Effects of Different Pollens on Primary Metabolism and Lignin Biosynthesis in Pear

 , ,
, ,
Abstract
:
1. Introduction
2. Results
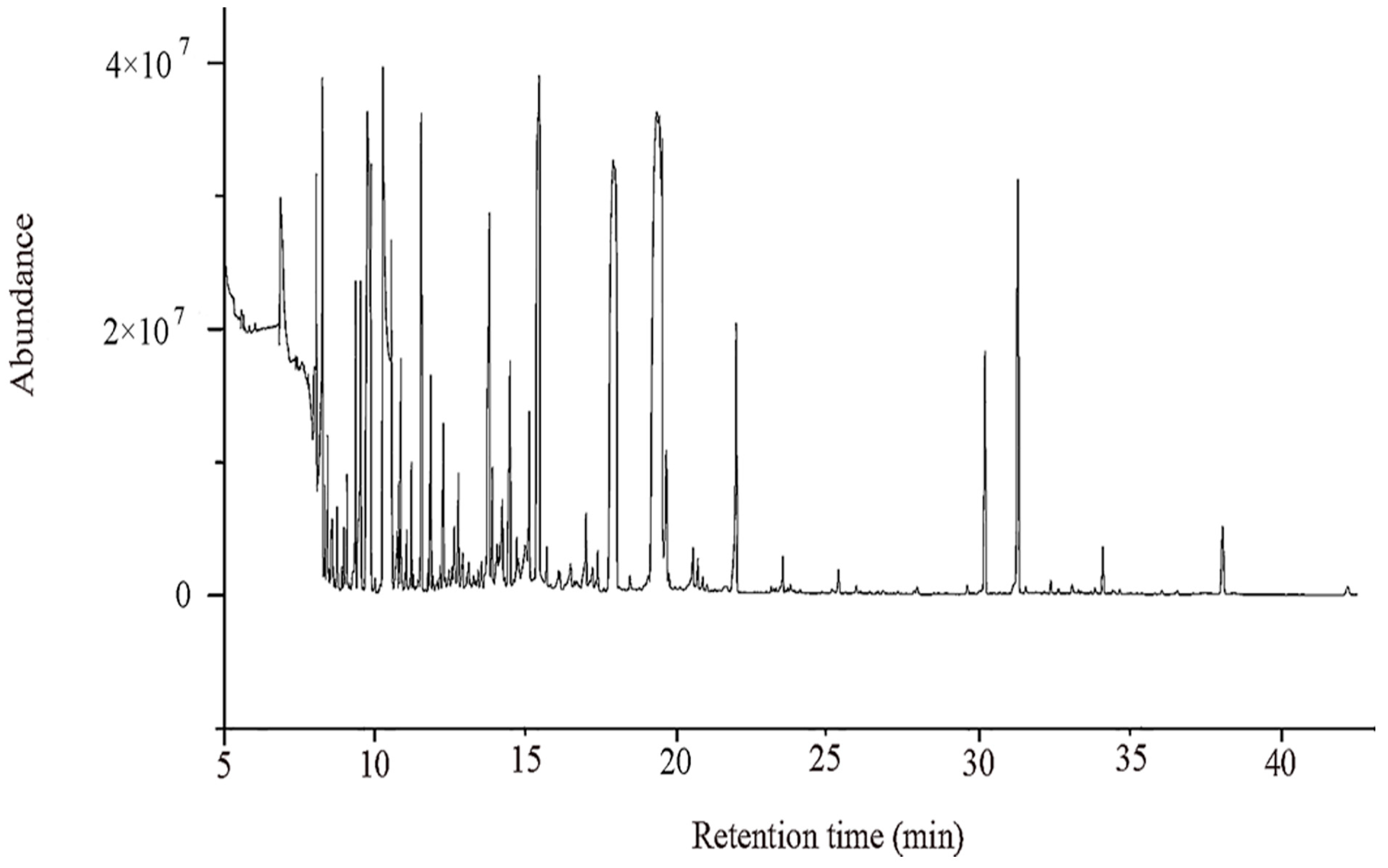
2.1. Un-Targeted Metabolomics Analysis of Primary Metabolites in ‘Dangshan Su’ Pear Fruit with Different Pollens
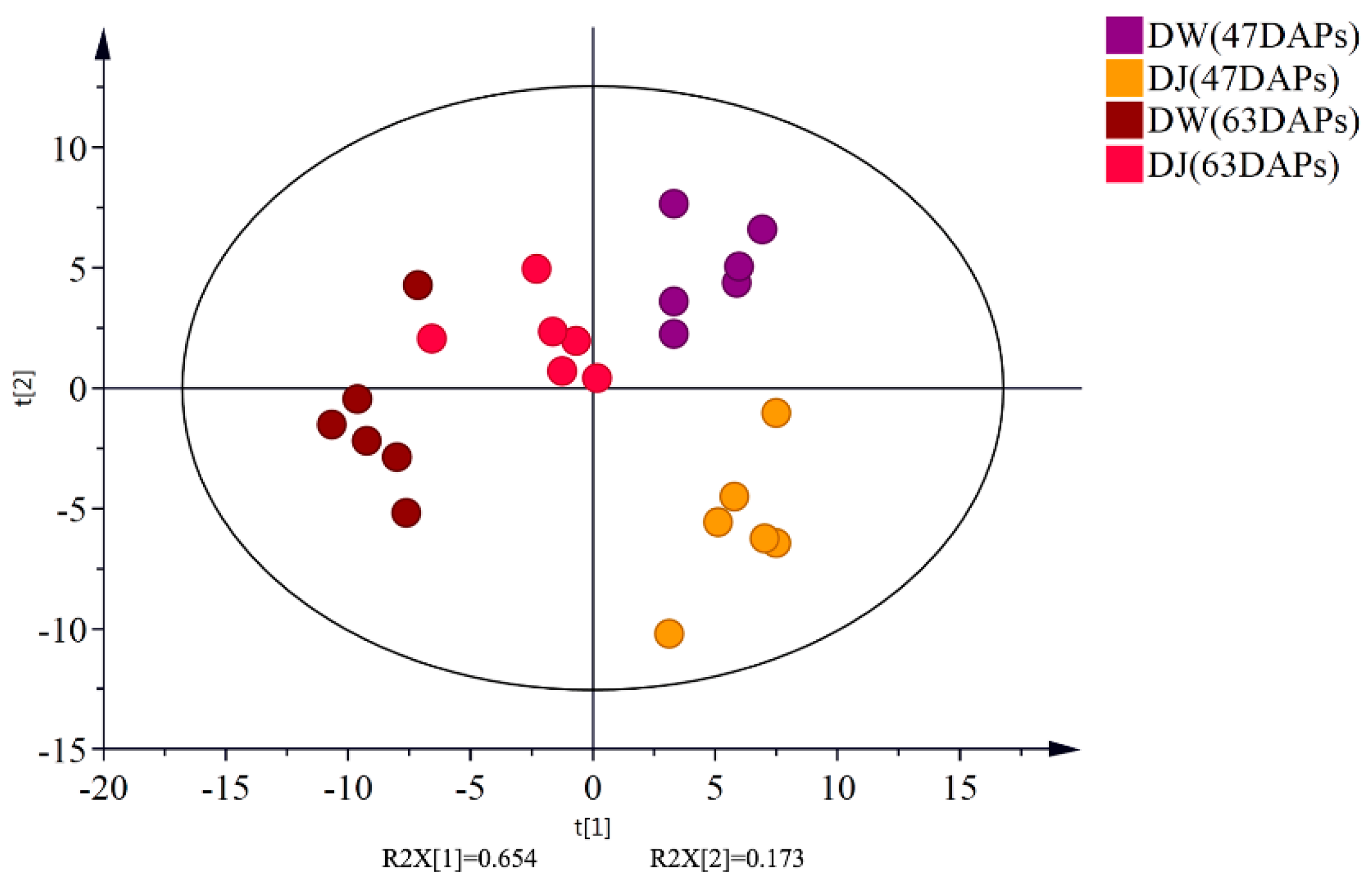
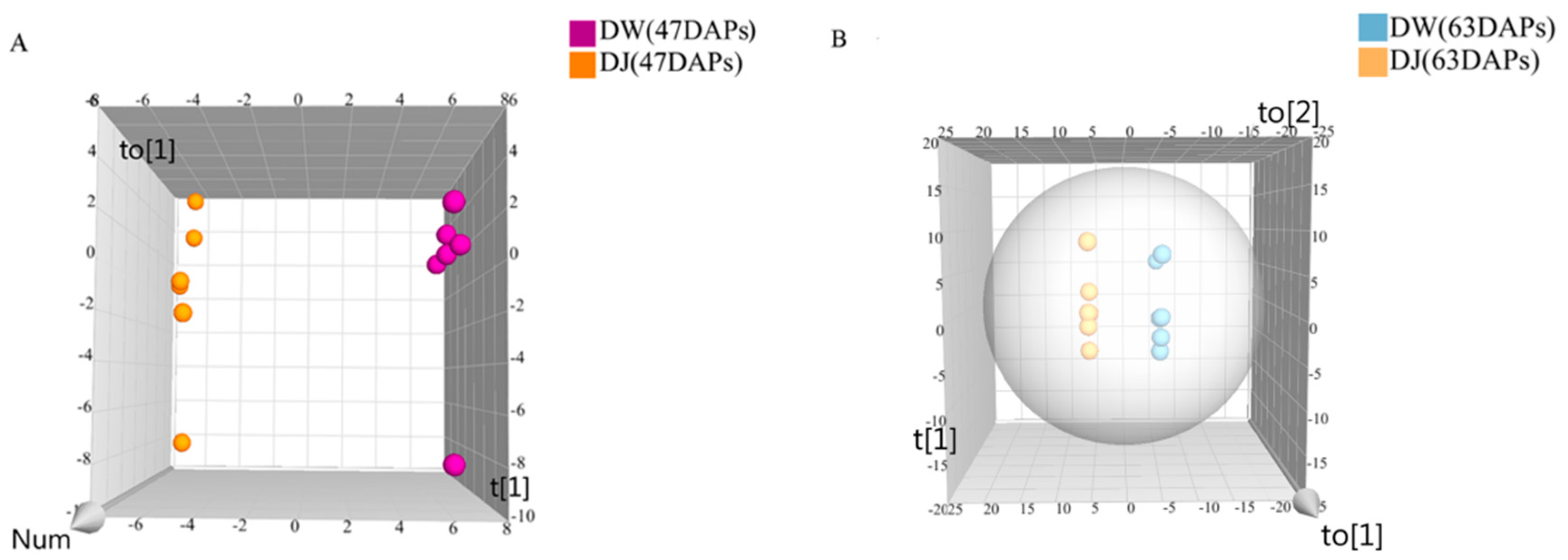
