Chrysanthemum DgWRKY2 Gene Enhances Tolerance to Salt Stress in Transgenic Chrysanthemum

Abstract
1. Introduction
2. Results
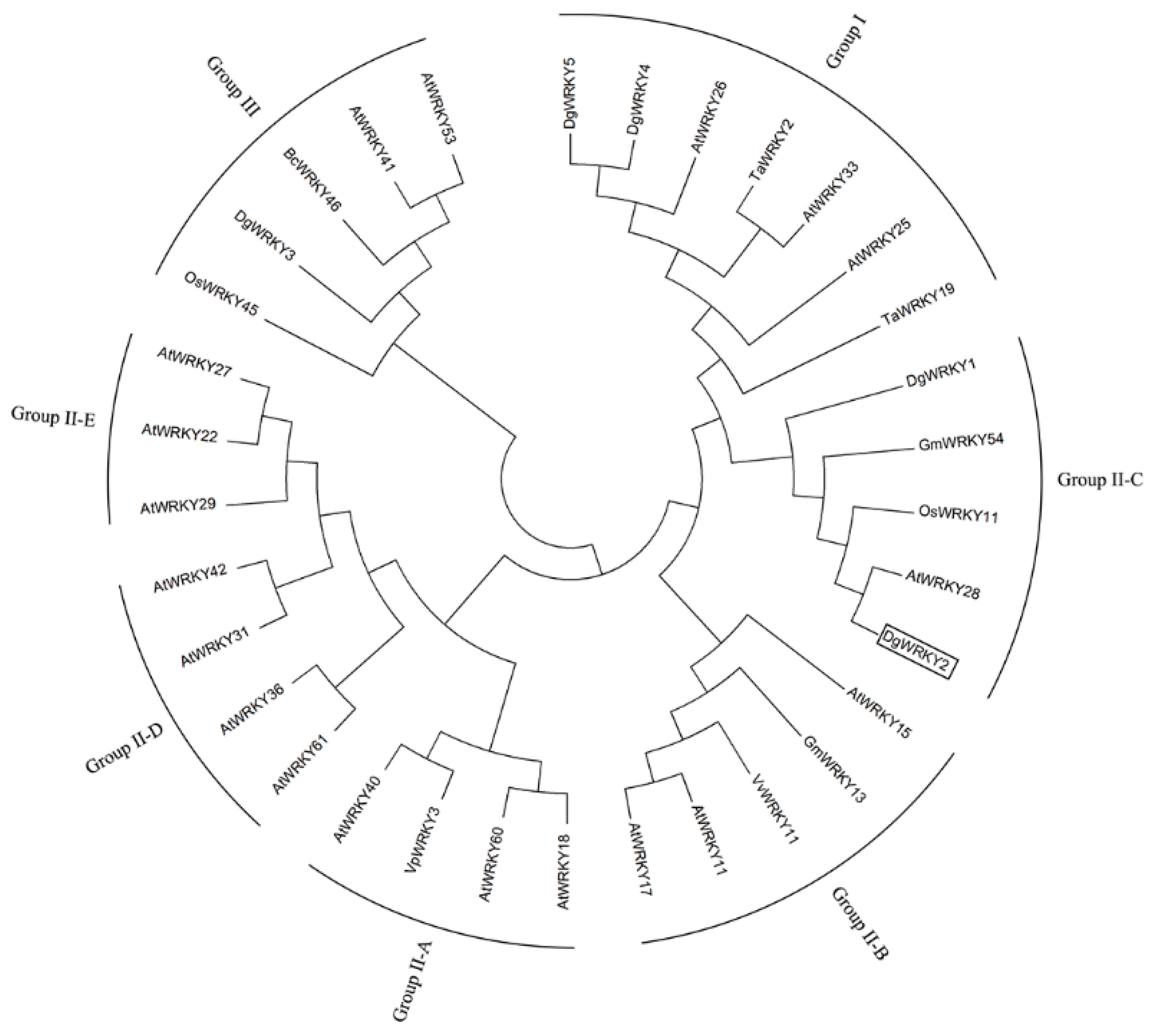
2.1. Isolation and Characterization of DgWRKY2
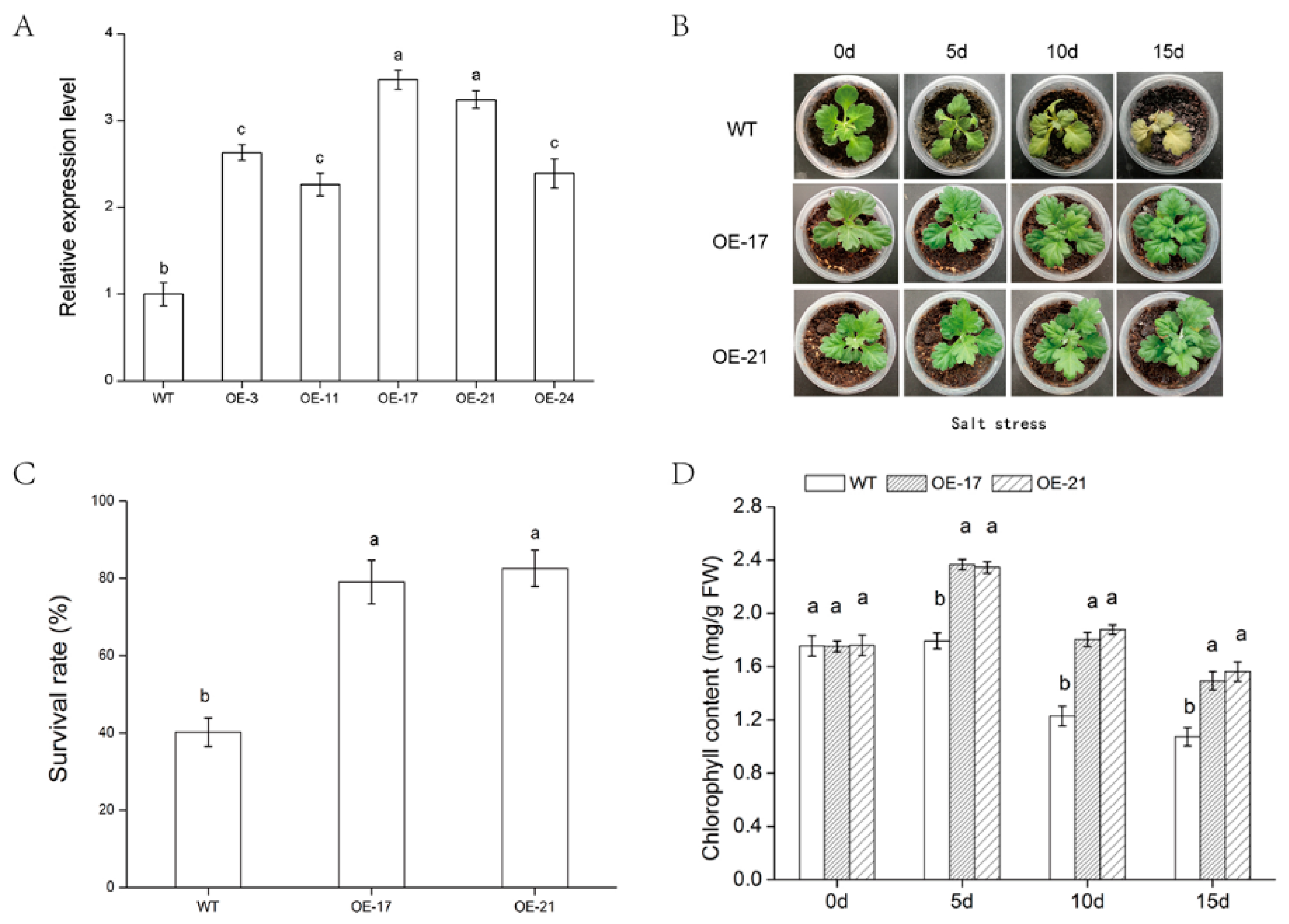
2.2. Salt-Tolerance Analysis of DgWRKY2 Transgenic Chrysanthemum
2.3. Analysis of Chlorophyll Content and under Salt Stress
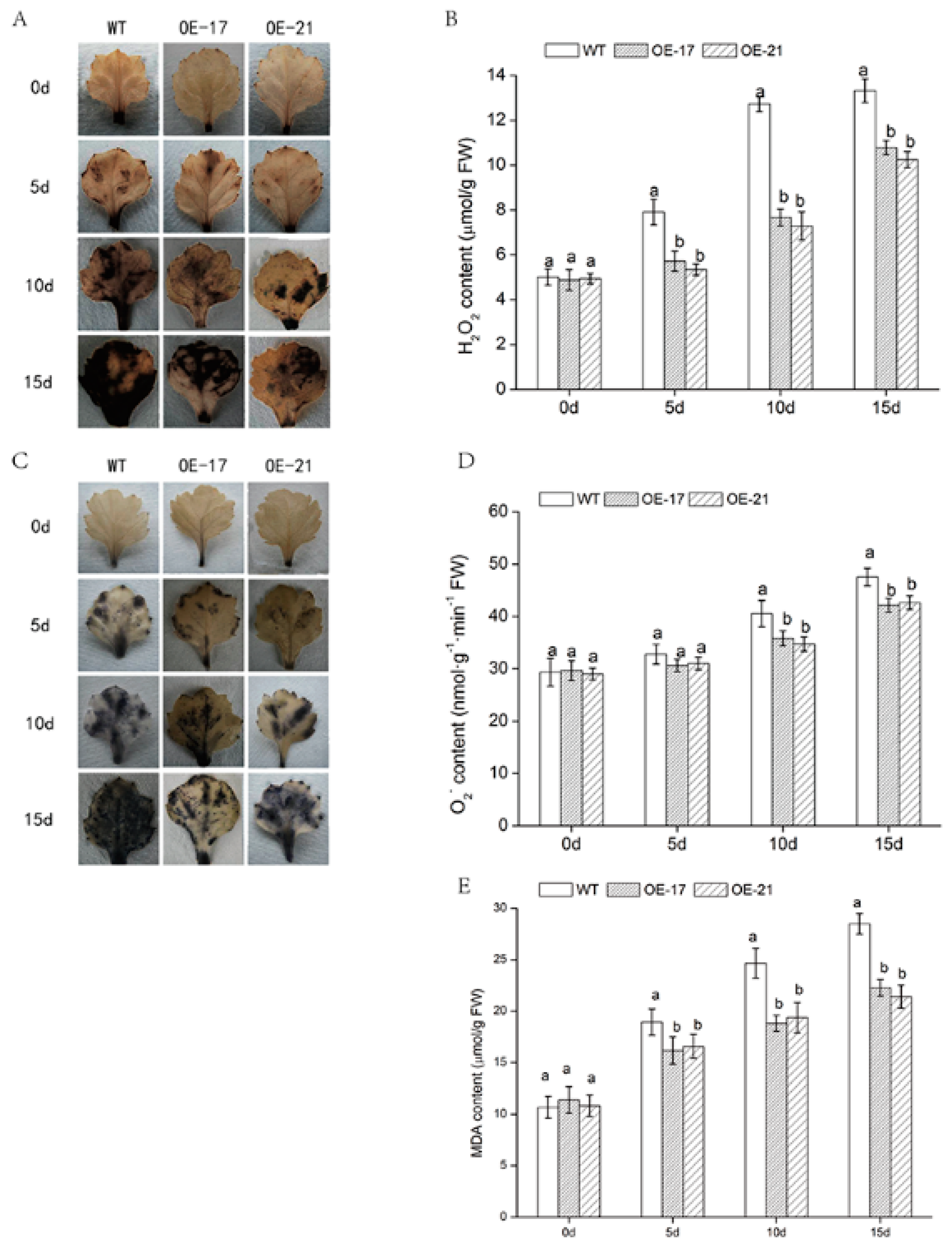
2.4. Accumulation of H2O2, O2−, and MDA in DgWRKY2 Transgenic Chrysanthemum under Salt Stress
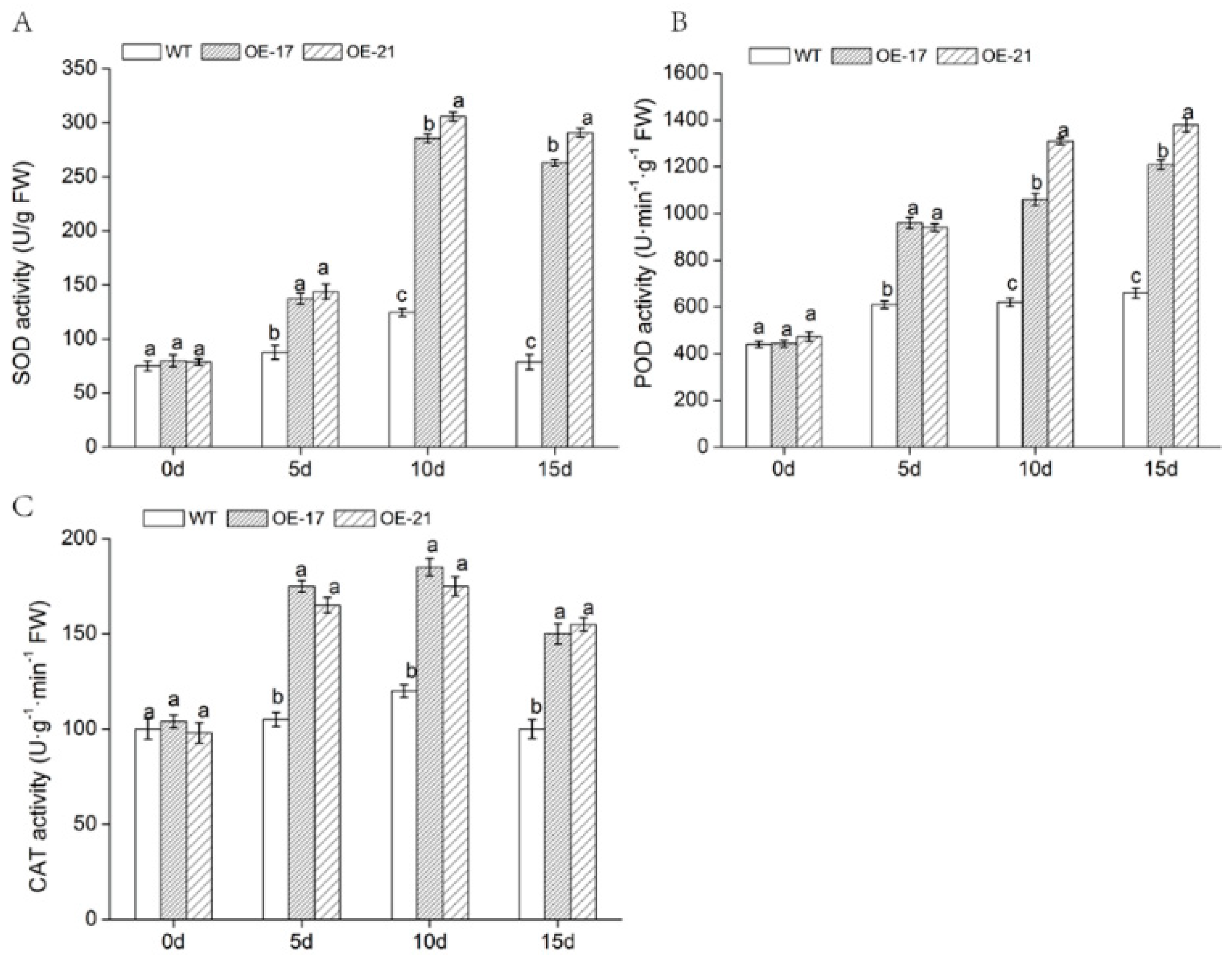
2.5. Physiological Changes in DgWRKY2 Transgenic Chrysanthemum
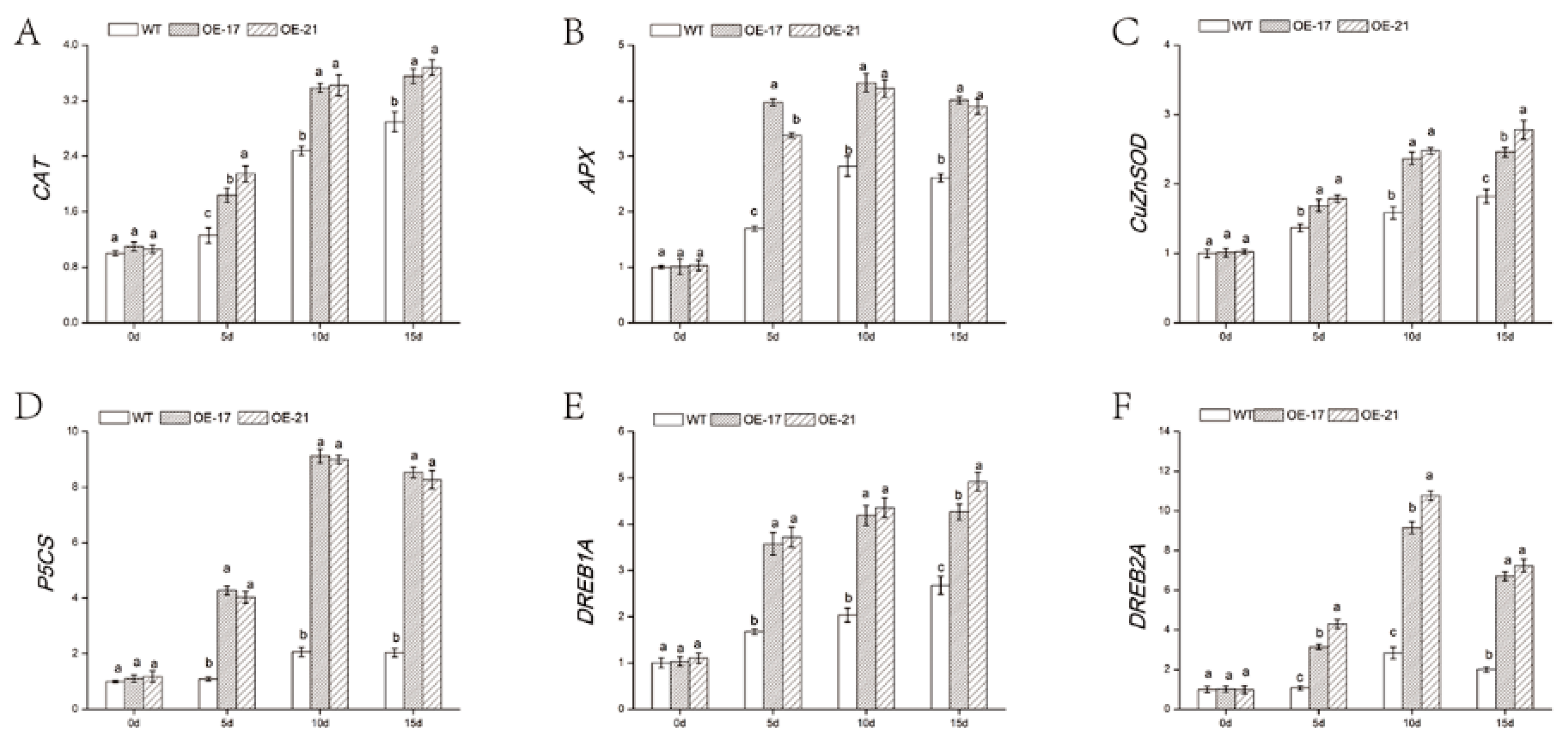
2.6. Expression of Abiotic Stress-Related Genes in DgWRKY2 Transformed Chrysanthemum
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Cloning of DgWRKY2 and Sequence Analysis
4.3. Generation of Transgenic Chrysanthemum
4.4. Expression of DgWRKY2 under Salt Treatment
4.5. Salt Treatment of Transgenic Chrysanthemum and Stress Tolerance Assays
4.6. Determination of Physiological Indexes of Transgenic Chrysanthemum under Salt Stress
4.7. Histochemical Detection of Reactive Oxygen Species (ROS)
4.8. Expression of Salt Stress Response Genes in Dgwrky2 Transgenic Chrysanthemum
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene Expression Profiling of Plants under Salt Stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Bing, L.; Zhao, B.C.; Shen, Y.Z.; Huang, Z.J.; Ge, R.C. Progress of Study on Salt Tolerance and Salt Tolerant Related Genes in Plant. J. Hebei Norm. Univ. 2008, 2, 243–248. [Google Scholar]
- Tuteja, N. Mechanisms of high salinity tolerance in plants. Methods Enzymol. 2007, 428, 419–438. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, H.; Yang, X.; Li, Q.; Ling, J.; Wang, H.; Gu, X.; Huang, S.; Jiang, W. CsWRKY46, a WRKY transcription factor from cucumber, confers cold resistance in transgenic-plant by regulating a set of cold-stress responsive genes in an ABA-dependent manner. Plant Physiol. Biochem. 