Preanalytical Biases in the Measurement of Human Blood Sphingolipids

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Characteristics of Study Participants
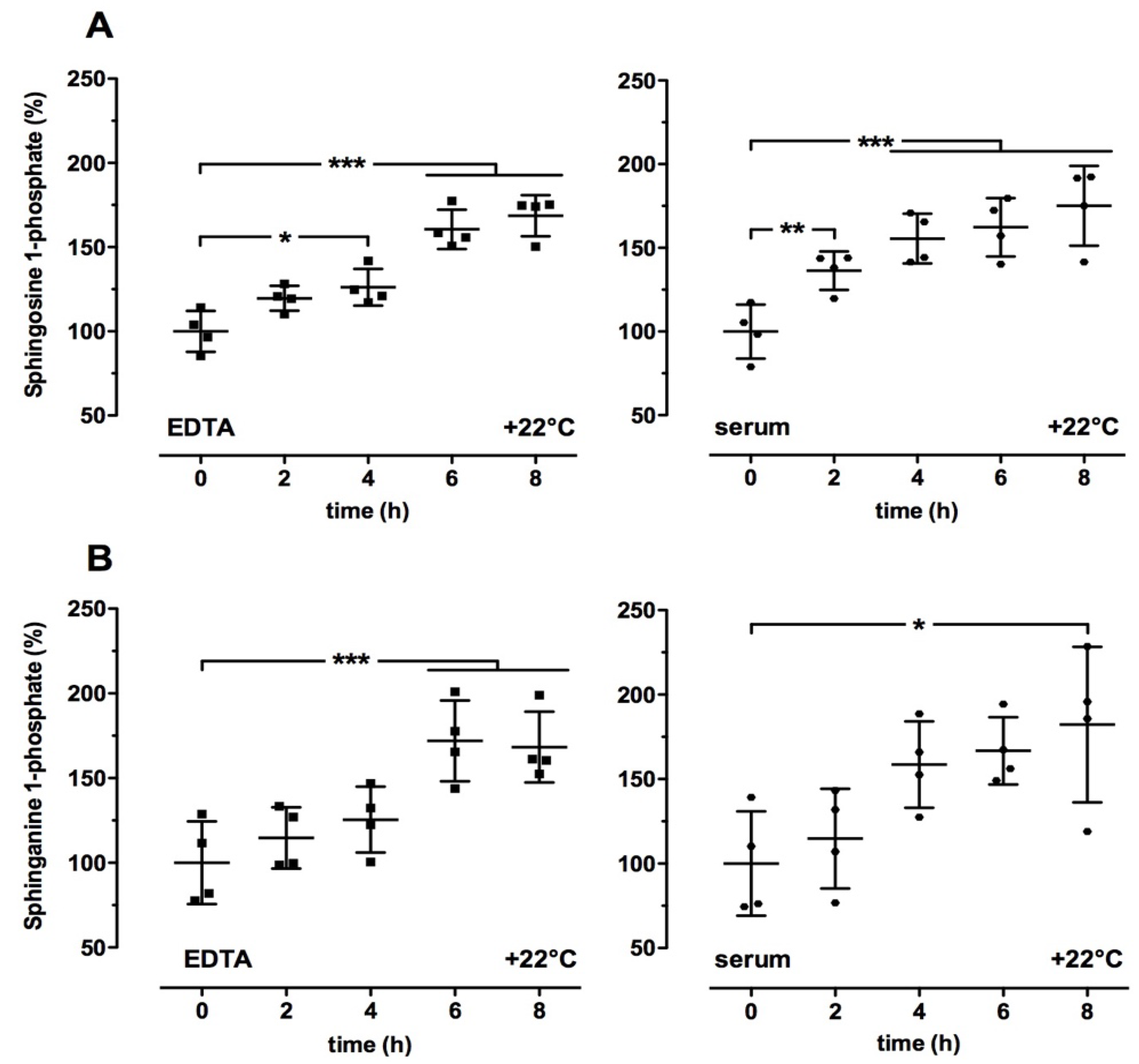
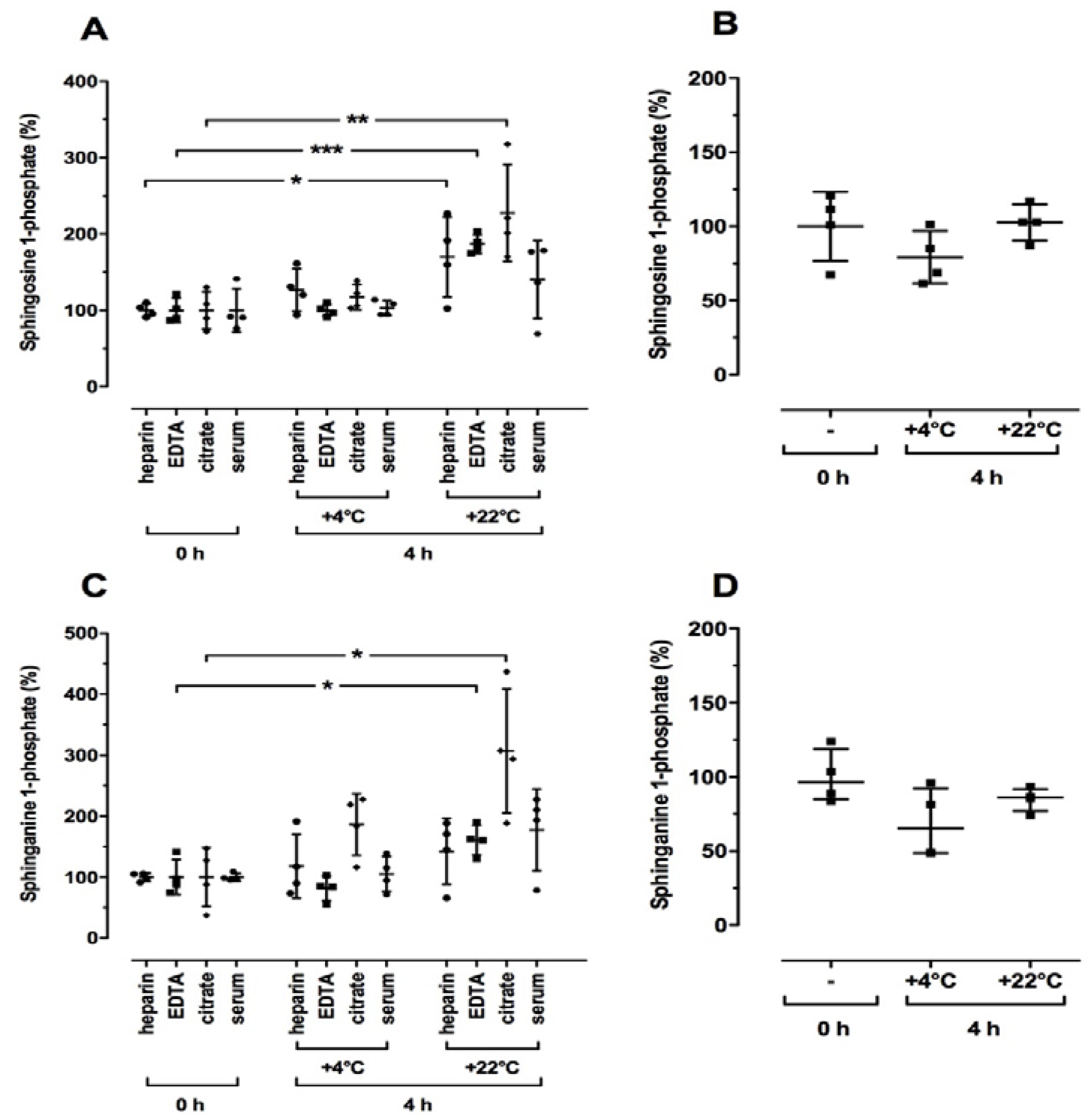
2.2. Time- and Temperature-Dependent Effects on Sphingolipid Levels in Human Blood Samples
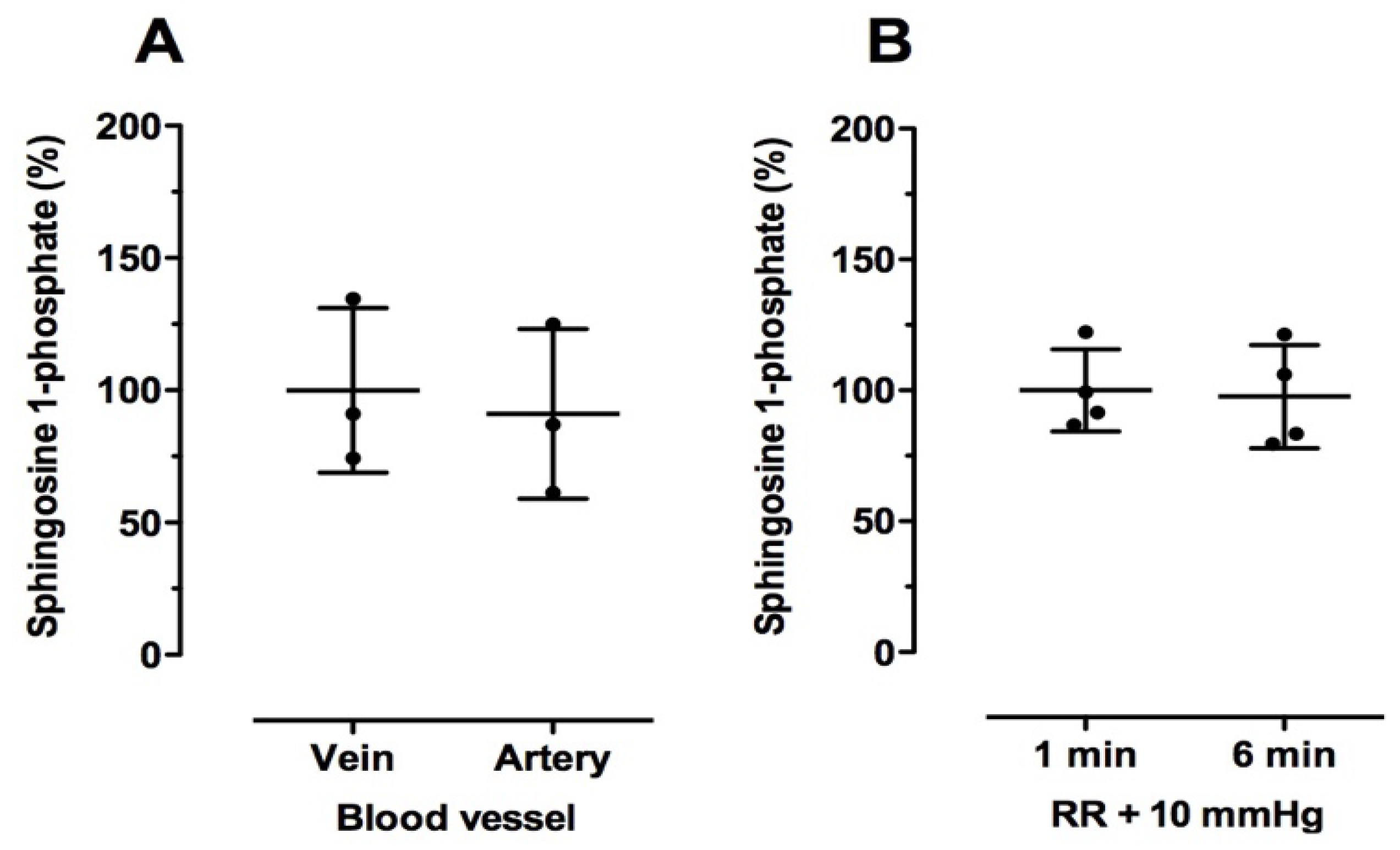
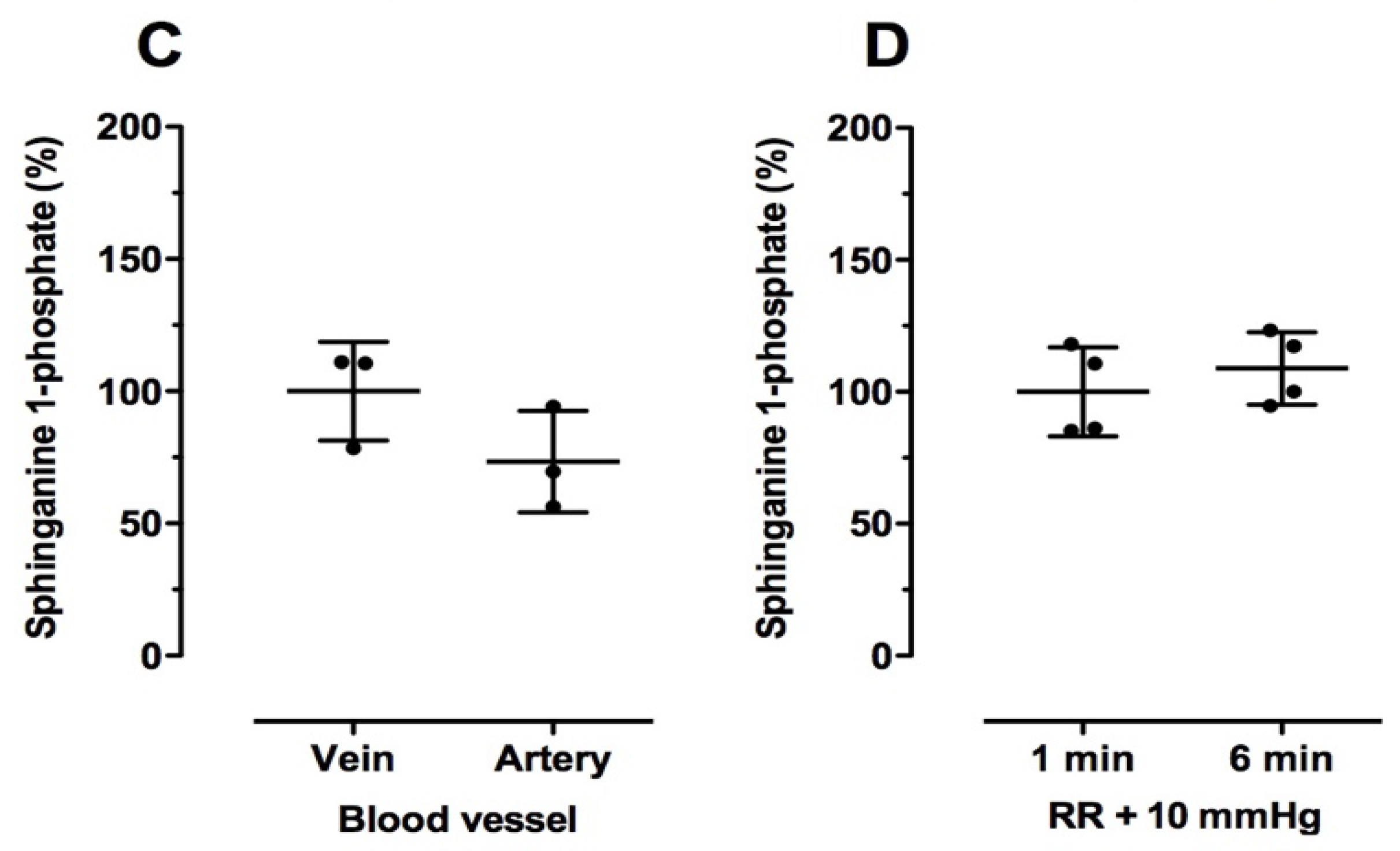
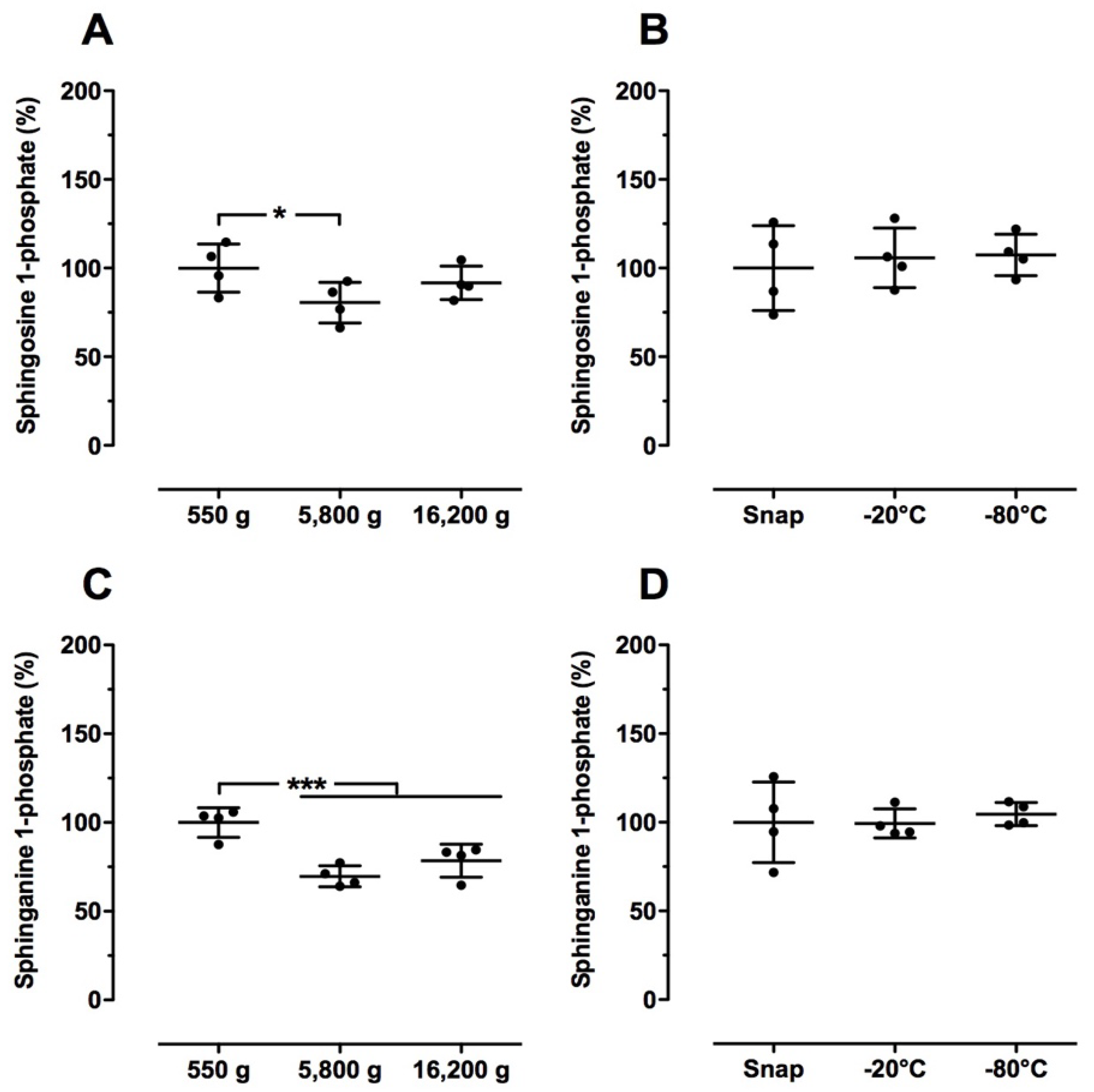
2.3. Effect of Blood Drawing Conditions on Sphingolipid Levels in Human Blood Samples
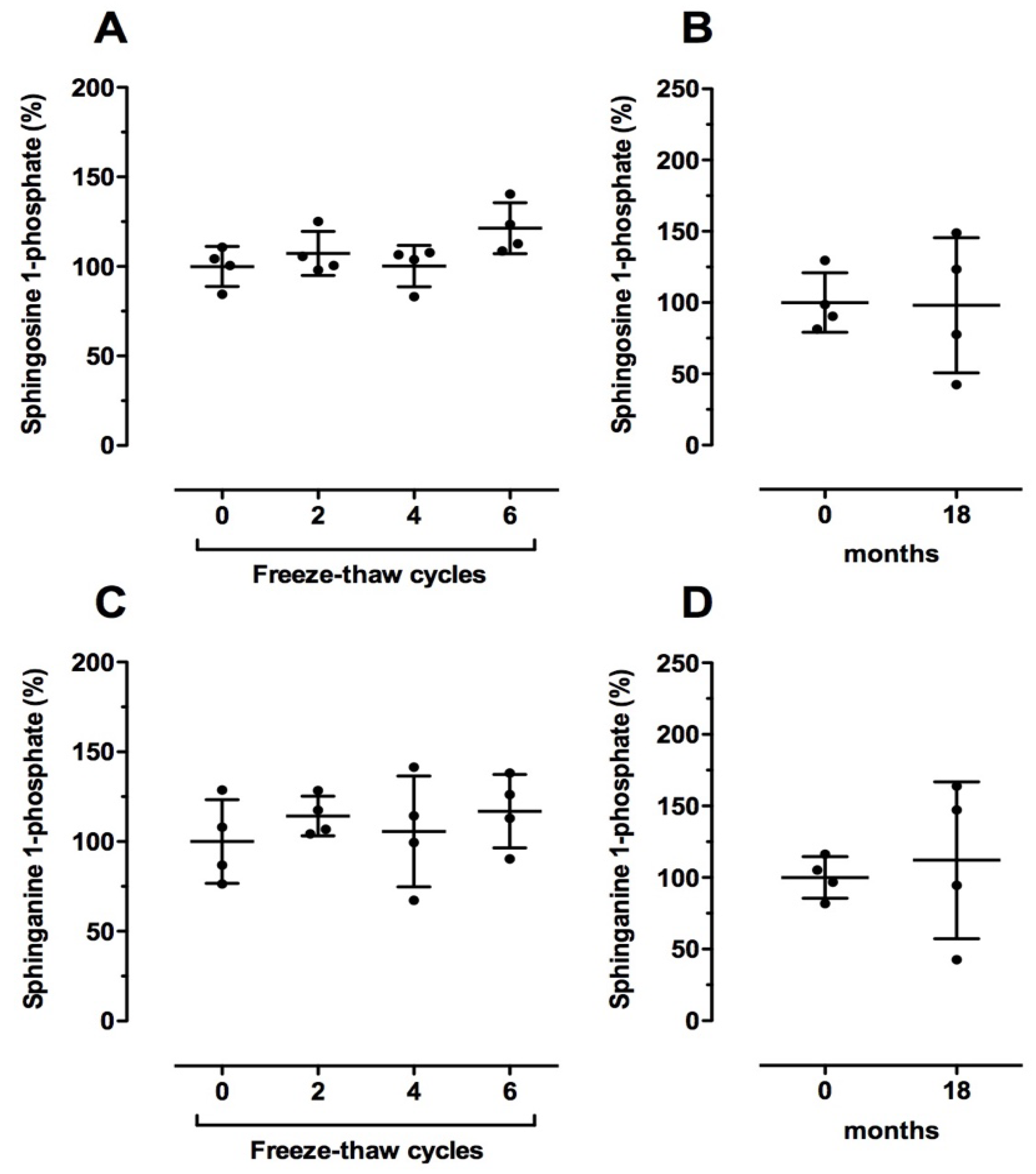
2.4. Effect of Sample Preparation on Sphingolipid Levels in Human Blood Samples
2.5. Effect of Long-Term Storage Conditions on Sphingolipid Levels in Human Blood Samples
3. Discussion
4. Materials and Methods
4.1. Blood Sampling
4.2. Experimental Setup
4.3. LC-MS/MS Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| Cer | ceramide |
| dhCer | dihydroceramide |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| SA1P | sphinganine 1-phosphate |
| S1P | sphingosine 1-phosphate |
| SPHK | sphingosine kinase |
References
- Hla, T.; Dannenberg, A.J. Sphingolipid signaling in metabolic disorders. Cell Metab. 2012, 16, 420–434. [Google Scholar] [CrossRef] [PubMed]
- Soltau, I.; Mudersbach, E.; Geissen, M.; Schwedhelm, E.; Winkler, M.S.; Geffken, M.; Peine, S.; Schoen, G.; Debus, E.S.; Larena-Avellaneda, A.; et al. Serum-Sphingosine-1-Phosphate Concentrations Are Inversely Associated with Atherosclerotic Diseases in Humans. PLoS ONE 2016, 11, e0168302. [Google Scholar] [CrossRef] [PubMed]