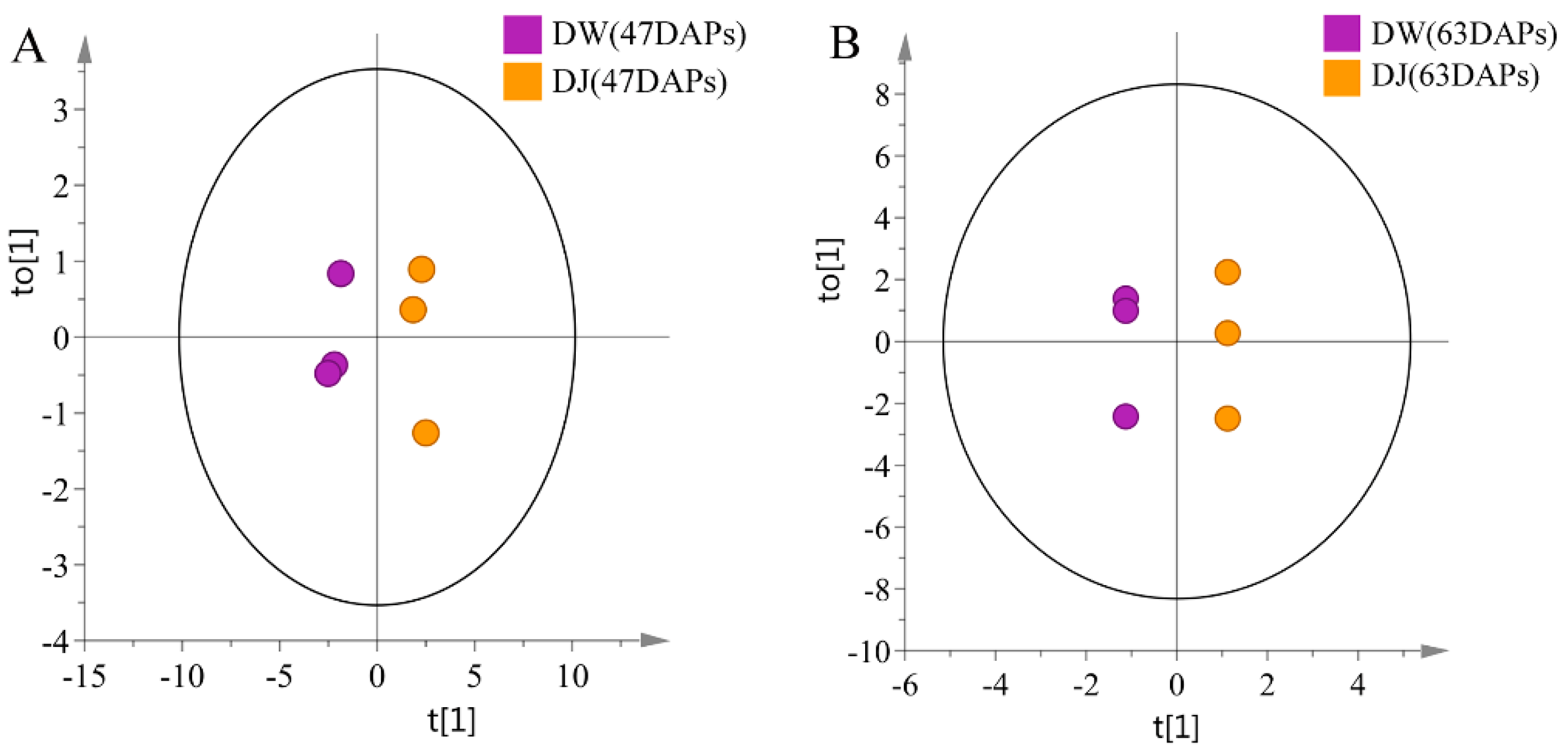
2.1.1. Effect of Pollination on Primary Metabolites in Dangshan Pear Fruit
2.1.2. Effects of Pollination on Carbohydrate Metabolism and Amino Acid Metabolism in Dangshan Pear Fruit
2.2. Effects of Pollination on Intermediate Metabolites of Lignin Synthesis in Dangshan Pear Fruit
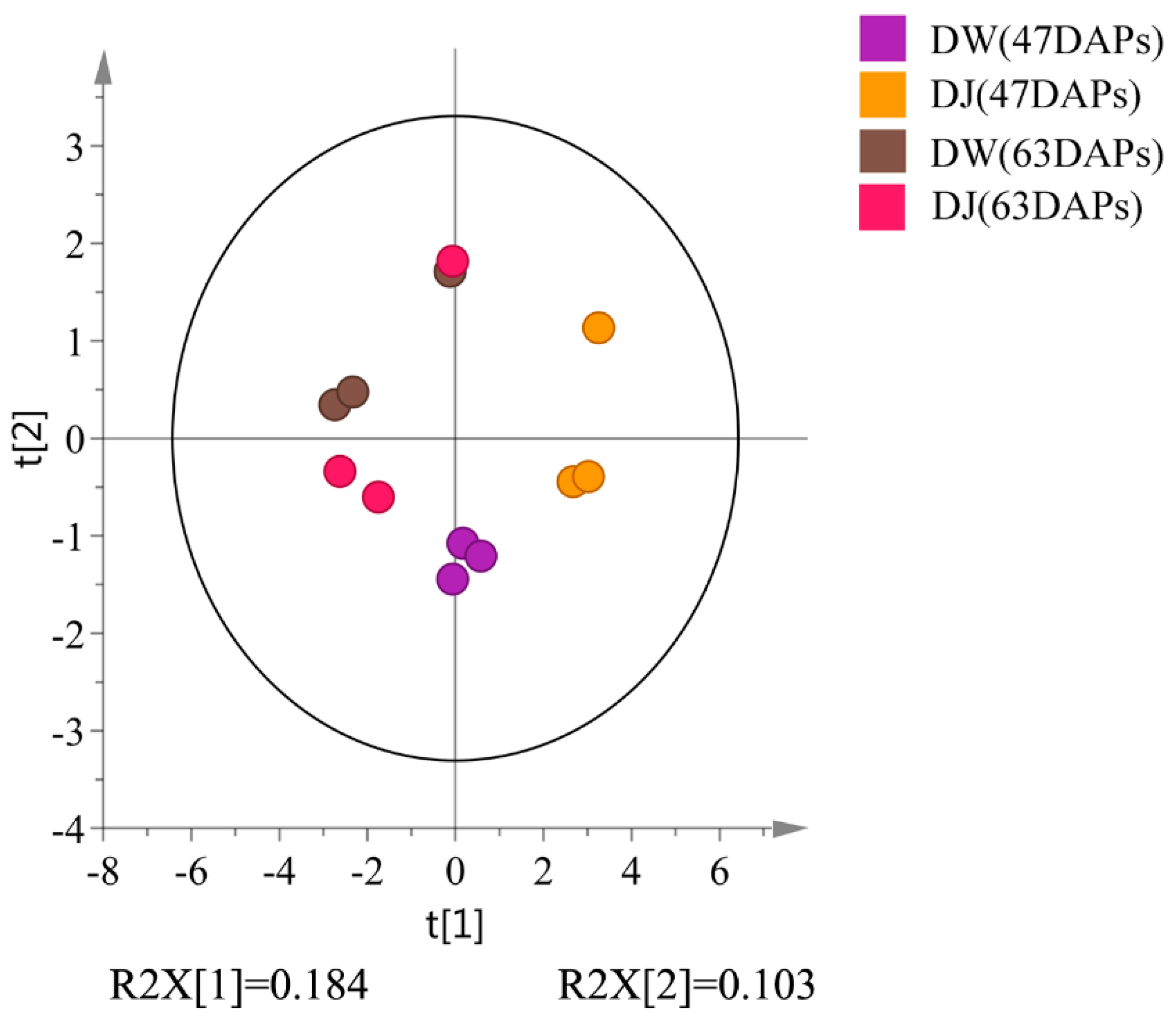
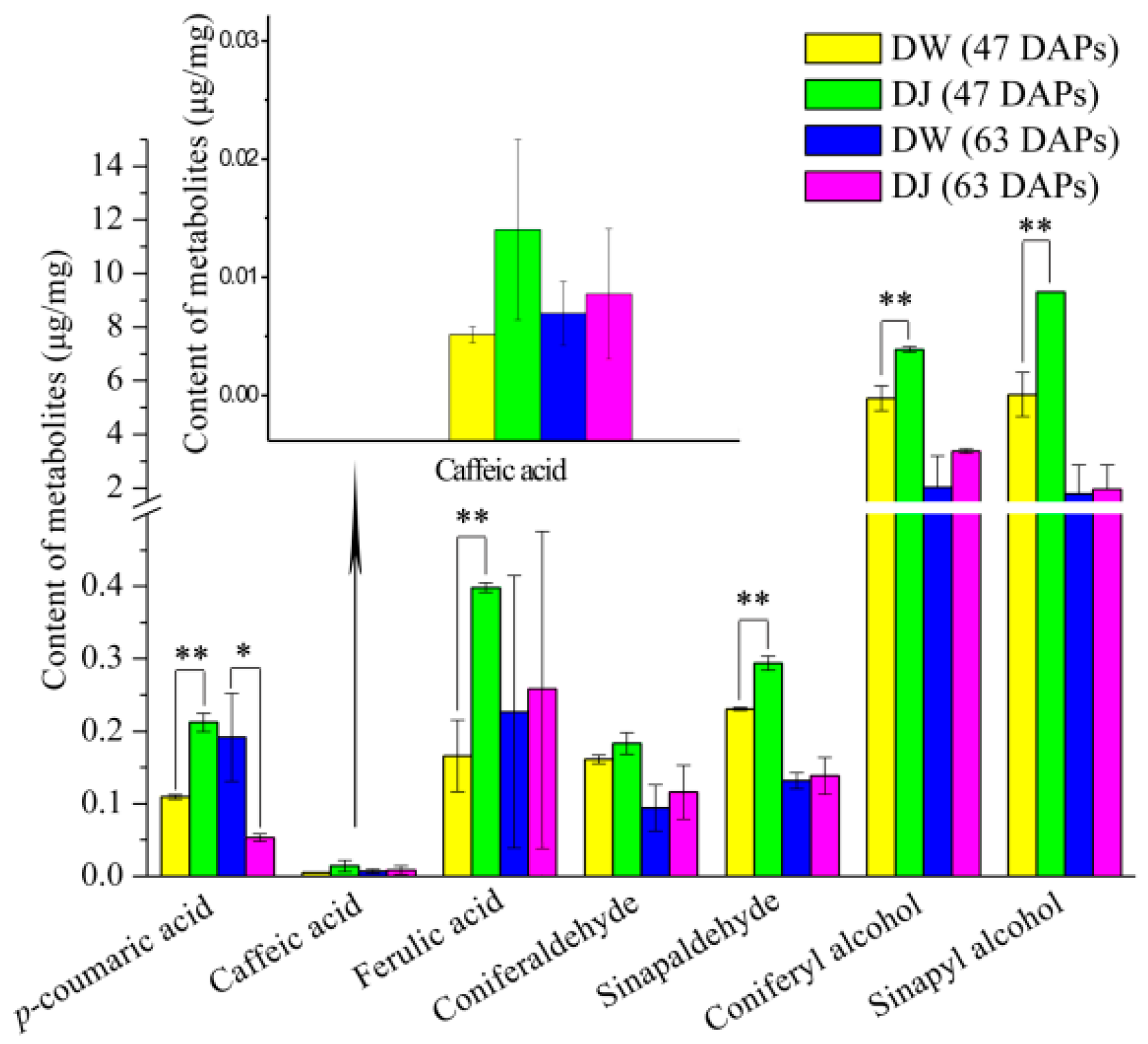
