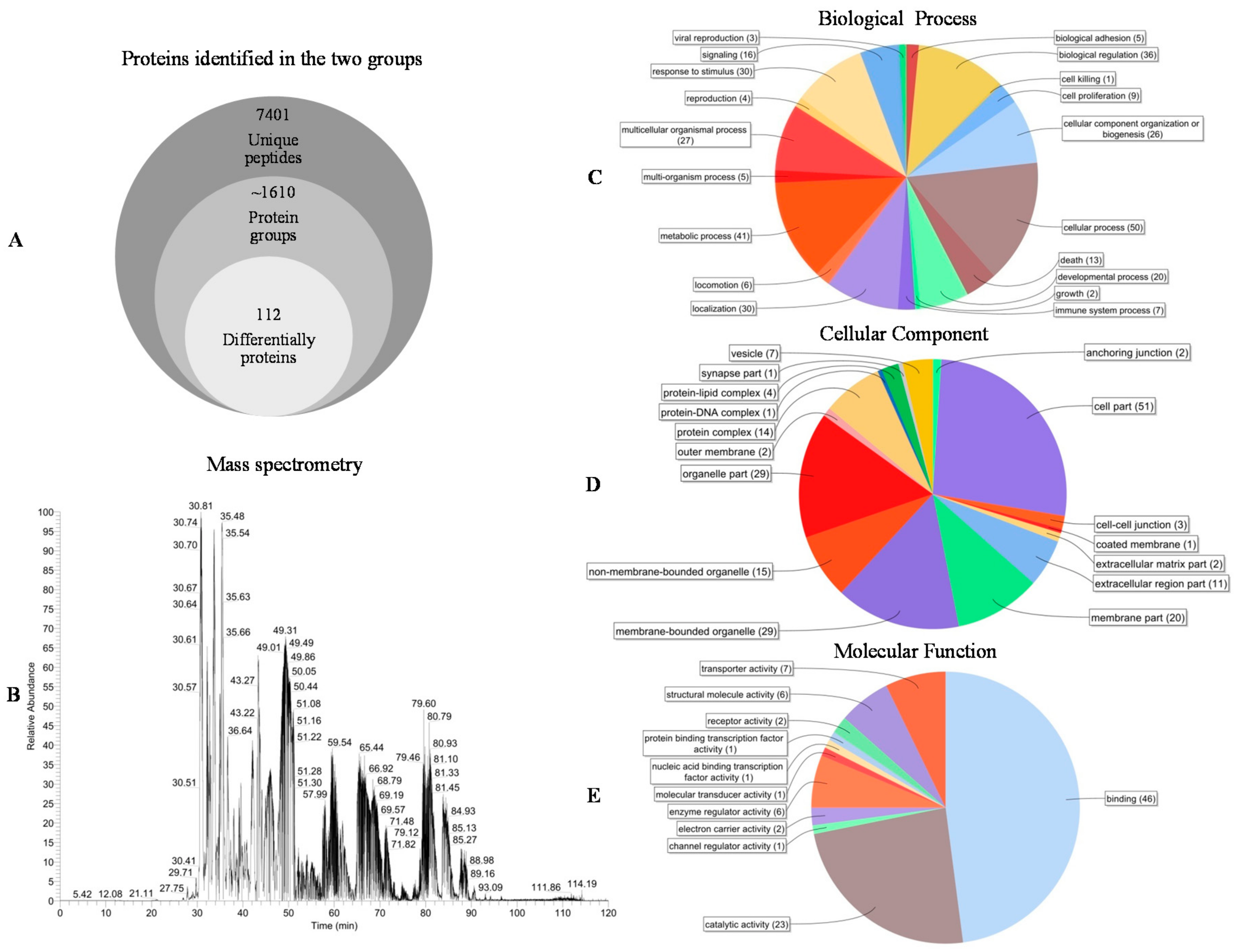
2.4. iTRAQ Analysis of Proteins Participating in Altay Sheep Rump Fat Metabolism under Persistent Starvation
2.5. qPCR Analysis of Differentially Expressed Candidate Genes
3. Discussion
3.1. Altay Sheep Present an Ideal Model for Studying Animal Fat Metabolism
3.2. Roles of Specific Hormones in Lipid Metabolism
3.3. Synergistic Actions of Key Lipid Metabolism-Related Hormones and Signaling Pathways
3.4. Inhibition of Lipid Droplet Fusion Ability in Combination with Enhanced Lipolysis
3.5. Activation of Critical Genes for Maintaining Internal Homeostasis
3.5.1. Insulin Resistance Promotes Lipolysis
3.5.2. Initiation of Body Stress Mechanisms
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Design
4.3. Altay Sheep Rump Width Distribution
4.4. Determination of the Serum Lipid Markers in Altay Sheep
4.5. Frozen Sections and Hematoxylin-Eosin Staining
4.6. iTRAQ Sample Preparation
4.7. LC and MS/MS Analyses
4.8. Database Searches and Analysis of the Identified Proteins
4.9. qPCR Analysis
4.10. Data Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| iTRAQ | Isobaric peptide labeling approach |
| WAT | White adipose tissue |
| BAT | Brown adipose tissue |
| MSC | Mesenchymal stem cells |
| LC-MS | Liquid chromatography-mass spectrometry |
| AMPK | AMP-activated protein kinase |
| MAPK | Mitogen-activated protein kinase |
| ERK | Extracellular signal-regulated protein kinase |
| JNK | Extracellular signal-regulated protein kinase |
| p38 MAPK | p38 Mitogen-activated protein kinase |
| PPAR | Peroxisome proliferator-activated receptor |
| HPLC | High-performance liquid chromatography |
| GO | Gene ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| FDR | False discovery rate |
| ELISA | Enzyme-linked immunosorbent assay |
| HE | Hematoxylin-eosin |
| SDS | Sodium dodecyl sulfate |
| DTT | Dithiothreitol |
| FA | Formic acid |
| ACN | Acetonitrile |
| ANOVA | Analysis of variance |
| TG | Triglyceride |
| FFA | Free Fatty Acid |
| TCH | Total Cholesterol |
| LEP | Leptin |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| Apo-AI | Apolipoprotein AI |
| ITGA6 | Integrin alpha 6 |
| CFD | Complement factor D |
| p120GAP | p120 GTPase activating protein |
| HSP72 | Heat-shock protein 72 |
| LPL | Lipoprotein lipase |
| PP2A | Protein Phosphatase 2A |
| ADIPOQ | C1Q and collagen domain-containing, Adiponectin, APN |
| CIEDC | Cell death-inducing DFFA-like effector C |
| FLNA | Filamin A |
| RETN | Resistin |
| C/EBP | CCAAT/enhancer binding protein |
| FAS | Fatty acid synthase |
| HSL | Hormone-sensitive lipase |
| ATGL | Adipose triglyceride lipase |
| TNF-α | Tumor necrosis factor-α |
References
- Rocha, J.; Chen, S.; Beja-Pereira, A. Molecular evidence for fat-tailed sheep domestication. Trop. Anim. Health Prod. 2011, 43, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Li, M.L.; Yuan, B.J.M.; Li, M.L.; Yuan, B.H. Winter feed for transhumant livestock in China: The Altay experience. World Anim. Rev. 1996, 87, 38–44. [Google Scholar]