- Sattler, K.J.; Elbasan, S.; Keul, P.; Elter-Schulz, M.; Bode, C.; Gräler, M.H.; Bröcker-Preuss, M.; Budde, T.; Erbel, R.; Heusch, G.; Levkau, B. Sphingosine 1-phosphate levels in plasma and HDL are altered in coronary artery disease. Basic Res. Cardiol. 2010, 105, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Mehling, M.; Kappos, L.; Derfuss, T. Fingolimod for multiple sclerosis: mechanism of action, clinical outcomes, and future direction. Curr. Neurol. Neurosci. Rep. 2011, 11, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Ottenlinger, F.M.; Mayer, C.A.; Ferreirós, N.; Schreiber, Y.; Schwiebs, A.; Schmidt, K.G.; Ackermann, H.; Pfeilschifter, J.M.; Radeke, H.H. Interferon-Beta Increases Plasma Ceramides of Specific Chain Length in Multiple Sclerosis Patients, Unlike Fingolimod or Natalizumab. Front. Pharmacol. 2016, 7, 412. [Google Scholar] [CrossRef] [PubMed]
- Barthelmes, J.; de Bazo, A.M.; Pewzner-Jung, Y.; Schmitz, K.; Mayer, C.A.; Foerch, C.; Eberle, M.; Tafferner, N.; Ferreirós, N.; Henke, M.; et al. Lack of ceramide synthase 2 suppresses the development of experimental autoimmune encephalomyelitis by impairing the migratory capacity of neutrophils. Brain Behav. Immun. 2015, 46, 280–292. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Schoell, N.; Ferreirós, N.; Bon, D.; Herrmann, E.; Farnik, H.; Köberle, V.; Piiper, A.; Zeuzem, S.; Kronenberger, B.; et al. Serum sphingolipidomic analyses reveal an upregulation of C16-ceramide and sphingosine-1-phosphate in hepatocellular carcinoma. Oncotarget 2016, 7, 18095–18105. [Google Scholar] [CrossRef] [PubMed]
- Grammatikos, G.; Ferreiros, N.; Bon, D.; Schwalm, S.; Dietz, J.; Berkowski, C.; Fitting, D.; Herrmann, E.; Zeuzem, S.; Sarrazin, C.; et al. Variations in serum sphingolipid levels associate with liver fibrosis progression and poor treatment outcome in hepatitis C virus but not hepatitis B virus infection. Hepatology 2015, 61, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Winkler, M.S.; Nierhaus, A.; Holzmann, M.; Mudersbach, E.; Bauer, A.; Robbe, L.; Zahrte, C.; Geffken, M.; Peine, S.; Schwedhelm, E.; et al. Decreased serum concentrations of sphingosine-1-phosphate in sepsis. Crit. Care 2015, 19, 372. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.; Wegner, D.; Groß, S.; Bahls, M.; Dörr, M.; Felix, S.B.; Ittermann, T.; Oswald, S.; Nauck, M.; Friedrich, N.; et al. Reference intervals for serum sphingosine-1-phosphate in the population-based Study of Health in Pomerania. Clin. Chim. Acta 2017, 468, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.; Fernando, S.; Fernando, R.H.; Wickramasinghe, N.; Shyamali, N.L.; Ogg, G.S.; Malavige, G.N. Sphingosine 1-phosphate in acute dengue infection. PLoS ONE 2014, 9, e113394. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Lehmann, R.; Xu, G. Effects of pre-analytical processes on blood samples used in metabolomics studies. Anal. Bioanal. Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef] [PubMed]