2.2.1. Effects of Pollination on the Content of Intermediate Metabolites in Lignin Synthesis of Dangshan Pear Fruit
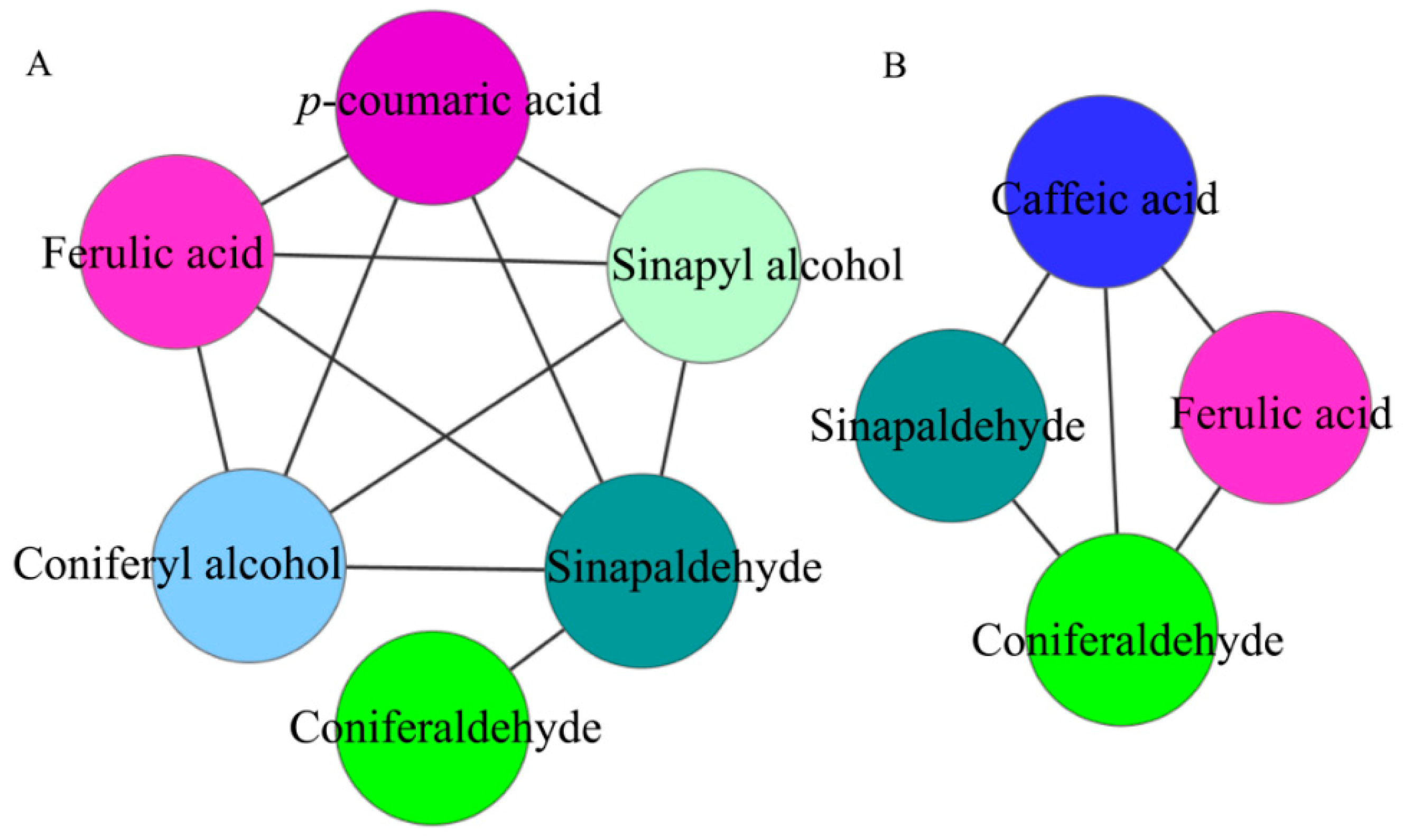
2.2.2. Correlation Analysis of Intermediate Metabolites in Lignin Synthesis in Pear Fruit
3. Discussion and Conclusions
4. Materials and Methods
4.1. Materials
4.2. Preparation and LC-MS Detection of Lignin Synthesis Intermediate Metabolite Standard Solution
4.3. Extraction and Derivation of Primary Metabolites from Pear and Analysis by GC-MS
- (1)
- A 100 mg sample frozen in liquid nitrogen was placed in a 2 mL centrifuge tube, and 1.4 mL of methanol (−20 °C precooled, chromatography grade) was added. The sample was ground with a ball mill and then vortexed for 1 min.