- Liu, C.; Zhu, Z.P.; Liu, Y.F.; Guo, T.J. Diversity and abundance of the rumen and fecal methanogens in Altay sheep native to Xinjiang and the influence of diversity on methane emissions. Arch. Microbiol. 2012, 194, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.C.; Lane, M.D. Adipose development: From stem cell to adipocyte. Crit. Rev. Biochem. Mol. 2005, 40, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Kahn, C.R. Triglycerides and toggling the tummy. Nat. Genet. 2002, 25, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M.; Flier, J.S. Obesity and the regulation of energy balance. Cell 2014, 104, 531–543. [Google Scholar] [CrossRef]
- Parlee, S.D.; Lentz, S.I.; Mori, H.; MacDougald, O.A. Quantifying size and number of adipocytes in adipose tissue. Methods Enzymol. 2014, 537, 93–122. [Google Scholar] [PubMed]
- Cwiklinska, A.; Gliwinska, A.; Senderowska, Z.; Kortas-Stempak, B.; Kuchta, A.; Dabkowski, K.; Jankowski, M. Impact of phosphatidylcholine liposomes on the compositional changes of VLDL during lipoprotein lipase(LPL)-mediated lipolysis. Chem. Phys. Lipids 2016, 195, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Schroeder-Gloeckler, J.M.; Rahman, S.M.; Janssen, R.C.; Qiao, L.; Shao, J.; Roper, M.; Fischer, S.J.; Lowe, E.; Orlicky, M.J.; McManaman, J.L.; et al. CCAAT/enhancer-binding protein beta deletion reduces adiposity, hepatic steatosis, and diabetes in Lepr(db/db) mice. J. Biol. Chem. 2007, 282, 15717–15729. [Google Scholar] [CrossRef] [PubMed]
- Nakano, R.; Kurosaki, E.; Yoshida, S.; Yokono, M.; Shimaya, A.; Maruyama, T.; Shibasaki, M. Antagonism of peroxisome proliferator-activated receptor γ prevents high-fat diet-induced obesity in vivo. Biochem. Pharmacol. 2006, 72, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, H.; Iritani, N.; Sugimoto, T.; Ikeda, H. Transcriptional regulation of fatty acid synthase gene by insulin/glucose, polyunsaturated fatty acid and leptin in hepatocytes and adipocytes in normal and genetically obese rats. FEBS J. 1999, 260, 505–511. [Google Scholar] [CrossRef]
- Saltiel, A.R. Another hormone-sensitive triglyceride lipase in fat cells? Proc. Natl. Acad. Sci. USA 2000, 97, 535–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Suh, Y.; Choi, Y.M.; Shin, S.; Han, J.Y.; Lee, K. Loss of fat with increased adipose triglyceride lipase-mediated lipolysis in adipose tissue during laying stages in Quail. Lipids 2013, 48, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.X.; Liu, K.Y.; Li, P.; Han, S.; Peng, X.D.; Shen, L. Adiponectin is expressed in the pancreas of high-fat-diet-fed mice and protects pancreatic endothelial function during the development of type 2 diabetes. Diabetes Metab. 2014, 40, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Ortega, L.; Riestra, P.; Navarro, P.; Gavala-Perez, T.; Soriano-Guillen, L.; Garces, C. Resistin levels are related to fat mass, but not to body mass index in children. Peptides 2013, 49, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Gomez-Smith, M.; Qin, Z.; Duquette, P.M.; Cardenas-Blanco, A.; Rai, P.S.; Haeper, M.; Tsai, E.C.; Anisman, H.; Chen, H. Ablation of LMO4 in glutamatergic neurons impairs leptin control of fat metabolism. Cell Mol. Life Sci. 2012, 69, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Miazgowski, T.; Safranow, K.; Krzyzanowska-Awiniarska, B.; Iskierska, K.; Widecka, K. Adiponectin, visfatin and regional fat depots in normal weight obese premenopausal women. Eur. J. Clin. Investig. 