- Scherer, M.; Schmitz, G.; Liebisch, G. High-throughput analysis of sphingosine 1-phosphate, sphinganine 1-phosphate, and lysophosphatidic acid in plasma samples by liquid chromatography-tandem mass spectrometry. Clin. Chem. 2009, 55, 1218–1222. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Peter, A.; Franken, H.; Zhao, X.; Neukamm, S.S.; Rosenbaum, L.; Lucio, M.; Zell, A.; Häring, H.U.; Xu, G.; Lehmann, R. Preanalytical aspects and sample quality assessment in metabolomics studies of human blood. Clin. Chem. 2013, 59, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Hammad, S.M.; Pierce, J.S.; Soodavar, F.; Smith, K.J.; Al Gadban, M.M.; Rembiesa, B.; Klein, R.L.; Hannun, Y.A.; Bielawski, J.; Bielawska, A. Blood sphingolipidomics in healthy humans: impact of sample collection methodology. J. Lipid Res. 2010, 51, 3074–3087. [Google Scholar] [CrossRef] [PubMed]
- Książek, M.; Chacińska, M.; Chabowski, A.; Baranowski, M. Sources, metabolism, and regulation of circulating sphingosine-1-phosphate. J. Lipid Res. 2015, 56, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Thuy, A.V.; Reimann, C.M.; Hemdan, N.Y.; Gräler, M.H. Sphingosine 1-phosphate in blood: Function, metabolism, and fate. Cell. Physiol. Biochem. 2014, 34, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Yang, P.; Proia, R.L.; Hla, T. Erythrocyte-derived sphingosine 1-phosphate is essential for vascular development. J. Clin. Investig. 