- (2)
- The centrifuge tube was sealed with parafilm and placed in an ultrasonic cleaner at 40 °C for 30 min.
- (3)
- After the end of the ultrasonication, the liquid and the residue was transferred from the 2 mL centrifuge tube to a 10 mL centrifuge tube, and 1.4 mL of purified water (precooled at 4 °C) was added. The sample was then centrifuged at 8000 rpm/min for 15 min.
- (4)
- Then, 100 μL of the upper methanol/water mixture supernatant was placed in a glass flask, and the sample was dried with nitrogen and derivatized. First, 60 μL of hydroxylamine hydrochloride solution (hydroxylamine hydrochloride dissolved in pyridine, 20 mg/mL) was added, and then, the sample was sealed with parafilm, vortexed for 30 s, and placed in an oven at 37 °C for 120 min. Subsequently, 60 μL of derivatizing reagent (99% BSTFA + 1%TMCS) was added, and the flask was sealed and incubated at 37 °C for 90 min. After centrifugation at 12,000 rpm for 4 min, the supernatant was taken in the inner cannula, and the sample was tested. All samples were replicated six times. The test conditions were as follows:
4.4. Extraction and LC-MS Detection of Intermediate Metabolites from Pyrus Lignin
- (1)
- Extraction of phenolic acid material: 100 mg of the samples was taken and frozen in liquid nitrogen. According to the material liquid volume, 80% ethanol solution was added at a 1:15 ratio, and the sample was ground with a ball mill, ultrasonicated for 40 min at 40 °C, and centrifuged to obtain the supernatant. The process was repeated for each group three times.
- (2)
- Phenolic alcohol extract and phenolic substances: 100 mg of each sample was frozen in liquid nitrogen. According to the material liquid volume, 90% ethanol solution was added at a 1:15 ratio, and the sample was ground with a ball mill, ultrasonicated for 30 min at 70 °C, and centrifuged to obtain the supernatant. The process was repeated for each group three times.
4.5. Data Processing and Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Liu, L.; Chen, C.X.; Zhu, Y.F.; Xue, L.; Liu, Q.W.; Qi, K.J.; Zhang, S.L.; Wu, J. Maternal inheritance has impact on organic acid content in progeny of pear (Pyrus spp.) fruit. Euphytica 2016, 209, 305–321. [Google Scholar] [CrossRef]
- Cai, Y.; Li, G.; Nie, J.; Lin, Y.; Nie, F.; Zhang, J.; Xu, Y. Study of the structure and biosynthetic pathway of lignin in stone cells of pear. Sci. Hortic. 2010, 125, 374–379. [Google Scholar] [CrossRef]
- Rosianski, Y.; Freiman, Z.E.; Cochavi, S.M.; Yablovitza, Z.; Kerem, Z.; Flaishman, M.A. Advanced analysis of developmental and ripening characteristics of pollinated common-type fig (Ficus carica L.). Sci. Hortic. 2016, 198, 98–106. [Google Scholar] [CrossRef]
- Cheng, X.; Yan, C.; Zhang, J.; Ma, C.; Li, S.; Jin, Q.; Zhang, N.; Cao, Y.; Lin, Y.; Cai, Y. The Effect of Different Pollination on the Expression of Dangshan Su Pear MicroRNA. BioMed Res. Int. 2017, 2017, 2794040. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, Z.; Wu, J.; Wang, Q.; Hu, X. Chemical compositional characterization of eight pear cultivars grown in China. Food Chem. 2007, 104, 268–275. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, A.; Otsuka, T.; Nakabayashi, R.; Jikumaru, Y.; Isuzugawa, K.; Murayama, H.; Saito, K.; Shiratake, K. Metabolic profiling of developing pear fruits reveals dynamic variation in primary and secondary metabolites, including plant hormones. PLoS ONE 2015, 10, e0131408. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, C.G.; Mansfield, S.D.; Lu, F.; Withers, S.; Park, J.Y.; Karlen, S.D.; Gonzales-Vigil, E.; Padmakshan, D.; Unda, F.; Rencoret, J.; et al. Monolignol ferulate transferase introduces chemically labile linkages into the lignin backbone. Science 2014, 344, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Bunzel, M.; Ralph, J. NMR characterization of lignins isolated from fruit and vegetable insoluble dietary fiber. J. Agric. Food Chem. 2006, 54, 8352–8361. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Yin, M.; Zhang, N.; Jin, Q.; Fang, Z.; Lin, Y.; Cai, Y. Stone cell distribution and lignin structure in various pear varieties. Sci. Hortic. 2014, 174, 142–150. [Google Scholar] [CrossRef]
- Jin, Q.; Yan, C.C.; Qiu, J.; Zhang, N.; Lin, Y.; Cai, Y.P. Structural characterization and deposition of stone cell lignin in Dangshan Su pear. Sci. Hortic. 2013, 155, 123–130. [Google Scholar] [CrossRef]
- Li, S.M.; Su, X.Q.; Jin, Q.; Li, G.H.; Sun, Y.M.; Abdullah, M.; Cai, Y.P.; Lin, Y. iTRAQ-Based Identification of Proteins Related to Lignin Synthesis in the Pear Pollinated with Pollen from Different Varieties. Molecules 2018, 23, 548. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, F.; Ibrahim, B.; Teh, C.H.; Murugaiyah, V.; Chan, K.L. NMR-based plasma metabolomic discrimination for male fertility assessment of rats treated with Eurycoma longifolia extracts. Syst. Biol. Reprod. Med. 2017, 63, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.B.; Salvador, V.H.; Santos, W.D.D.; Bubna, G.A.; Finger-Teixeira, A.; Soares, A.R.; Marchiosi, R.; Mde, L.F.; Ferrarese-Filho, O. Enhanced Lignin Monomer Production Caused by Cinnamic Acid and Its Hydroxylated Derivatives Inhibits Soybean Root Growth. PLoS ONE 2013, 8, e80542. [Google Scholar] [CrossRef] [PubMed]