2013, 43, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Olszanecka-Glinianowicz, M.; Chudek, J.; Kocelak, P.; Szromek, A.; Zahorska-Markiewicz, B. Body fat changes and activity of tumor necrosis factor α system-a 5-year follow-up study. Metabolism 2011, 60, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, G.; Zhang, R.; Guo, J.; Li, C.; Martin, G.; Chen, Y.; Wang, X. Comparative proteomic analyses using iTRAQ-labeling provides insights into fiber diversity in sheep and goats. J. Proteom. 2018, 172, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Etherton, T.D.; Aberle, E.D.; Thompson, E.H.; Allen, C.E. Effects of cell size and animal age on glucose metabolism in pig adipose tissue. J. Lipid Res. 1981, 22, 72–80. [Google Scholar] [PubMed]
- Gondret, F.; Lebret, B. Feeding intensity and dietary protein level affect adipocyte cellularity and lipogenic capacity of muscle homogenates in growing pigs, without modification of the expression of sterol regulatory element binding protein. J. Anim. Sci. 2002, 80, 3184–3193. [Google Scholar] [CrossRef] [PubMed]
- Van den-Heuvel, J.K.; Eggels, L.; Van-Rozen, A.J.; Luijendijk, M.C.; Fliers, E.; Kalsbeek, A.; Adan, R.A.; La-Fleur, S.E. Neuropeptide Y and leptin sensitivity is dependent on diet composition. J. Neuroendocrinol. 2014, 26, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Takahashi, N.; Hileman, S.M.; Patel, H.R.; Pajvani, U.B.; Scherer, P.E.; Ahima, R.S. Adiponectin acts in the brain to decrease body weight. Nat. Med. 2004, 10, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Dib, J.; Thomas, A.; Delahaut, P.; Fichant, E.; Schanzer, W.; Thevis, M. Identification and characterization of in vitro and in vivo generated metabolites of the adiponectin receptor agonists AdipoRon and 112,254. J. Pharmaceut. Biomed. 2016, 125, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Lapsys, N.M.; Kriketos, A.D.; Lim-Fraser, M.; Poynten, A.M.; Lowy, A.; Furler, S.M.; Chisholm, D.J.; Cooney, G.J. Expression of genes involved in lipid metabolism correlate with peroxisome proliferator activated receptor gamma expression in human skeletal lmuscle. J. Clin. Endocr. Metab. 2000, 85, 4293–4297. [Google Scholar] [PubMed]
- Ren, M.Q.; Wegner, J.; Bellmann, O.; Brockmann, G.A.; Schneider, F.; Teuscher, F.; Ender, K. Comparing mRNA levels of genes encoding leptin, leptin receptor, and lipoprotein lipase between dairy and beef cattle. Domest. Anim. Endocrin. 2002, 23, 371–381. [Google Scholar] [CrossRef]
- Lee, Y.H.; Choi, S.H.; Lee, K.W.; Kim, D.J. Apolipoprotein B/A1 ratio is associated with free androgen index and visceral adiposity and may be an indicator of metabolic syndrome in male children and adolescents. Clin. Endocrinol. 2011, 74, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minokoshi, Y.; Kim, Y.; Peroni, O.D.; Fryer, L.G.D.; Muller, C.; Carling, D.; Kahn, B.B. Leptin stimulates fatty-acid oxidation by activating AMP-activated proteinkinase. Nature 2002, 415, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Nagasawa, M.; Hara, T.; Ide, T.; Murakami, K. Differential roles of Cidea and Cidec in insulin-induced anti-apoptosis and lipid droplet (LD) formation in human adipocytes. J. Lipid Res. 2010, 51, 1676–1684. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Nagasawa, M.; Hara, T.; Ide, T.; Murakami, K. Differential regulation of CIDEA and CIDEC expression by insulin via Akt1/2 and JNK2-dependent pathways in human adipocytes. J. Lipid Res. 2011, 52, 1450–1460. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.Y.; Sun, Z.Q.; Li, P. CIDE proteins and metabolic disorders. Curr. Opin. Lipidol. 2009, 20, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, S.J.; Hong, S.H.; Bajracharya, R.; Yang, S.Y.; Lee, K.S.; Yu, K. Drosophila Adiponectin receptor in insulin producing cells regulates glucose and lipid metabolism by controlling insulin secretion. PLoS ONE 2013, 8, e68641. [Google Scholar] [CrossRef] [PubMed]
- Sliwicka, E.; Pilaczynska-Szczesniak, L.; Nowak, A.; Zielinski, J. Resistin, visfatin and insulin sensitivity in selected phases of annual training cycle of triathletes. Acta Physiol. Hung. 2012, 99, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Nguyen, A.K.; Henstridge, D.C.; Holmes, A.G.; Stanley-Chan, M.H.; Mesa, J.L.; Lancaster, G.I.; Southgate, R.J.; Bruce, C.R.; Duffy, S.J.; et al. HSP72 protects against obesity-induced insulin resistance. Proc. Natl. Acad. Sci. USA 2008, 105, 1739–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, A.A.; Bomhoff, G.L.; Geiger, P.C. Age-related differences in skeletal muscle insulin signaling: The role of stress kinases and heat shock proteins. J. Appl. Physiol. 2008, 105, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.L.; Shike, T.; Shigihara, T.; Tanimoto, M.; Gohda, T.; Makita, Y.; Wang, L.N.; Horikoshi, S.; Tomino, Y. Gene expression profile in diabetic KK/Ta mice. Kidney Int. 2003, 64, 1978–1985. [Google Scholar] [CrossRef] [PubMed]
- Levin, N.; Nelson, C.; Gurney, A.; Vandlen, R.; Sauvage, F.D. Decreased food intake does not completely account for adiposity reduction after ob protein infusion. Proc. Natl. Acad. Sci. USA 1996, 93, 1726–1730. [Google Scholar] [CrossRef] [PubMed]
- Gupte, A.A.; Bomhoff, G.L.; Touchberry, C.D.; Geiger, P.C. Acute heat treatment improves insulin-stimulated glucose uptake in aged skeletal muscle. J. Appl. Physiol. 2011, 110, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, P.C.; Gupte, A.A. Heat shock proteins are important mediators of skeletal muscle insulin sensitivity. Exerc. Sport Sci. Rev. 2011, 39, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.L.; Hopper, P.L. Inflammation, heat shock proteins, and type 2 diabetes. Cell Stress Chaperon. 2009, 14, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.H. Site-specific oxidation of apolipoprotein a-I impairs cholesterol export by ABCA 1, a key cardioprotective function of HDL. BBA-Biomembranes 2012, 1821, 490–501. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Gene | Protein Description | Coverage | Peptides | Theoretial MW/pI | FF/PS | p-Value |
|---|---|---|---|---|---|---|---|
| Q5E9M1 | ARL6IP5 | PRA1 family protein 3 | 18.09 | 3 | 21.6/9.49 | 1.280389 | 0.001434 |
| F1MMU4 | H1FX | H1 Histone Family, Member X | 7.01 | 1 | 22.3/10.71 | 2.019082 | 0.003628 |
| G1DFT2 | RPS4X | Ribosomal Protein S4, X-Linked | 9.89 | 3 | 29.6/10.25 | 1.279684 | 0.0069 |
| Q3T0S6 | RPL8 | 60S ribosomal protein L8 | 16.34 | 4 | 28.0/11.03 | 1.292493 | 0.007701 |