2014, 124, 4823–4828. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhang, Y.; D’Alessandro, A.; Nemkov, T.; Song, A.; Wu, H.; Liu, H.; Adebiyi, M.; Huang, A.; Wen, Y.E.; et al. Sphingosine-1-phosphate promotes erythrocyte glycolysis and oxygen release for adaptation to high-altitude hypoxia. Nat. Commun. 2016, 7, 12086. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.; Hönig, S.; Liebisch, G.; Lüth, A.; Kleuser, B.; Gulbins, E.; Schmitz, G.; Kornhuber, J. Alterations of plasma glycerophospholipid and sphingolipid species in male alcohol-dependent patients. Biochim. Biophys. Acta 2015, 1851, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.; Wegner, D.; Groß, S.; Bahls, M.; Dörr, M.; Felix, S.B.; Ittermann, T.; Oswald, S.; Nauck, M.; Friedrich, N.; et al. Data on subgroup specific baseline characteristics and serum sphingosine-1-phosphate concentrations in the Study of Health in Pomerania. Data Brief 2017, 11, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, M.; Charmas, M.; Długołęcka, B.; Górski, J. Exercise increases plasma levels of sphingoid base-1 phosphates in humans. Acta Physiol. 2011, 203, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Książek, M.; Charmas, M.; Klusiewicz, A.; Zabielski, P.; Długołęcka, B.; Chabowski, A.; Baranowski, M. Endurance training selectively increases high-density lipoprotein-bound sphingosine-1-phosphate in the plasma. Scand. J. Med. Sci. Sports 2018, 28, 57–64. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant | Age (years) | BMI (kg/m2) | Triglycerides (mg/dL) | Cholesterol (mg/dL) | HDL (mg/dL) | LDL (mg/dL) |