- Shivashankar, S.; Sumathi, M.; Krishnakumar, N.K.; Rao, V.K. Role of phenolic acids and enzymes of phenylpropanoid pathway in resistance of chayote fruit (Sechium edule) against infestation by melon fly, Bactrocera cucurbitae. Ann. Appl. Biol. 2015, 166, 420–433. [Google Scholar] [CrossRef]
- Zheng, K.; Shen, N.; Chen, H.; Ni, S.; Zhang, T.; Hu, M.; Wang, J.; Sun, L.; Yang, X. Global and targeted metabolomics of synovial fluid discovers special osteoarthritis metabolites. J. Orthop. Res. 2017, 35, 1973–1981. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Peng, J.S.; Yao, D.S.; Yang, Z.L.; Liu, H.L.; Zeng, Y.K.; Shi, X.P.; Lu, B.Y. Serum metabolic profiling of human gastric cancer based on gas chromatography/mass spectrometry. Braz. J. Med. Biol. Res. 2012, 45, 78–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, X.; Zhong, X.; Tian, X. Metabolomics of papillary thyroid carcinoma tissues: Potential biomarkers for diagnosis and promising targets for therapy. Tumor Biol. 2016, 37, 11163–11175. [Google Scholar] [CrossRef] [PubMed]
- Cumming, G.; Fidler, F.; Vaux, D.L. Error bars in experimental biology. J. Cell Biol. 2007, 177, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement? Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef] [PubMed]
- De Souza, L.P.; Naake, T.; Tohge, T.; Fernie, A.R. From chromatogram to analyte to metabolite. How to pick horses for courses from the massive web-resources for mass spectral plant metabolomics. Gigascience 2017, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Rist, M.J.; Roth, A.; Frommherz, L.; Weinert, C.H.; Krüger, R.; Merz, B.; Bunzel, D.; Mack, C.; Egert, B.; Bub, A.; et al. Metabolite patterns predicting sex and age in participants of the Karlsruhe Metabolomics and Nutrition (KarMeN) study. PLoS ONE 2017, 12, e0183228. [Google Scholar] [CrossRef] [PubMed]
- Georgita, C.; Sora, I.; Albu, F.; Monciu, C.M. Comparison of a LC/MS method with a LC/UV method for the determination of metformin in plasma samples. Farmacia 2010, 58, 158–169. [Google Scholar]
- Shehab, M.A.; Biggar, K.; Singal, S.S.; Nygard, K.; Shun-Cheng, L.S.; Jansson, T.; Gupta, M.B. Exposure of decidualized HIESC to low oxygen tension and leucine deprivation results in increased IGFBP-1 phosphorylation and reduced IGF-I bioactivity. Mol. Cell. Endocrinol. 2017, 452, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, N.A.; Tobimatsu, Y.; Ciesielski, P.N.; Ximenes, E.; Ralph, J.; Donohoe, B.S.; Ladisch, M.; Chapple, C. Manipulation of guaiacyl and syringyl monomer biosynthesis in an Arabidopsis cinnamyl alcohol dehydrogenase mutant results in atypical lignin biosynthesis and modified cell wall structure. Plant Cell 2015, 27, 2195–2209. [Google Scholar] [CrossRef] [PubMed]
- Zanardo, D.I.L.; Lima, R.B.; Ferrarese, M.L.L.; Bubna, G.A.; Ferrarese-Filho, O. Soybean root growth inhibition and lignification induced by p-coumaric acid. Environ. Exp. Bot. 2009, 66, 25–30. [Google Scholar] [CrossRef]
- Bunsiri, A.; Ketsa, S.; Paull, R.E. Phenolic metabolism and lignin synthesis in damaged pericarp of mangosteen fruit after impact. Postharvest Biol. Technol. 2003, 29, 61–71. [Google Scholar] [CrossRef]
- Tao, S.T.; Khanizadeh, S.; Zhang, H.; Zhang, S.L. Anatomy, ultrastructure and lignin distribution of stone cells in two Pyrus species. Plant Sci. 2009, 176, 413–419. [Google Scholar] [CrossRef]
- Brahim, S.B.; Kelebek, H.; Ammar, S.; Abichou, M.; Bouaziz, M. LC-MS phenolic profiling combined with multivariate analysis as an approach for the characterization of extra virgin olive oils of four rare Tunisian cultivars during ripening. Food Chem. 2017, 229, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Faccin, H.; Loose, R.F.; Viana, C.; de Carvalho, L.M. Determination of phenolic compounds in extracts of Amazonian medicinal plants by liquid chromatography-electrospray tandem mass spectrometry. Anal. Methods 2017, 9, 1141–1151. [Google Scholar] [CrossRef]
- Gajbhiye, N.A.; Makasana, J.; Saha, A.; Jat, R.S. LC-ESI-MS/MS Method for Simultaneous Determination of Triterpenoid Glycosides and Aglycones in Centella asiatica L. Chromatographia 2016, 79, 727–739. [Google Scholar] [CrossRef]
- Rehman, S.U.; Choi, M.S.; Kim, I.S.; Kim, S.H.; Yoo, H.H. An ultra-high-performance liquid chromatography-tandem mass spectrometric method for the determination of hederacoside C, a drug candidate for respiratory disorder, in rat plasma. J. Pharm. Biomed. Anal. 2016, 129, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Roessner, U.; Wagner, C.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Simultaneous analysis of metabolites in potato tuber by gas chromatography–mass spectrometry. Plant J. 2000, 23, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C.Y.; Song, C.; Zheng, S.Y.; Zhu, Y.P.; Jin, Q.; Cai, Y.P.; Lin, Y. Metabolic Profiling of Dendrobium officinale in Response to Precursors and Methyl Jasmonate. Int. J. Mol. Sci. 2018, 19, 728. [Google Scholar] [CrossRef] [PubMed]