| D7R7V6 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | 42.94 | 14 | 35.8/8.35 | 1.174065 | 0.007889 |
| W5PGZ8 | APOF | Apolipoprotein F | 4.32 | 1 | 35.4/8.06 | 1.546838 | 0.00792 |
| W5PHJ8 | RP | 40S ribosomal protein | 13.42 | 3 | 26.0/9/96 | 1.149951 | 0.008232 |
| A5D9B4 | HNRPH2 | Heterogeneous nuclear ribonucleoprotein H2 | 7.57 | 3 | 49.2/6.30 | 1.158449 | 0.008327 |
| F1MN90 | CIDEC | Cell death-inducing DFFA like effector c | 6.84 | 1 | 27.3/7.97 | 1.224292 | 0.008439 |
| L8IUX9 | M91_16660 | Cytochrome b-c1 complex subunit 8 | 8.54 | 1 | 9.6/10.33 | 1.228296 | 0.009478 |
| F1MXT1 | HACD2 | Very-long-chain (3R)-3-hydroxyacyl-CoA dehydratase | 8.27 | 2 | 28.4/9.56 | 1.331041 | 0.010441 |
| W5QBL2 | CUL1 | Cullin 1 | 1.05 | 1 | 87.4/8.22 | 1.225131 | 0.012333 |
| P20072 | ANXA7 | Annexin A7 | 6.7 | 3 | 49.9/6.80 | 1.252655 | 0.0138 |
| Q9XSK2 | CD63 | CD63 Molecule | 4.2 | 1 | 25.7/7.64 | 1.750327 | 0.01455 |
| W5PED5 | CYP4V2 | CytochromeP450 | 7.78 | 3 | 61.0/7.15 | 1.417145 | 0.016098 |
| A4ZYA6 | VIM | Vimentin | 72.75 | 37 | 53.6/5.09 | 1.370539 | 0.017049 |
| W5NY07 | ATL3 | Atlastin GTPase 3 | 11.5 | 6 | 60.0/5.71 | 1.437302 | 0.018839 |
| Q8WN55 | PTBP1 | Polypyrimidine tract-binding protein 1 | 8.85 | 4 | 57.0/9.17 | 1.142094 | 0.021158 |
| L8I566 | M91_19776 | Coatomer subunit alpha | 1.07 | 1 | 137.7/7.29 | 1.330717 | 0.021314 |
| F4YD28 | RPS24 | 40S ribosomal protein S24 | 7..23 | 1 | 9.5/10.69 | 1.159825 | 0.022228 |
| W5P7F8 | TSPAN | Tetraspanin | 4.24 | 1 | 25.8/8.10 | 1.713756 | 0.022317 |
| W5PK26 | LASP1 | LIM and SH3 Protein 1 | 10 | 2 | 29.6/7.04 | 1.619281 | 0.02251 |
| A8D8X1 | RPL10 | Ribosomal Protein L10 | 18.69 | 5 | 24.5/10.14 | 1.363970 | 0.0233 |
| D3YC70 | HB | Heparin-binding protein | 25 | 1 | 5.6/8.61 | 1.230956 | 0.024758 |
| C3V9V7 | RNP24 | Ribonuclear protein 24 | 7.46 | 2 | 22.7/5.17 | 1.427028 | 0.024858 |
| W5PSJ3 | DCTN1 | Dynactin 1 | 2.62 | 3 | 143.0/5.85 | 1.184554 | 0.025664 |
| F1MBS3 | TGFBI | Transforming growth factor-beta- induced protein ig-h3 | 13.16 | 7 | 72.2/7.12 | 1.500014 | 0.026213 |
| W5NSV5 | ITGA6 | Integrin Alpha 6 | 4.28 | 4 | 106.5/7.43 | 1.246541 | 0.026711 |
| E3SAZ8 | NPM1 | Nucleophosmin | 12.07 | 4 | 32.1/4.85 | 1.283429 | 0.02941 |
| Q307E6 | COL1A1 | Collagen, Type 1, Alpha 1 | 8 | 1 | 15.5/9.12 | 3.591183 | 0.0297 |
| L8IR22 | M91_01904 | Basigin | 16.61 | 4 | 29.7/5.93 | 1.164516 | 0.0299 |
| Q5E971 | TMED10 | Transmembrane emp24 domain- containing protein 10 | 4.11 | 1 | 24.8/6.68 | 1.485596 | 0.03096 |
| W5PIP4 | AZU1 | Azurocidin 1 | 4.45 | 1 | 26.6/11.13 | 1.673528 | 0.031409 |
| Q30B83 | RPL35a | Ribosomal protein L35a | 14 | 1 | 5.8/9.78 | 1.211594 | 0.031542 |
| L8IPP0 | M91_20624 | 78 kDa glucose-regulated protein | 36.03 | 19 | 72.3/5.16 | 1.105556 | 0.031702 |
| W5NSY8 | SH3BGRL | SH3 domain-binding glutamic acid-rich- like protein | 64.65 | 4 | 11.2/5.04 | 1.147976 | 0.031807 |
| C5IJ89 | RHEB | Ras Homolog Enriched In Brain | 7.61 | 2 | 20.4/5.92 | 1.151032 | 0.0318 |
| W5QF34 | LMF2 | Lipase maturation factor | 2.15 | 1 | 72.8/9.96 | 1.289405 | 0.034567 |
| D6BRG0 | ITGB | Integrin, Beta | 2.76 | 2 | 84.3/5.24 | 1.703519 | 0.0359 |
| L8HQV0 | M91_13225 | Dehydrogenase/reductase SDR family member 1 | 8.01 | 2 | 34/8.36 | 1.316582 | 0.03605 |
