Mechanisms of Tumor Growth Inhibition by Depletion of γ-Glutamylcyclotransferase (GGCT): A Novel Molecular Target for Anticancer Therapy

 , , ,
, , , {kind=link}
{kind=link}
Abstract
1. Introduction
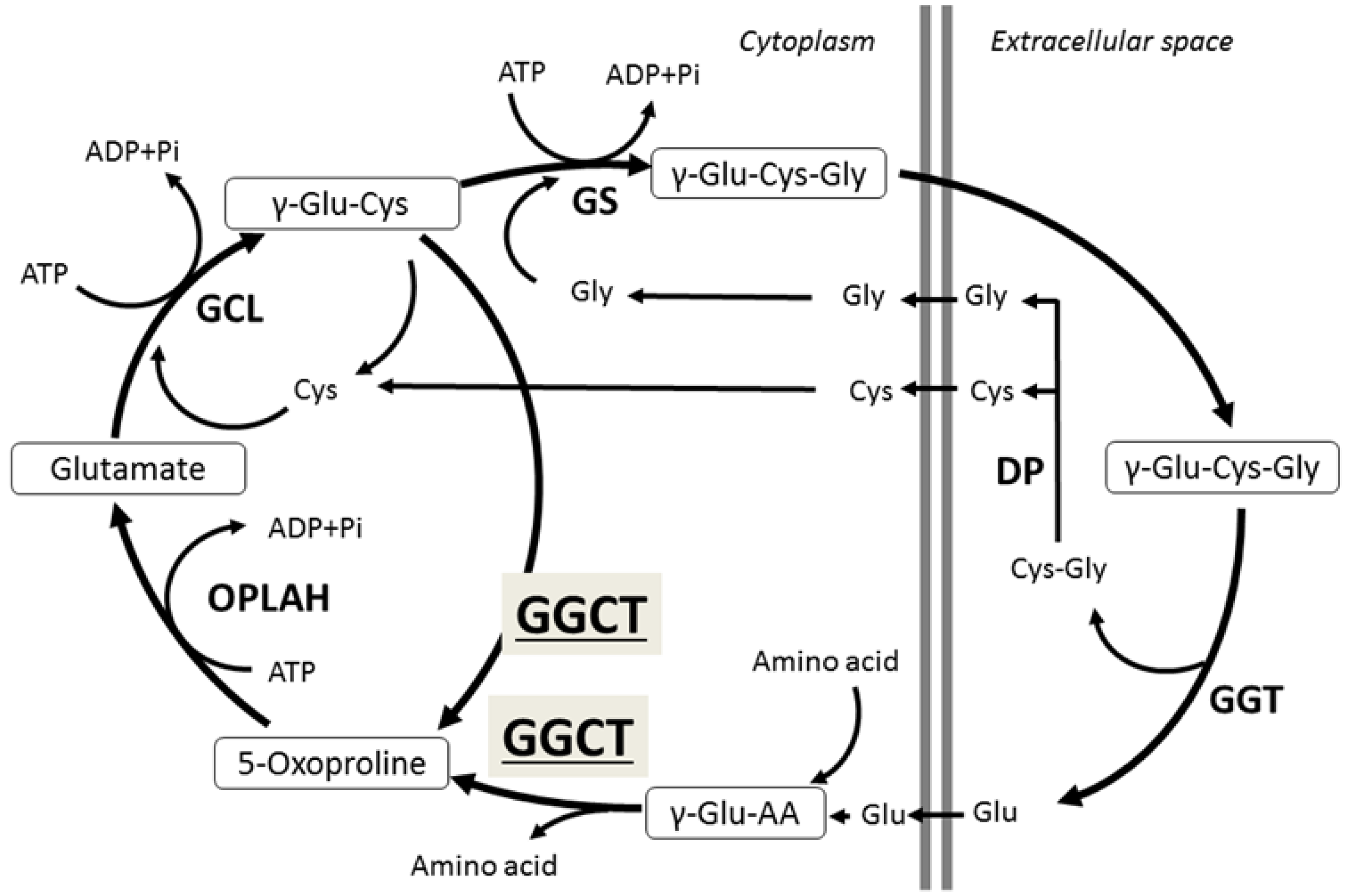
2. γ-Glutamylcyclotransferase (GGCT)
3. GGCT Upregulation in Cancer
4. Molecular Regulation Mechanism of GGCT Expression in Cancer Cells
5. Inhibition of Cancer Cell Proliferation and Induction of Cell Death by GGCT Knockdown
6. Intracellular Changes Caused by GGCT Depletion
6.1. Morphological Changes
6.2. Cellular Senescence
6.3. Autophagy
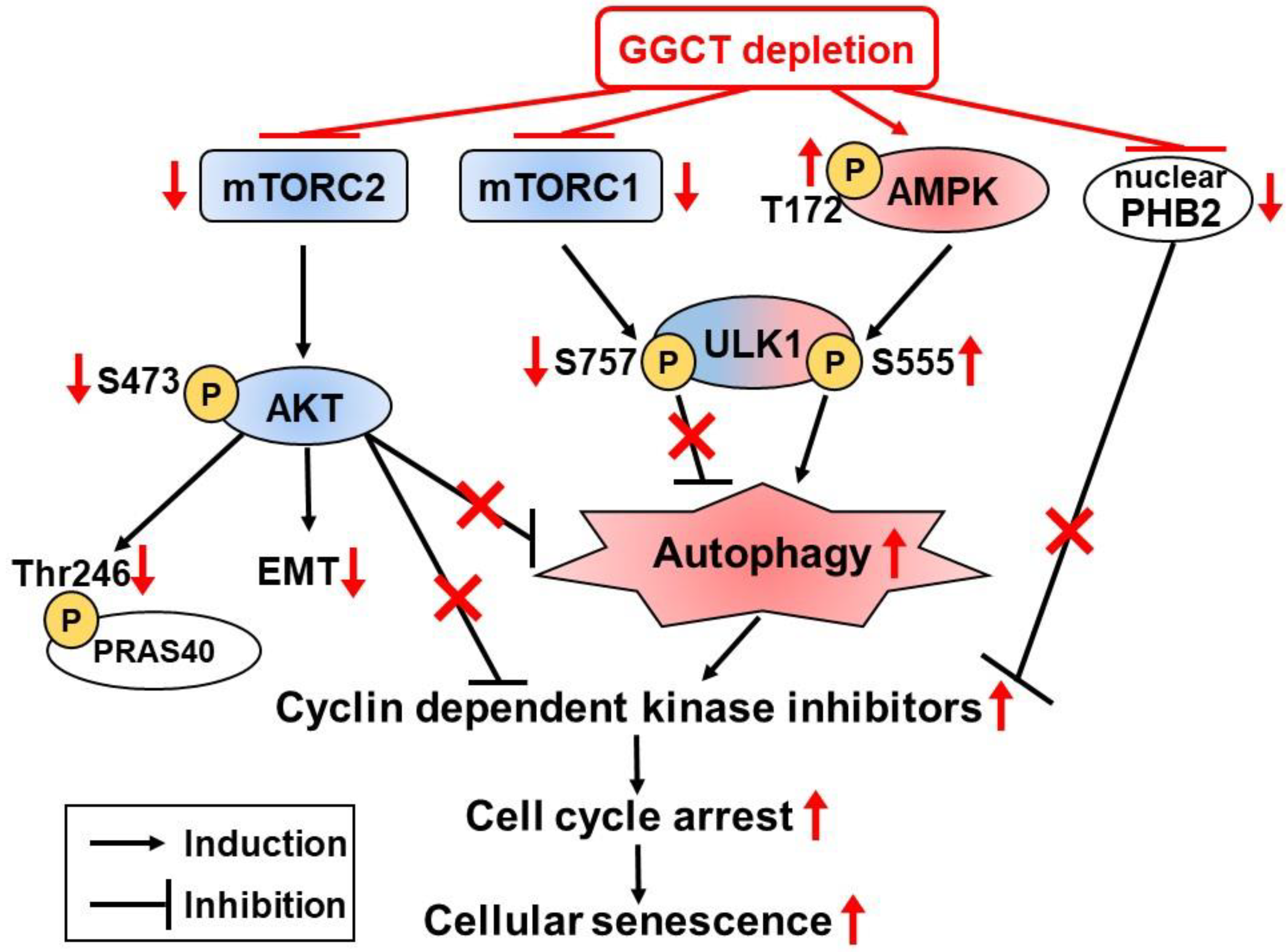
6.4. Changes of Intracellular Signaling Pathways
6.5. Epithelial Mesenchymal Transition (EMT)
6.6. GGCT Interacting Proteins
7. GGCT Fluorogenic Probes, LISA-4 and LISA-101
8. Development of GGCT Inhibitors
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Atg5 | Autophagy related 5 |
| C7orf24 | Chromosome 7 open reading frame 24 |
| CDK | Cyclin dependent kinase |
| EMT | Epithelial mesenchymal transition |
| GA | N-glutaryl-l-alanine |
| GGCT | γ-glutamylcyclotransferase |
| LC3-II | Microtubule-associated protein light chain 3-II |
| PHB2 | Prohibitin 2 |
| Pro-GA | Prodrug of N-glutaryl-l-alanine |
| SA-β-gal | Senescence associated β-galactosidase |
References
- Liu, Y.; Hyde, A.S.; Simpson, M.A.; Barycki, J.J. Emerging regulatory paradigms in glutathione metabolism. Adv. Cancer Res. 2014, 122, 69–101. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Hanada, E.; Ii, H.; Tomita, K.; Yoshiki, T.; Kawauchi, A. Gamma-Glutamylcyclotransferase: A Novel Target Molecule for Cancer Diagnosis and Treatment. Biomed Res. Int. 2015, 2015, 45219. [Google Scholar] [CrossRef] [PubMed]
- Orlowski, M.; Meister, A. The γ-glutamyl cycle: A possible transport system for amino acids. Proc. Natl. Acad. Sci. USA 1970, 67, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Meister, A. The γ-glutamyl cycle: Diseases associated with specific enzyme deficiencies. Ann. Intern. Med. 1974, 81, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Oakley, A.J.; Yamada, T.; Liu, D.; Coggan, M.; Clark, A.G.; Board, P.G. The identification and structural characterization of C7orf24 as γ-glutamyl cyclotransferase. An essential enzyme in the gamma-glutamyl cycle. J. Biol. Chem. 2008, 283, 22031–22042. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Makino, S.; Masuda, C.; Yoshiki, T.; Kitamura, Y.; Takata, K.; Yanagisawa, D.; Taniguchi, T.; Tooyama, I. The mRNA distribution of C7orf24, a γ-glutamyl cyclotransferase, in rat tissues. J. Histochem. Cytochem. 2009, 57, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Gromov, P.; Gromova, I.; Friis, E.; Timmermans-Wielenga, V.; Rank, F.; Simon, R.; Sauter, G.; Moreira, J.M. Proteomic profiling of mammary carcinomas identifies C7orf24, a γ-glutamyl cyclotransferase, as a potential cancer biomarker. J. Proteome Res. 2010, 9, 3941–3953. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Eishi, Y.; Yamada, T.; Uchida, K.; Minegishi, K.; Tamura, T.; Kobayashi, D.; Hiroshi, K.; Suzuki, T.; Board, P.G. Widespread expression of γ-glutamyl cyclotransferase suggests it is not a general tumor marker. J. Histochem. Cytochem. 2012, 60, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Maeda, S.; Watanabe, A.; Sano, Y.; Aiuchi, T.; Nakajo, S.; Itabe, H.; Nakaya, K. A novel 21-kDa cytochrome c-releasing factor is generated upon treatment of human leukemia U937 cells with geranylgeraniol. Biochem. Biophys. Res. Commun. 2006, 346, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Geman, D.; Winslow, R.L. Large-scale integration of cancer microarray data identifies a robust common cancer signature. BMC Bioinform. 2007, 8, 275. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Isono, T.; Iwaki, H.; Wakabayashi, Y.; Okada, Y.; Kontani, K.; Yoshimura, K.; Terai, A.; Arai, Y.; Yoshiki, T. Identification by proteomic analysis of calreticulin as a marker for bladder cancer and evaluation of the diagnostic accuracy of its detection in urine. Clin. Chem. 2004, 50, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Iwaki, H.; Inoue, H.; Isono, T.; Yuasa, T.; Nogawa, M.; Maekawa, T.; Ueda, M.; Kajita, Y.; Ogawa, O.; et al. A novel tumor-related protein, C7orf24, identified by proteome differential display of bladder urothelial carcinoma. Proteom. Clin. Appl. 2007, 1, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Uejima, D.; Nishijo, K.; Kajita, Y.; Ishibe, T.; Aoyama, T.; Kageyama, S.; Iwaki, H.; Nakamura, T.; Iida, H.; Yoshiki, T.; et al. Involvement of cancer biomarker C7orf24 in the growth of human osteosarcoma. Anticancer Res. 2011, 31, 1297–1305. [Google Scholar] [PubMed]