2016, 108, 478. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, M.; Oelmüller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 9, e27700. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Rabara, R.C.; Rushton, P.J. A systems biology perspective on the role of WRKY transcription factors in drought responses in plants. Planta 2014, 239, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cai, Z.; Gan, S. Transcriptome of Arabidopsis leaf senescence. Plant Cell Environ. 2004, 27, 521–549. [Google Scholar] [CrossRef]
- Qin, Y.; Tian, Y.; Liu, X. A wheat salinity-induced WRKY transcription factor TaWRKY93 confers multiple abiotic stress tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 428. [Google Scholar] [CrossRef] [PubMed]
- Nam, T.N.; Le, H.T.; Mai, D.S.; Tuan, N.V. Overexpression of NbWRKY79, enhances salt stress tolerance in Nicotiana benthamiana. Acta Physiol. Plant. 2017, 39, 121. [Google Scholar] [CrossRef]
- Du, C.; Zhao, P.; Zhang, H.; Li, N.; Zheng, L.; Wang, Y. The Reaumuria trigyna transcription factor RtWRKY1 confers tolerance to salt stress in transgenic Arabidopsis. J. Plant Physiol. 2017, 215, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Hou, L.; Xiao, P.; Guo, Y.; Deyholos, M.K.; Liu, X. VvWRKY30, a grape WRKY transcription factor, plays a positive regulatory role under salinity stress. Plant Sci. 2018. [Google Scholar] [CrossRef]
- Powers, S.K.; Lennon, S.L.; Quindry, J.; Mehta, J.L. Exercise and cardioprotection. Curr. Opin. Cardiol. 2002, 17, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Meloni, D.A.; Oliva, M.A.; Martinez, C.A.; Cambraia, J. Photosynthesis and activity of superoxide dismutase, peroxidase and glutathione reductase in cotton under salt stress. Environ. Exp. Bot. 2003, 49, 69–76. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Negi, N.P.; Sharma, V.; Sarin, N.B. Pyramiding of Two Antioxidant Enzymes CuZnSOD and cAPX from Salt Tolerant Cell Lines of Arachis hypogeae Confers Drought Stress Tolerance in Nicotiana tabacum. Indian J. Agric. Biochem. 2017, 30, 141. [Google Scholar] [CrossRef]
- Hui, Y.; Qiang, L.; Park, S.C.; Wang, X.; Liu, Y.J.; Zhang, Y.G.; Tang, W.; Kou, M.; Ma, D.F. Overexpression of CuZnSOD, and APX, enhance salt stress tolerance in sweet potato. Plant Physiol. Biochem. 2016, 109, 20–27. [Google Scholar] [CrossRef]
- Yang, Z.; Zhou, Y.; Ge, L.; Li, G.; Liu, Q.; Xu, Y.; Jiang, L.; Yang, Y.; School of Agriculture, Jiangxi Agricultural University; School of Sciences, Jiangxi Agricultural University. Expression of Cucumber CsCAT3 Gene under Stress and Its Salt Tolerance in Transgenic Arabidopsis thaliana. Mol. Plant Breed. 2018. [Google Scholar]
- Guerzoni, J.T.S.; Belintani, N.G.; Moreira, R.M.P.; Hoshino, A.A.; Domingues, D.S.; Filho, J.C.B.; Vieira, L.G.E. Stress-induced Δ1-pyrroline-5-carboxylate synthetase (P5CS) gene confers tolerance to salt stress in transgenic sugarcane. Acta Physiol. Plant. 2014, 36, 2309–2319. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Kakimoto, M.; Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J. 2010, 50, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.L.; Ma, J.T.; Zhao, Y.M.; Wei, Y.H.; Tang, Y.X.; Wu, Y.M. Improvement of drought and salt tolerance in Arabidopsis and Lotus corniculatus by overexpression of a novel DREB transcription factor from Populus euphratica. Gene 2012, 506, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.T.; Yin, C.C.; Guo, Q.Q.; Zhou, M.L.; Wang, Z.L.; Wu, Y.M. A novel DREB transcription factor from Halimodendron halodendron, leads to enhance drought and salt tolerance in Arabidopsis. Biol. Plant. 2014, 59, 74–82. [Google Scholar] [CrossRef]