| A1L555 | PSAP | Prosaposin | 1.9 | 1 | 58.1/5.24 | 1.542171 | 0.03648 |
| W5P5A0 | FLNA | Filamin A, Alpha | 10.82 | 23 | 279.6/6.27 | 1.077701 | 0.0336 |
| L8IBL4 | M91_10571 | Lipoma-preferred partner | 1.91 | 1 | 49.9/7.78 | 1.143675 | 0.037407 |
| K0IVJ6 | metallothionein | Metallothionein | 26.09 | 1 | 4.3/8.23 | 1.870961 | 0.038453 |
| W5PJD0 | PGM2 | Phosphoglucomutase 2 | 1.2 | 1 | 64.9/6.44 | 1.18212 | 0.039518 |
| W5P263 | ABHD5 | alpha/beta hydrolase domain-containing protein 5 | 2.4 | 37.2/8.10 | 1.399659 | 0.040579 | |
| L8HSP5 | M91_02935 | Basic leucine zipper and W2 domain-containing protein 1 | 6.86 | 3 | 48.4/5.77 | 1.529994 | 0.042248 |
| P07514 | CYB5R3 | NADH-cytochrome b5 reductase 3 | 44.19 | 11 | 34.0/7.28 | 1.304232 | 0.042261 |
| W5NVF1 | CYBRD1 | Cytochrome B Reductase 1 | 5.88 | 1 | 22.3/9.27 | 2.219898 | 0.042413 |
| A6QQR5 | TMEM43 | Transmembrane Protein 43 | 9 | 3 | 44.7/8.70 | 1.578646 | 0.044298 |
| W5QHC9 | EPB42 | Erythrocytemembrane protein band 42 | 11.87 | 8 | 77.2/6.85 | 1.263628 | 0.04501 |
| Q3SZF2 | ARF4 | ADP-Ribosylation Factor 4 | 35 | 2 | 20.5/6.18 | 1.683021 | 0.0453 |
| L8IHC9 | M91_15717 | Uncharacterized protein | 11.11 | 1 | 6.2/9.48 | 1.095345 | 0.045529 |
| W5PN85 | ACAT1 | Acyl-coenzyme A:cholesterol acyltrans- ferase | 24.36 | 9 | 45.4/8.55 | 1.382609 | 0.049793 |
| W5Q2N1 | p120GAP | Ras GTPase-activating protein | 1.19 | 1 | 106.0/7.76 | 0.733429 | 0.00144 |
| A0FI82 | LPL | Lipoprotein lipase | 5.65 | 2 | 53.5/8.36 | 0.701299 | 0.004433 |
| W5QA54 | LOC101119629 | Uncharacterized protein | 7.08 | 1 | 22.4/5.31 | 0.273731 | 0.005858 |
| P02510 | CRYAB | Alpha-crystallin B chain | 25.14 | 4 | 20.0/7.32 | 0.749447 | 0.006911 |
| B0BL70 | mbl | Mannose-binding lectin | 4.42 | 1 | 26.5/4.89 | 0.626459 | 0.008106 |
| Q1JPJ2 | XPNPEP1 | Xaa-Pro aminopeptidase 1 | 3.53 | 2 | 69.7/5.68 | 0.860442 | 0.008278 |
| Q2KIL3 | ALAD | Delta-aminolevulinic acid dehydratase | 10.03 | 2 | 36.1/6.99 | 0.570859 | 0.008597 |
| F1N2I5 | CMBL | Carboxymethylenebutenolidase | 8.16 | 2 | 27.9/7.07 | 0.675716 | 0.009004 |
| L8IMU6 | PP2A | Protein Phosphatase 2 Alpha | 8.3 | 1 | 28.7/5.43 | 0.719336 | 0.009013 |
| L8IEF9 | M91_08893 | Vacuolar protein sorting-associated protein 28 homolog | 5.94 | 1 | 22.7/8.42 | 0.656248 | 0.009294 |
| A7XV32 | HSP72 | Heat Shock Protein 72 | 28.24 | 18 | 70.3/5.65 | 0.806279 | 0.010322 |
| A0A0U1YZ59 | ADIPOQ | Adiponectin | 31.8 | 5 | 25.9/6.3 | 0.764493 | 0.0107 |
| H9A6H7 | MSTN | Myostatin | 6.17 | 1 | 9.3/5.17 | 0.294348 | 0.0127 |
| W5PCD1 | LOC101110151 | Uncharacterized protein | 19.15 | 3 | 15.3/5.19 | 0.771917 | 0.013342 |
| F1N5Q0 | ST | Sulfotransferase | 3.14 | 1 | 26.5/8.83 | 0.793002 | 0.013347 |
| W5QGT1 | CUL3 | Uncharacterized protein | 2.68 | 2 | 86.4/7.98 | 0.79521 | 0.013851 |
| B6UV62 | SERPINF1 | SERPINF1 | 18.03 | 7 | 45.9/7.95 | 0.624886 | 0.014795 |
| W5NX51 | APOA1 | Apolipoprotein A-I | 65.64 | 22 | 29.5/6.20 | 0.489784 | 0.015501 |
| L8IVL6 | M91_02549 | Glutamate-cysteine ligase catalytic subunit | 6.3 | 3 | 66.6/5.57 | 0.65182 | 0.015547 |
| L8IZ67 | M91_00380 | Glutaredoxin-3 | 4.19 | 1 | 35.0/6.18 | 0.715634 | 0.015974 |