|---|---|---|---|---|---|---|
| 1 | 35 | 24.7 | 143 | 185 | 40 | 117 |
| 2 | 20 | 26.6 | 80 | 153 | 48 | 89 |
| 3 | 32 | 24.3 | 127 | 190 | 61 | 104 |
| 4 | 24 | 22.1 | 97 | 122 | 61 | 42 |
| +22 °C | |||||
|---|---|---|---|---|---|
| 0 h | 2 h | 4 h | 6 h | 8 h | |
| EDTA | |||||
| C16Cer | 100 ± 68.4 | 66.9 ± 35.4 | 96.3 ± 76.4 | 58.8 ± 12.0 | 65.7 ± 21.9 |
| C18Cer | 100 ± 32.0 | 86.1 ± 62.5 | 98.8 ± 44.1 | 69.8 ± 29.3 | 88.5 ± 42.7 |
| C20Cer | 100 ± 17.9 | 89.6 ± 33.8 | 105 ± 56.2 | 96.3 ± 54.9 | 98.7 ± 52.0 |
| C24Cer | 100 ± 17.9 | 92.9 ± 26.0 | 92.7 ± 27.0 | 92.0 ± 23.5 | 92.7 ± 24.3 |
| C24:1Cer | 100 ± 11.4 | 95.8 ± 35.3 | 98.7 ± 12.1 | 100 ± 34.1 | 99.1 ± 37.1 |
| C24dhCer | 100 ± 20.6 | 91.6 ± 20.2 | 103 ± 18.7 | 89.2 ± 16.0 | 101 ± 14.6 |
| C24:1dhCer | 100 ± 22.6 | 104 ± 26.4 | 103 ± 27.8 | 112 ± 43.0 | 108 ± 34.7 |
| Serum | |||||
| C16Cer | 100 ± 62.5 | 80.9 ± 45.8 | 62.9 ± 27.8 | 55.8 ± 30.2 | 102 ± 84.0 |
| C18Cer | 100 ± 111 | 82.3 ± 74.4 | 55.5 ± 25.0 | 42.3 ± 12.5 | 65.9 ± 17.0 |
| C20Cer | 100 ± 19.6 | 101 ± 79.6 | 85.7 ± 34.2 | 82.1 ± 18.0 | 105 ± 50.4 |
| C24Cer | 100 ± 24.6 | 113 ± 21.9 | 105 ± 25.4 | 99.6 ± 30.3 | 106 ± 23.6 |
| C24:1Cer | 100 ± 19.3 | 104 ± 28.3 | 88.8 ± 34.1 | 82.0 ± 30.1 | 110 ± 41.5 |