- Akça, Y.; Samsunlu, E. The effect of salt stress on growth, chlorophyll content, proline and nutrient accumulation, and k/na ratio in walnut. Am. Bank. 2012, 1999, 1513–1520. [Google Scholar]
- Wu, Y.H.; Wang, T.; Wang, K.; Liang, Q.Y.; Bai, Z.Y.; Liu, Q.L.; Jiang, B.B.; Zhang, L. Comparative Analysis of the Chrysanthemum Leaf Transcript Profiling in Response to Salt Stress. PLoS ONE 2016, 11, e0159721. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.L.; Xu, K.D.; Pan, Y.Z.; Jiang, B.B.; Liu, G.L.; Jia, Y.; Zhang, H.Q. Functional Analysis of a Novel Chrysanthemum WRKY Transcription Factor Gene Involved in Salt Tolerance. Plant Mol. Biol. Rep. 2014, 32, 282–289. [Google Scholar] [CrossRef]
- Liu, Q.L.; Zhong, M.; Li, S.; Pan, Y.Z.; Jiang, B.B.; Jia, Y.; Zhang, H.Q. Overexpression of a chrysanthemum transcription factor gene, DgWRKY3, intobacco enhances tolerance to salt stress. Plant Physiol. Biochem. 2013, 69, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wu, Y.H.; Tian, X.Q.; Bai, Z.Y.; Liang, Q.Y.; Liu, Q.L.; Pan, Y.Z.; Zhang, L.; Jiang, B.B. Overexpression of DgWRKY4 Enhances Salt Tolerance in Chrysanthemum Seedlings. Front. Plant Sci. 2017, 8, 1592. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.Y.; Wu, Y.H.; Wang, K.; Bai, Z.Y.; Liu, Q.L.; Pan, Y.Z.; Zhang, L.; Jiang, B.B. Chrysanthemum WRKY gene DgWRKY5 enhances tolerance to salt stress in transgenic chrysanthemum. Sci. Rep. 2017, 7, 4799. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H. Selenium Promotes the Growth and Photosynthesis of Tomato Seedlings Under Salt Stress by Enhancing Chloroplast Antioxidant Defense System. J. Plant Growth Regul. 2014, 33, 671–682. [Google Scholar] [CrossRef]
- Ben, K.R.; Abdelly, C.; Savouré, A. Proline, a multifunctional amino-acid involved in plant adaptation to environmental constraints. Biol. Aujourdhui 2012, 206, 291. [Google Scholar] [CrossRef]