- Takemura, K.; Kawachi, H.; Eishi, Y.; Kitagaki, K.; Negi, M.; Kobayashi, M.; Uchida, K.; Inoue, J.; Inazawa, J.; Kawano, T.; et al. γ-Glutamylcyclotransferase as a novel immunohistochemical biomarker for the malignancy of esophageal squamous tumors. Hum. Pathol. 2014, 45, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.H.; Yu, N.; Liu, X.Y.; Tan, G.W.; Wang, Z.X. γ-glutamylcyclotransferase promotes the growth of human glioma cells by activating Notch-Akt signaling. Biochem. Biophys. Res. Commun. 2016, 471, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, T.; Wang, Y.; Yang, L.; Hu, C.; Chen, L.; Wu, S. γ-Glutamyl cyclotransferase contributes to tumor progression in high grade serous ovarian cancer by regulating epithelial-mesenchymal transition via activating PI3K/AKT/mTOR pathway. Gynecol. Oncol. 2018, 149, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Azumi, K.; Ikeda, Y.; Takeuchi, T.; Nomura, T.; Sabau, S.V.; Hamada, J.; Okada, F.; Hosokawa, M.; Yokosawa, H. Localization and characterization of γ-glutamyl cyclotransferase in cancer cells. Mol. Med. Rep. 2009, 2, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Hattori, A.; Ueda, M.; Kageyama, S.; Yoshiki, T.; Kakeya, H. Multiple NF-Y-binding CCAAT boxes are essential for transcriptional regulation of the human C7orf24 gene, a novel tumor-associated gene. FEBS J. 2011, 278, 4088–4099. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Xiong, L.; Zhou, J.; Wang, J.; Li, Z.; Hu, H.; Lin, Q. γ-Glutamylcyclotransferase knockdown inhibits growth of lung cancer cells through G0/G1 phase arrest. Cancer Biother. Radiopharm. 2015, 30, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, L.; Xiang, H.; Hu, C.; Shi, W.; Dong, P.; Lv, W. Knockdown of GGCT inhibits cell proliferation and induces late apoptosis in human gastric cancer. BMC Biochem. 2016, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhou, Y.; Liao, Z.; Huang, Q.; Feng, S.; Li, Y. Role of γ-glutamyl cyclotransferase as a therapeutic target for colorectal cancer based on the lentivirus-mediated system. Anticancer Drugs 2016, 27, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, K.; Nakata, S.; Taniguchi, K.; Ii, H.; Ashihara, E.; Kageyama, S.; Kawauchi, A.; Yoshiki, T. Depletion of γ-glutamylcyclotransferase inhibits breast cancer cell growth via cellular senescence induction mediated by CDK inhibitor upregulation. BMC Cancer 2016, 16, 748. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Hattori, A.; Yoshiki, T.; Kakeya, H. Association of epigenetic alterations in the human C7orf24 gene with the aberrant gene expression in malignant cells. J. Biochem. 2013, 154, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.K.; Dubick, M.A.; Omaye, S.T. γ-Glutamylcysteine inhibits oxidative stress in human endothelial cells. Life Sci. 2012, 90, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Hama, S.; Arata, M.; Nakamura, I.; Kasetani, T.; Itakura, S.; Tsuchiya, H.; Yoshiki, T.; Kogure, K. Prevention of tumor growth by needle-free jet injection of anti-C7orf24 siRNA. Cancer Gene Ther. 2012, 19, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Ran, R.; Liu, Y.; Gao, H.; Kuang, Q.; Zhang, Q.; Tang, J.; Fu, H.; Zhang, Z.; He, Q. PEGylated hyaluronic acid-modified liposomal delivery system with anti-γ-glutamylcyclotransferase siRNA for drug-resistant MCF-7 breast cancer therapy. J. Pharm. Sci. 2015, 104, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, C.; Hu, K. MiR-194 suppresses human gastric cancer cell proliferation and induces apoptosis by targeting GGCT. Int. J. Clin. Exp. Pathol. 2017, 10, 2589–2598. [Google Scholar]
- Taniguchi, K.; Matsumura, K.; Ii, H.; Kageyama, S.; Ashihara, E.; Chano, T.; Kawauchi, A.; Yoshiki, T.; Nakata, S. Depletion of gamma-glutamylcyclotransferase in cancer cells induces autophagy followed by cellular senescence. Am. J. Cancer Res. 2018, 8, 650–661. [Google Scholar] [PubMed]
- Taniguchi, K.; Matsumura, K.; Kageyama, S.; Ii, H.; Ashihara, E.; Chano, T.; Kawauchi, A.; Yoshiki, T.; Nakata, S. Prohibitin-2 is a novel regulator of p21(WAF1/CIP1) induced by depletion of γ-glutamylcyclotransferase. Biochem. Biophys. Res. Commun. 2018, 496, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Yoshiya, T.; Tsuda, S.; Mochizuki, M.; Hidaka, K.; Tsuda, Y.; Kiso, Y.; Kageyama, S.; Ii, H.; Yoshiki, T.; Nishiuchi, Y. A fluorogenic probe for γ-glutamyl cyclotransferase: Application of an enzyme-triggered O-to-N acyl migration-type reaction. Chembiochem 2013, 14, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Yoshiya, T.; Ii, H.; Tsuda, S.; Kageyama, S.; Yoshiki, T.; Nishiuchi, Y. A GGCT fluorogenic probe: Design, synthesis and application to cancer-related cells. OrgBiomol. Chem. 2015, 13, 3182–3185. [Google Scholar] [CrossRef] [PubMed]