| Q762I5 | RETN | Resistin | 18.74 | 1 | 5.6/8.69 | 0.799622 | 0.0167 |
| W5QDD0 | LOC101108092 | Uncharacterized protein | 59.83 | 15 | 26.7/7.00 | 0.737769 | 0.017758 |
| Q3T0A3 | CFD | Complement factor D | 67.85 | 2 | 61.1/7.53 | 0.698614 | 0.018044 |
| P29701 | AHSG | Alpha-2-HS-glycoprotein | 46.15 | 12 | 38.6/5.37 | 0.720115 | 0.01812 |
| W5P8E1 | DNPH1 | 2′-deoxynucleoside 5′-phosphate N-hydrolase 1 | 5.75 | 1 | 18.7/5.40 | 0.833179 | 0.020246 |
| Q32LE5 | ASRGL1 | Isoaspartyl peptidase/l-asparaginase | 6.82 | 2 | 32.0/7.40 | 0.730442 | 0.020663 |
| D6PZY4 | fH | Factor H | 9.09 | 10 | 125.2/6.71 | 0.772665 | 0.020943 |
| W5PJ97 | APOA2 | Apolipoprotein A-II | 25.49 | 2 | 11.2/8.10 | 0.456384 | 0.02237 |
| W5Q961 | LOC101107947 | Uncharacterized protein | 7.48 | 2 | 28.3/5.79 | 0.67714 | 0.022512 |
| L8IFD5 | M91_10356 | Guanine nucleotide-binding protein subunit alpha-13 | 5 | 2 | 44.3/7.22 | 0.837336 | 0.024252 |
| W5Q0Y4 | FAH | Fumarylacetoacetase | 12.25 | 5 | 43.9/7.40 | 0.937515 | 0.024511 |
| W5P214 | TAGLN | Transgelin | 40.89 | 8 | 25.0/8.95 | 0.701171 | 0.02459 |
| I1U3B9 | ALDH1L1 | Aldehyde Dehydrogenase 1 Family, Member L1 | 4.08 | 1 | 56.1/7.63 | 0.670099 | 0.0248 |
| Q3SZA0 | COPS4 | COP9 signalosome complex subunit 4 | 11.58 | 4 | 46.2/5.83 | 0.838176 | 0.024938 |
| L8I8L3 | M91_03748 | Vesicle-associated membrane protein 4 | 5.04 | 1 | 16.1/6.04 | 0.475656 | 0.026818 |
| A1L5B0 | MYL9 | Myosin regulatory light polypeptide 9 | 39.88 | 5 | 19.3/4.85 | 0.825417 | 0.028267 |
| W5Q7J0 | APOB | Apolipoprotein B | 7.65 | 30 | 513.8/7.12 | 0.779207 | 0.02886 |
| L8I9W2 | M91_02159 | Tyrosine-protein phosphatase non- receptor type 11 | 3.54 | 2 | 67.8/7.16 | 0.931432 | 0.032163 |
| I1W1N3 | RAB18 | Member RAS Oncogene Family | 5.34 | 1 | 22.9/5.23 | 0.815468 | 0.0336 |
| W5PE22 | GDI2 | Rab GDP dissociation inhibitor | 35.58 | 13 | 48.8/6.97 | 0.841934 | 0.034471 |
| Q28603 | LEP | Leptin | 3.79 | 4 | 16.1/5.71 | 0.792210 | 0.0352 |
| W5Q8X9 | PGP | P-glycoprotein | 5.91 | 1 | 25.1/5.28 | 0.776609 | 0.035498 |
| W5NTW3 | ITIH1 | Inter-alpha-trypsin inhibitor heavy chain H1 | 12.06 | 9 | 101.5/7.26 | 0.530584 | 0.037196 |
| C6ZP47 | HBA1 | I alpha globin | 95.77 | 1 | 15.1/8.67 | 0.508000 | 0.0377 |
| W5PFC9 | LOC101117129 | Uncharacterized protein | 28.09 | 15 | 77.8/6.60 | 0.670015 | 0.039088 |
| W5NZJ1 | LOC101114075 | Sulfotransferase | 12.54 | 4 | 34.3/6.94 | 0.69946 | 0.039392 |
| W5PR48 | HPRT1 | Hypoxanthine-guanine phosphoribosyltransferase 1 | 13.76 | 3 | 24.6/7.09 | 0.708477 | 0.039453 |
| A6H7G2 | DBNL | Drebrin-like protein | 4.73 | 1 | 47.69/4.98 | 0.635779 | 0.041767 |
| L8I9D6 | M91_05702 | B-cell lymphoma 3 protein | 3.66 | 1 | 40.1/7.18 | 0.428236 | 0.041814 |
| W5NR71 | DIAPH1 | Diaphanous 1 | 1.36 | 1 | 138.4/5.51 | 0.764331 | 0.042049 |
| B9VH04 | UQCRH | Cytochrome b-c1 complex subunit 6 | 28.57 | 2 | 10.6/4.50 | 0.811089 | 0.042158 |
| P15497 | APOA1 | Apolipoprotein A-I | 59.25 | 21 | 30.2/5.97 | 0.385227 | 0.04279 |
| Q5E9A6 | VPS25 | Vacuolar protein-sorting-associated protein 25 | 8.52 | 1 | 20.7/6.34 | 0.794093 | 0.044223 |
| W5PU66 | IQGAP1 | IQ motif containing GTP ase activating protein 1 | 8.87 | 12 | 192.8/6.37 | 0.908765 | 0.044485 |