- Chaleff, R.S. Further characterization of picloram tolerant mutance of Nicotinana tabacum. Theor. Appl. Genet. 1980, 58, 91–95. [Google Scholar] [CrossRef]
- Wang, F.; Liu, P.; Zhu, J. Effect of magnesium (Mg) on contents of free proline, soluble sugar and protein in soybean leaves. J. Henan Agric. Sci. 2004, 6, 35–38. [Google Scholar]
- Fu, Q.T.; Yu, D.Q. Expression profiles of AtWRKY25, AtWRKY26 and AtWRKY33 under abiotic stresses. Hereditas 2010, 32, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.X. Salt-Tolerant Drought-Tolerant Wheat Gene TaWRKY79 and Application Thereof. CN 102703465A, 3 October 2012. [Google Scholar]
- Tian, Y.C.; Qin, Y.X. Wheat Salt-Tolerant and Drought-Resistant Gene TaWRKY80 and Application Thereof. CN 102703466B, 3 July 2013. [Google Scholar]
- Wang, H.; Hao, J.; Chen, X.; Hao, Z.; Wang, X.; Lou, Y.; Peng, Y.; Guo, Z. Overexpression of rice WRKY89 enhances ultraviolet B tolerance and disease resistance in rice plants. Plant Mol. Biol. 2007, 65, 799–815. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.Y.; Tian, A.G.; Zou, H.F.; Xie, Z.M.; Lei, G.; Huang, J.; Wang, C.M.; Wang, H.W.; Zhang, J.S.; Chen, S.Y. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants. Plant Biotechnol. J. 2008, 6, 486–503. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Hou, X.; Tang, J.; Wang, Z.; Wang, S.; Jiang, F.; Li, Y. A novel cold-inducible gene from Pak-choi (Brassica campestris, ssp. chinensis), BcWRKY46, enhances the cold, salt and dehydration stress tolerance in transgenic tobacco. Mol. Biol. Rep. 2012, 39, 4553. [Google Scholar] [CrossRef]
- Song, Y.; Jing, S.J.; Yu, D.Q. Overexpression of the stress-induced OsWRKY08 improves osmotic stress tolerance in Arabidopsis. Chin. Sci. Bull. 2009, 54, 4671–4678. [Google Scholar] [CrossRef]