- Ii, H.; Yoshiya, T.; Nakata, S.; Taniguchi, K.; Hidaka, K.; Tsuda, S.; Mochizuki, M.; Nishiuchi, Y.; Tsuda, Y.; Ito, K.; et al. A novel prodrug of a γ-glutamylcyclotransferase inhibitor suppresses cancer cell proliferation in vitro and inhibits tumor growth in a xenograft mouse model of prostate cancer. ChemMedChem 2018, 13, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Yoshiya, T.; Ii, H.; Tsuda, S.; Mochizuki, M.; Kageyama, S.; Yoshiki, T. Design of fluorogenic probes and fluorescent-tagged inhibitors for γ-glutamyl cyclotransferase. J. Pept. Sci. 2017, 23, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Ii, H.; Yoshiki, T.; Hoshiya, N.; Uenishi, J. Synthesis and GGCT Inhibitory Activity of N-Glutaryl-L-alanine Analogues. Chem. Pharm. Bull. 2016, 64, 785–792. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kageyama, S.; Ii, H.; Taniguchi, K.; Kubota, S.; Yoshida, T.; Isono, T.; Chano, T.; Yoshiya, T.; Ito, K.; Yoshiki, T.; et al. Mechanisms of Tumor Growth Inhibition by Depletion of γ-Glutamylcyclotransferase (GGCT): A Novel Molecular Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 2054. https://doi.org/10.3390/ijms19072054
Kageyama S, Ii H, Taniguchi K, Kubota S, Yoshida T, Isono T, Chano T, Yoshiya T, Ito K, Yoshiki T, et al. Mechanisms of Tumor Growth Inhibition by Depletion of γ-Glutamylcyclotransferase (GGCT): A Novel Molecular Target for Anticancer Therapy. International Journal of Molecular Sciences. 2018; 19(7):2054. https://doi.org/10.3390/ijms19072054
Chicago/Turabian StyleKageyama, Susumu, Hiromi Ii, Keiko Taniguchi, Shigehisa Kubota, Tetsuya Yoshida, Takahiro Isono, Tokuhiro Chano, Taku Yoshiya, Kosei Ito, Tatsuhiro Yoshiki, and et al. 2018. "Mechanisms of Tumor Growth Inhibition by Depletion of γ-Glutamylcyclotransferase (GGCT): A Novel Molecular Target for Anticancer Therapy" International Journal of Molecular Sciences 19, no. 7: 2054. https://doi.org/10.3390/ijms19072054
APA StyleKageyama, S., Ii, H., Taniguchi, K., Kubota, S., Yoshida, T., Isono, T., Chano, T., Yoshiya, T., Ito, K., Yoshiki, T., Kawauchi, A., & Nakata, S. (2018). Mechanisms of Tumor Growth Inhibition by Depletion of γ-Glutamylcyclotransferase (GGCT): A Novel Molecular Target for Anticancer Therapy. International Journal of Molecular Sciences, 19(7), 2054. https://doi.org/10.3390/ijms19072054