| L8IBI3 | M91_18442 | Terminal uridylyltransferase 7 | 1.08 | 1 | 168.8/7.06 | 0.390906 | 0.044636 |
| W5P6Y6 | AK8 | Adenylatekinase 8 | 5.85 | 1 | 54.7/6.72 | 0.755031 | 0.046026 |
| L8I977 | M91_12085 | Actin-related protein 10 | 3.48 | 1 | 44.7/7.62 | 0.791158 | 0.048268 |
| W5QIC3 | PRUNE | Uncharacterized protein | 4.74 | 1 | 48.8/5.28 | 0.512771 | 0.048663 |
| Gene Name | Gene Ontology, Molecular Function |
|---|---|
| GPIHBP1 | Lipase binding; Lipid binding; Lipoprotein particle binding; |
| AHSG | Glucose, Energy Metabolism; Lipid binding |
| CYP450 | Cholesterol, steroids and other lipids synthesis |
| CYB5R3 | Cholesterol biosynthetic |
| HACD2 | Fatty Acyl-CoA Biosynthesis |
| LPL | Lipoprotein lipase activity; Triglyceride lipase activity; Phospholipase activity |
| Apo-AI | Lipid transporter activity; Phospholipid binding; Phospholipid transporter activity |
| Pathways | Proteins |
|---|---|
| PPAR signaling pathway | Apo-AI, LPL |
| AMPK signaling pathway | PP2A |
| PI3K–Akt signaling pathway | Cdc37, PP2A, ITGB, ITGA |
| Oxidative phosphorylation | UQCRH |
| MAPK signaling pathway | FLNA, HSP72, p120GAP |
| Glycerolipid metabolism | 3.1.1.34 |
| Gene | Accession No. | Primer Sequence (5′→3′) | Product Size (bp) |
|---|---|---|---|
| GAPDH | NM_001190390 | F: CTGACCTGCCGCCTGGAGAAA R: GTAGAAGAGTGAGTGTCGCTGTT | 149 |
| ADIPOQ | KM216385.1 | F: AGTGGTGCCGTCATAGTGG R: CAGTGTAAATGGGGATGTGG | 138 |
| RETN | KJ704841.1 | F: GCAGCACCTGCAGGATGAAG R: GTGGTCTCAGCACGCACGTC | 263 |
| CFD | XM015093908.1 | F: TATCACGACGGCACCATCAC R: TACCGGGCTTCTTGCGGTTG | 160 |
| FLNA | FJ458435.1 | F: CCATCGACGGGCCTTCCAAG R: TGGCAGTCTTGGTCAGGGAG | 234 |
| p120GAP | XM015096185.1 | F: CATCCTGAGTCCACGGATGT R: GGAAGTTCAGGTACATTCCC | 219 |
| PP2A | XM012116023.2 | F: GAGGGAATCATGAAAGCCGT R: CAGACCATAAGAGATCACAC | 261 |
| LEP | XM012177090.2 | F: CCTATCTCTCCTACGTGGAGG R: GGATCTGTTGGTAGATTGCCA | 213 |
| ITGA6 | XM012120035.2 | F: GTGGCCATTCTTGCTGGGAT R: CTATCATCGTACCTAGAGCG | 222 |
| LPL | NM001009394.1 | F: GCTGCTGGTATTGCAGGAAG R: CACTTCACTAGCTGGTCCAC | 308 |
| HSP72 | NM001267874.1 | F: ACCATCCCCACGAAGCAGAC R: CCTCGTCCTCTGCCTTGTAC | 337 |
| Apo-AI | XM012095497.1 | F: GCCCAATTTGAAGCCTCCGC R: TCTGGCGGTAGATCTCCACC | 246 |
| CIDEC | KM199684 | F: ATGGAATACGCCAAGAAGTC R: GGATTGGAAATACCCTTCTG | 300 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Xu, M.; Wang, X.; Yang, J.; Gao, L.; Zhang, Y.; Huang, X.; Han, M.; Gao, R.; Gan, S. Comparative Proteome Analysis Reveals Lipid Metabolism-Related Protein Networks in Response to Rump Fat Mobilization. Int. J. Mol. Sci. 2018, 19, 2556. https://doi.org/10.3390/ijms19092556
Wang J, Xu M, Wang X, Yang J, Gao L, Zhang Y, Huang X, Han M, Gao R, Gan S. Comparative Proteome Analysis Reveals Lipid Metabolism-Related Protein Networks in Response to Rump Fat Mobilization. International Journal of Molecular Sciences. 2018; 19(9):2556. https://doi.org/10.3390/ijms19092556
Chicago/Turabian StyleWang, Juanjuan, Mengsi Xu, Xinhua Wang, Jinquan Yang, Lei Gao, Yan Zhang, Xin Huang, Mengli Han, Rui Gao, and Shangquan Gan. 2018. "Comparative Proteome Analysis Reveals Lipid Metabolism-Related Protein Networks in Response to Rump Fat Mobilization" International Journal of Molecular Sciences 19, no. 9: 2556. https://doi.org/10.3390/ijms19092556