- Zhong, G.M.; Wu, L.T.; Wang, J.M.; Yang, Y.; Li, X.F. Subcellular localization and expression analysis of transcription factor AtWRKY28 under biotic stresses. J. Agric. Sci. Technol. 2012, 14, 57–63. [Google Scholar]
- Zhao, Y.; Zhou, Y.; Jiang, H.; Li, X.; Gan, D.; Peng, X.; Zhu, S.; Cheng, B. Systematic Analysis of Sequences and Expression Patterns of Drought-Responsive Members of the HD-Zip Gene Family in Maize. PLoS ONE 2011, 6, e28488. [Google Scholar] [CrossRef] [PubMed]
- Raghavendra, A.S.; Padmasree, K.; Saradadevi, K. Interdependence of photosynthesis and respiration in plant cells: Interactions between chloroplasts and mitochondria. Plant Sci. 1994, 97, 1–14. [Google Scholar] [CrossRef]
- Li, P.; Song, A.; Gao, C.; Wang, L.; Wang, Y.; Sun, J.; Jiang, J.; Chen, F.; Chen, S. Chrysanthemum WRKY gene CmWRKY17, negatively regulates salt stress tolerance in transgenic chrysanthemum and Arabidopsis plants. Plant Cell Rep. 2015, 34, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, L.; Chen, J.; Tao, L.; An, Y.; Cai, H.; Guo, C. Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean. PLoS ONE 2018, 13, e0192382. [Google Scholar] [CrossRef] [PubMed]
- Skórzyńskapolit, E. Lipid peroxidation in plant cells, its physiological role and changes under heavy metal stress. Acta Soc. Bot. Pol. 2007, 76, 49–54. [Google Scholar] [CrossRef]
- Jain, M.; Mathur, G.; Koul, S.; Sarin, N. Ameliorative effects of proline on salt stress-induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant Cell Rep. 2001, 20, 463–468. [Google Scholar] [CrossRef]
- Tang, M.; Liu, X.; Deng, H.; Shen, S. Over-expression of JcDREB, a putative AP2/EREBP domain-containing transcription factor gene in woody biodiesel plant Jatropha curcas, enhances salt and freezing tolerance in transgenic Arabidopsis thaliana. Plant Sci. Int. J. Exp. Plant Biol. 2011, 181, 623. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 2006, 18, 1292–1309. [Google Scholar] [CrossRef] [PubMed]