- De la Barca, J.M.C.; Huang, N.T.; Jiao, H.; Tessier, L.; Gadras, C.; Simard, G.; Natoli, R.; Tcherkez, G.; Reynier, P.; Valter, K. Retinal metabolic events in preconditioning light stress as revealed by wide-spectrum targeted metabolomics. Metabolomics 2017, 13, 22. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | VIP | p-Value | Fold Change |
|---|---|---|---|
| 3-hydroxypyruvate | 1.959 | 2.760 × 10−19 | + |
| l-kynurenine | 1.958 | 4.438 × 10−16 | + |
| S-carboxymethylcysteine | 1.875 | 2.322 × 10−6 | 3.374 |
| aspartic acid | 1.849 | 4.036 × 10−6 | 0.385 |
| Cumic Acid | 1.839 | 9.472 × 10−6 | 3.197 |
| valine | 1.835 | 1.009 × 10−5 | 0.394 |
| Cysteinylglycine | 1.832 | 2.973 × 10−5 | 3.979 |
| 5,6-dihydrouracil | 1.824 | 2.572 × 10−5 | 2.963 |
| proline | 1.812 | 2.147 × 10−5 | 0.359 |
| adenosine | 1.793 | 2.837 × 10−5 | 0.513 |
| oxoproline | 1.786 | 5.476 × 10−5 | 0.469 |
| malonic acid | 1.778 | 2.021 × 10−4 | 7.683 |
| glycine | 1.776 | 3.924 × 10−5 | 0.536 |
| Maleamate | 1.769 | 1.128 × 10−4 | 1.633 |
| stearic acid | 1.733 | 2.268 × 10−4 | 1.890 |
| catechol | 1.730 | 1.458 × 10−4 | 0.466 |
| Adipamide | 1.706 | 1.866 × 10−4 | 0.302 |
| palmitic acid | 1.704 | 4.068 × 10−4 | 1.719 |
| maltotriose | 1.703 | 2.545 × 10−4 | 0.573 |
| Linoleic acid methyl ester | 1.687 | 1.359 × 10−3 | 2.320 |
| linolenic acid | 1.682 | 9.211 × 10−4 | 1.930 |
| MALONAMIDE | 1.649 | 9.539 × 10−4 | 1.735 |
| linoleic acid | 1.639 | 2.797 × 10−3 | 1.646 |
| creatine degr | 1.620 | 1.159 × 10−3 | 1.702 |
| Elaidic acid | 1.563 | 4.451 × 10−3 | 1.568 |
| sorbitol | 1.559 | 2.184 × 10−3 | 1.583 |
| glucose-6-phosphate | 1.557 | 3.682 × 10−3 | 1.850 |
| Methyl Palmitoleate | 1.483 | 0.013 | 1,989 |
| Monostearin | 1.457 | 7.202 × 10−3 | 1.391 |
| d-(glycerol 1-phosphate) | 1.452 | 5.2941 × 10−3 | 0.687 |
| caffeic acid | 1.444 | 7.203 × 10−3 | 3.383 |
| Phenylphosphoric acid | 1.435 | 7.398 × 10−3 | 1.995 |
| Maleimide | 1.429 | 0.015 | 1.519 |
| 1-Monopalmitin | 1.416 | 9.943 × 10−3 | 1.410 |
| thymine | 1.413 | 0.013 | 24.224 |
| hexadecane | 1.408 | 0.010 | + |
| 2-amino-2-methylpropane-1,3-diol | 1.378 | 0.010 | + |
| Nicotinoylglycine | 1.370 | 0.011 | + |
| Dioctyl phthalate | 1.358 | 0.019 | 0.552 |
| lysine | 1.351 | 0.017 | + |
| 3-Hydroxyanthranilic acid | 1.347 | 0.026 | 0.584 |
| shikimic acid | 1.330 | 0.016 | 1.446 |
| arbutin | 1.303 | 0.019 | 1.705 |
| asparagine | 1.291 | 0.019 | + |
| maltitol | 1.280 | 0.024 | 0.661 |
| Glucose-1-phosphate | 1.269 | 0.028 | 0.599 |
| N-epsilon-Acetyl-l-lysine | 1.234 | 0.038 | 0.682 |
| Synephrine | 1.206 | 0.034 | 0.674 |
| 3-Methyloxindole | 1.199 | 0.045 | 2.544 |
| Alizarin | 1.198 | 0.041 | 0.370 |
| Mono(2-ethylhexyl) phthalate | 1.184 | 0.050 | + |
| Metabolite | VIP | p-Value | Fold Change |
|---|---|---|---|
| Leucrose | 1.918 | 5.181 × 10−14 | - |
| Valine | 1.907 | 2.145 × 10−10 | 5.355 |
| Myo-inositol | 1.766 | 4.069 × 10−5 | 0.682 |
| Palatinitol | 1.751 | 1.052 × 10−4 | 0.628 |
| Sorbitol | 1.720 | 1.402 × 10−4 | 2.293 |
| Cellobiotol | 1.702 | 2.840 × 10−4 | 0.622 |
| Mannitol | 1.651 | 7.616 × 10−4 | 0.478 |
| Maltotriose | 1.650 | 1.137 × 10−3 | 0.647 |
| 1-Methyladenosine | 1.649 | 4.547 × 10−4 | - |
| Lysine | 1.646 | 5.772 × 10−4 | - |
| Aspartic acid | 1.643 | 5.372 × 10−4 | 0.557 |
| 2-Amino-2-methylpropane-1,3-diol | 1.626 | 6.194 × 10−4 | - |
| Elaidic acid | 1.626 | 1.583 × 10−3 | 0.677 |
| Glycine | 1.625 | 1.313 × 10−3 | 1.396 |
| Linoleic acid | 1.558 | 4.088 × 10−3 | 0.629 |
| Oxoproline | 1.549 | 2.818 × 10−3 | 0.690 |
| Galactinol | 1.540 | 0.590 × 10−3 | 0.579 |
| Sitosterol | 1.510 | 6.632 × 10−3 | 0.702 |
| N-epsilon-Acetyl-l-lysine | 1.479 | 8.339 × 10−3 | 0.808 |
| β-Mannosylglycerate | 1.453 | 6.939 × 10−3 | 0.649 |
| Ribitol | 1.449 | 0.015 | 0.669 |
| Adipamide | 1.435 | 6.447 × 10−3 | 1.396 |
| Fructose | 1.423 | 0.010 | 0.615 |
| Proline | 1.411 | 0.017 | 0.472 |
| Vanillylmandelic acid | 1.386 | 0.019 | 0.715 |
| Heptadecanoic acid | 1.380 | 0.019 | 0.780 |
| Gallic acid | 1.379 | 0.024 | 0.605 |
| Citramalic acid | 1.346 | 0.016 | + |
| 3-Methyloxindole | 1.288 | 0.025 | 0.727 |
| d-Glyceric acid | 1.272 | 0.045 | 1.384 |
| 3-Hydroxypropionic acid | 1.211 | 0.043 | 0.566 |
| Hydantoin, 5-(4-hydroxybutyl)- | 1.194 | 0.049 | - |
| Sorbose | 1.196 | 0.050 | - |
| 2-Indanone | 1.177 | 0.050 | - |
| KEGG Pathway | p-Value | FDR |
|---|---|---|
| Glycerophospholipid metabolism | 5.330 × 10−8 | 9.592 × 10−7 |
| Glycerolipid metabolism | 5.330 × 10−8 | 9.592 × 10−7 |
| α-Linolenic acid metabolism | 2.098 × 10−7 | 2.518 × 10−6 |
| Glutathione metabolism | 6.650 × 10−7 | 2.946 × 10−6 |
| Biosynthesis of unsaturated fatty acids | 8.173 × 10−7 | 2.946 × 10−6 |
| Pyrimidine metabolism | 8.620 × 10−7 | 2.946 × 10−6 |