| C24dhCer | 100 ± 27.7 | 102 ± 28.8 | 102 ± 29.6 | 103 ± 47.7 | 101 ± 32.2 |
| C24:1dhCer | 100 ± 16.7 | 112 ± 22.0 | 97.9 ± 29.4 | 94.3 ± 31.0 | 117 ± 40.4 |
| 0 h | 4 h | ||
|---|---|---|---|
| - | +4 °C | +22 °C | |
| Heparin | |||
| C16Cer | 100 ± 34.5 | 88.3 ± 41.6 | 91.1 ± 23.5 |
| C18Cer | 100 ± 42.7 | 103 ± 44.4 | 120 ± 34.9 * |
| C20Cer | 100 ± 40.6 | 106 ± 46.5 | 91.5 ± 35.0 |
| C24Cer | 100 ± 23.9 | 102 ± 22.8 | 107 ± 25.5 |
| C24:1Cer | 100 ± 26.5 | 95.7 ± 17.8 | 105 ± 32.1 |
| C24dhCer | 100 ± 37.7 | 94.1 ± 26.1 | 94.6 ± 35.6 |
| C24:1dhCer | 100 ± 27.7 | 91.2 ± 24.5 | 100 ± 34.5 |
| EDTA | |||
| C16Cer | 100 ± 23.0 | 107 ± 38.6 | 91.9 ± 24.3 |
| C18Cer | 100 ± 49.1 | 99.1 ± 44.4 | 110 ± 44.1 |
| C20Cer | 100 ± 66.8 | 90.7 ± 32.2 | 102 ± 43.4 |
| C24Cer | 100 ± 25.9 | 102 ± 24.9 | 105 ± 25.8 |
| C24:1Cer | 100 ± 32.0 | 101 ± 25.7 | 106 ± 25.5 |
| C24dhCer | 100 ± 28.9 | 117 ± 36.2 | 98.1 ± 21.9 |
| C24:1dhCer | 100 ± 32.2 | 94.5 ± 23.5 | 103 ± 25.9 |
| Citrate | |||
| C16Cer | 100 ± 29.2 | 110 ± 21.4 | 102 ± 15.4 |
| C18Cer | 100 ± 50.1 | 104 ± 37.3 | 92.3 ± 37.6 |
| C20Cer | 100 ± 51.7 | 98.9 ± 47.5 | 101 ± 41.3 |
| C24Cer | 100 ± 26.0 | 105 ± 24.4 | 104 ± 24.4 |
| C24:1Cer | 100 ± 19.8 | 105 ± 16.7 | 99.5 ± 24.3 |
| C24dhCer | 100 ± 35.9 | 106 ± 26.8 | 117 ± 38.8 |
| C24:1dhCer | 100 ± 26.6 | 102 ± 18.2 | 100 ± 29.3 |
| Serum | |||
| C16Cer | 100 ± 45.4 | 106 ± 46.5 | 116 ± 43.3 |
| C18Cer | 100 ± 42.9 | 102 ± 43.9 | 101 ± 51.1 |
| C20Cer | 100 ± 40.8 | 98.1 ± 34.9 | 93.0 ± 34.2 |