- Mallikarjuna, G.; Mallikarjuna, K.; Reddy, M.K.; Kaul, T. Expression of OsDREB2A, transcription factor confers enhanced dehydration and salt stress tolerance in rice (Oryza sativa, L.). Biotechnol. Lett. 2011, 33, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- An, G.; Watson, B.D.; Chiang, C.C. Transformation of Tobacco, Tomato, Potato, and Arabidopsis thaliana Using a Binary Ti Vector System. Plant Physiol. 1986, 81, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.P.; Yu, M.; Zhang, A.M. Studies on callus induced from leaves and plantlets regeneration of the traditional Chinese medicine Chrysanthemum morifolium. China J. Chin. Mater. Med. 2003, 28, 213–216. [Google Scholar]
- Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Jiang, J.; Chen, S.; Chen, F.; Guan, Z.; Fang, W. The constitutive expression of Chrysanthemum dichrum ICE1 in Chrysanthemum grandiflorum improves the level of low temperature, salinity and drought tolerance. Plant Cell Rep. 2012, 31, 1747–1758. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiment, 3rd ed.; Higher Education Press: Beijing, China, 2015; pp. 182–184. [Google Scholar]
- Zhang, L.; Tian, L.H.; Zhao, J.F.; Song, Y.; Zhang, C.J.; Guo, Y. Identification of an apoplastic protein involved in the initial phase of salt stress response in rice root by two-dimensional electrophoresis. Plant Physiol. Plant Signal. Behav. 2009, 149, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Wang, S.F.; Chen, Y.T. Effects of salt stress on growth and physiological index of 6 tree species. For. Res. 2009, 22, 315–324. [Google Scholar]
- Jin, Y.; Donglin, L.I.; Ding, Y.; Wang, L. Effects of salt stress on photosynthetic characteristics and chlorophyll content of Sapium sebiferum seedlings. J. Nanjing For. Univ. 2011, 35, 29–33. [Google Scholar]