| Pantothenate and CoA biosynthesis | 8.805 × 10−7 | 2.946 × 10−6 |
| Fatty acid biosynthesis | 1.010 × 10−6 | 2.946 × 10−6 |
| Fatty acid elongation in mitochondria | 1.080 × 10−6 | 2.946 × 10−6 |
| Fatty acid metabolism | 1.080 × 10−6 | 2.946 × 10−6 |
| β-Alanine metabolism | 1.124 × 10−6 | 2.946 × 10−6 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 1.130 × 10−6 | 2.946 × 10−6 |
| Fructose and mannose metabolism | 1.191 × 10−6 | 2.946 × 10−6 |
| Valine, leucine and isoleucine biosynthesis | 1.309 × 10−6 | 2.946 × 10−6 |
| Valine, leucine and isoleucine degradation | 1.309 × 10−6 | 2.946 × 10−6 |
| Glucosinolate biosynthesis | 1.309 × 10−6 | 2.946 × 10−6 |
| Arginine and proline metabolism | 1.802 × 10−6 | 3.280 × 10−6 |
| Cysteine and methionine metabolism | 1.823 × 10−6 | 3.280 × 10−6 |
| Carbon fixation in photosynthetic organisms | 1.823 × 10−6 | 3.280 × 10−6 |
| Nicotinate and nicotinamide metabolism | 1.823 × 10−6 | 3.280 × 10−6 |
| Cyanoamino acid metabolism | 1.968 × 10−6 | 3.374 × 10−6 |
| Galactose metabolism | 2.552 × 10−6 | 4.18 × 10−6 |
| Tryptophan metabolism | 4.674 × 10−6 | 7.32 × 10−6 |
| Methane metabolism | 7.196 × 10−6 | 1.04 × 10−5 |
| Nitrogen metabolism | 7.196 × 10−6 | 1.04 × 10−5 |
| Glycine, serine and threonine metabolism | 2.417 × 10−5 | 3.35 × 10−5 |
| Starch and sucrose metabolism | 1.051 × 10−4 | 1.261 × 10−4 |
| Amino sugar and nucleotide sugar metabolism | 1.051 × 10−4 | 1.261 × 10−4 |
| Glycolysis or Gluconeogenesis | 1.051 × 10−4 | 1.261 × 10−4 |
| Pentose and glucuronate interconversions | 1.051 × 10−4 | 1.261 × 10−4 |
| Lysine biosynthesis | 6.498 × 10−3 | 7.546 × 10−3 |
| Metabolite | 47 DAPs | 63 DAPs | ||
|---|---|---|---|---|
| VIP | p-Value | VIP | p-Value | |
| p-coumaric acid | 1.08325 | 0.00018 ** | 1.90639 | 0.01701 * |
| Caffeic acid | 0.776344 | 0.11483 | 0.483629 | 0.66653 |
| Ferulic acid | 1.063 | 0.00129 ** | 0.182974 | 0.87196 |
| Coniferaldehyde | 0.839268 | 0.07556 | 0.766778 | 0.48518 |
| Sinapaldehyde | 1.07733 | 0.00040 ** | 0.444275 | 0.69285 |
| Coniferyl alcohol | 1.04866 | 0.00268 ** | 1.50585 | 0.11830 |
| Sinapyl alcohol | 1.06264 | 0.00132 ** | 0.213007 | 0.85107 |
| Metabolite | Ion Mode | Precursor Ion (m/z) | Product Ion (m/z) | DP | CE |
|---|---|---|---|---|---|
| Cinnamic acid | - | 146.9 | 103.0 | −30 | −13 |
| p-coumaric acid | - | 162.8 | 119.0 | −30 | −15.5 |
| Caffeic acid | - | 179.0 | 135.0 | −30 | −20 |
| Ferulic acid | - | 193.0 | 134.0 | −30 | −20 |
| 149.1 | −30 | −13 | |||
| 116.9 | −30 | −20 | |||
| Sinapic acid | - | 223.0 | 148.9 | −30 | −17 |
| 164.0 | −30 | −18 | |||
| Coniferaldehyde | - | 177.0 | 162.0 | −30 | −17 |
| 134.0 | −30 | −28 | |||
| Sinapaldehyde | - | 207.0 | 192.0 | −24 | −17 |
| 176.8 | −30 | −27 | |||
| 149.2 | −30 | −35 | |||
| Coniferyl alcohol | - | 179.1 | 146.0 | −30 | −19 |
| 164.0 | −30 | −18 | |||
| 161 | −30 | −13 | |||
| Sinapyl alcohol | - | 209.0 | 194.0 | −30 | −17 |
| 179.0 | −40 | −24 | |||
| 176.1 | −40 | −18 |
| Metabolite | Chemical Formula | Formula Weight | Standard Curve | Correlation Coefficient | Retention Time (min) |
|---|---|---|---|---|---|
| Cinnamic acid | C9H8O2 | 148.161 | Y = 1.12 × 103X + 4.35 × 104 | R2 = 1.0000 | 11.8 |
| p-coumaric acid | C9H8O3 | 164.160 | Y = 2.36 × 104X + 2.29 × 105 | R2 = 0.9994 | 10.4 |
| Caffeic acid | C9H8O4 | 180.159 | Y = 4.57 × 104X − 1.33 × 105 | R2 = 0.9995 | 9.9 |
| Ferulic acid | C10H10O4 | 194.186 | Y = 8.38 × 103X + 6.2 × 103 | R2 = 0.9999 | 10.5 |
| Sinapic acid | C11H12O5 | 224.212 | Y = 455X − 603 | R2 = 0.9992 | 10.3 |
| Coniferaldehyde | C10H10O3 | 178.187 | Y = 2.38 × 104X + 6.08 × 104 | R2 = 0.9998 | 11.0 |
| Sinapaldehyde | C11H12O4 | 208.213 | Y = 1.22 × 104X − 2.55 × 104 | R2 = 0.9999 | 10.9 |
| Coniferyl alcohol | C10H12O3 | 180.203 | Y = 98.4X − 519 | R2 = 0.9998 | 10.3 |
| Sinapyl alcohol | C11H14O4 | 210.229 | Y = 150X − 776 | R2 = 0.9998 | 10.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Su, X.; Abdullah, M.; Sun, Y.; Li, G.; Cheng, X.; Lin, Y.; Cai, Y.; Jin, Q. Effects of Different Pollens on Primary Metabolism and Lignin Biosynthesis in Pear. Int. J. Mol. Sci. 2018, 19, 2273. https://doi.org/10.3390/ijms19082273
Li S, Su X, Abdullah M, Sun Y, Li G, Cheng X, Lin Y, Cai Y, Jin Q. Effects of Different Pollens on Primary Metabolism and Lignin Biosynthesis in Pear. International Journal of Molecular Sciences. 2018; 19(8):2273. https://doi.org/10.3390/ijms19082273
Chicago/Turabian StyleLi, Shumei, Xueqiang Su, Muhammad Abdullah, Yanming Sun, Guohui Li, Xi Cheng, Yi Lin, Yongping Cai, and Qing Jin. 2018. "Effects of Different Pollens on Primary Metabolism and Lignin Biosynthesis in Pear" International Journal of Molecular Sciences 19, no. 8: 2273. https://doi.org/10.3390/ijms19082273
APA StyleLi, S., Su, X., Abdullah, M., Sun, Y., Li, G., Cheng, X., Lin, Y., Cai, Y., & Jin, Q. (2018). Effects of Different Pollens on Primary Metabolism and Lignin Biosynthesis in Pear. International Journal of Molecular Sciences, 19(8), 2273. https://doi.org/10.3390/ijms19082273