- Shi, J.; Fu, X.Z.; Peng, T.; Huang, X.S.; Fan, Q.J.; Liu, J.H. Spermine pretreatment confers dehydration tolerance of citrus in vitro plants via modulation of antioxidative capacity and stomatal response. Tree Physiol. 2010, 30, 914–922. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| DgWRKY2 | F: ATTTGTCAAACTTCTCCTCTCTTCT |
| R: GTGGGGGTGGGGGTGGATA | |
| EF1a | F: TTTTGGTATCTGGTCCTGGAG |
| R: CCATTCAAGCGACAGACTCA | |
| Cu/Zn SOD | F: CCATTGTTGACAAGCAGATTCCACTCA |
| R: ATCATCAGGATCAGCATGGACGACTAC | |
| CAT | F: TACAAGCAACGCCCTTCAA |
| R: GACCTCTGTTCCCAACAGTCA | |
| APX | F: GTTGGCTGGTGTTGTTGCT |
| R: GATGGTCGTTTCCCTTAGTTG | |
| P5CS | F: TTGGAGCAGAGGTTGGAAT |
| R: GCAGGTCTTTGTGGGTGTAG | |
| DREB1A | F: CGGTTTTGGCTATGAGGGGT |
| R: TTCTTCTGCCAGCGTCACAT | |
| DREB2A | F: GATCGTGGCTGAGAGACTCG |
| R: TACCCCACGTTCTTTGCCTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Wu, Y.-H.; Zhao, Q.; Wang, B.; Liu, Q.-L.; Zhang, L. Chrysanthemum DgWRKY2 Gene Enhances Tolerance to Salt Stress in Transgenic Chrysanthemum. Int. J. Mol. Sci. 2018, 19, 2062. https://doi.org/10.3390/ijms19072062
He L, Wu Y-H, Zhao Q, Wang B, Liu Q-L, Zhang L. Chrysanthemum DgWRKY2 Gene Enhances Tolerance to Salt Stress in Transgenic Chrysanthemum. International Journal of Molecular Sciences. 2018; 19(7):2062. https://doi.org/10.3390/ijms19072062
Chicago/Turabian StyleHe, Ling, Yin-Huan Wu, Qian Zhao, Bei Wang, Qing-Lin Liu, and Lei Zhang. 2018. "Chrysanthemum DgWRKY2 Gene Enhances Tolerance to Salt Stress in Transgenic Chrysanthemum" International Journal of Molecular Sciences 19, no. 7: 2062. https://doi.org/10.3390/ijms19072062
APA StyleHe, L., Wu, Y.-H., Zhao, Q., Wang, B., Liu, Q.-L., & Zhang, L. (2018). Chrysanthemum DgWRKY2 Gene Enhances Tolerance to Salt Stress in Transgenic Chrysanthemum. International Journal of Molecular Sciences, 19(7), 2062. https://doi.org/10.3390/ijms19072062