| C24Cer | 100 ± 26.0 | 99.8 ± 21.6 | 99.9 ± 28.1 |
| C24:1Cer | 100 ± 22.3 | 100 ± 13.2 | 109 ± 36.9 |
| C24dhCer | 100 ± 39.4 | 102 ± 24.6 | 107 ± 50.8 |
| C24:1dhCer | 100 ± 31.6 | 90.8 ± 14.0 | 104 ± 34.1 |
| 0 h | 4 h | ||
|---|---|---|---|
| - | +4 °C | +22 °C | |
| C16Cer | 100 ± 31.1 | 113 ± 36.8 | 115 ± 44.2 |
| C18Cer | 100 ± 43.3 | 114 ± 39.2 | 98.5 ± 37.0 |
| C20Cer | 100 ± 52.3 | 110 ± 68.7 | 74.0 ± 30.9 |
| C24Cer | 100 ± 30.7 | 99.7 ± 20.7 | 95.3 ± 25.0 |
| C24:1Cer | 100 ± 40.7 | 110 ± 34.1 | 113 ± 52.4 |
| C24dhCer | 100 ± 29.7 | 99.5 ± 29.3 | 91.7 ± 25.2 |
| C24:1dhCer | 100 ± 41.9 | 106 ± 28.9 | 95.5 ± 35.5 |
| Blood vessel | RR + 10 mmHg | |||
|---|---|---|---|---|
| Vein | Artery | 1 min | 6 min | |
| C16Cer | 100 ± 10.2 | 106 ± 23.2 | 100 ± 20.1 | 93.4 ± 20.8 |
| C18Cer | 100 ± 10.4 | 108 ± 21.3 | 100 ± 25.6 | 93.5 ± 30.1 |
| C20Cer | 100 ± 7.56 | 116 ± 4.92 | 100 ± 41.2 | 96.4 ± 35.6 |
| C24Cer | 100 ± 10.5 | 76.1 ± 10.0 | 100 ± 30.4 | 95.9 ± 22.7 |
| C24:1Cer | 100 ± 11.0 | 109 ± 11.0 | 100 ± 20.6 | 93.2 ± 22.4 * |
| C24dhCer | 100 ± 32.3 | 124 ± 27.0 | 100 ± 35.5 | 94.9 ± 32.2 |
| C24:1dhCer | 100 ± 20.7 | 89.7 ± 13.6 | 100 ± 27.0 | 95.0 ± 22.1 |
| Centrifugation | Freezing | |||||
|---|---|---|---|---|---|---|
| 550× g | 5800× g | 16,200× g | Snap | −20 °C | −80 °C | |
| C16Cer | 100 ± 15.9 | 95.2 ± 17.5 | 93.6 ± 22.8 | 100 ± 36.0 | 112 ± 22.6 | 105 ± 23.0 |
| C18Cer | 100 ± 33.9 | 105 ± 50.0 | 96.7 ± 42.4 | 100 ± 50.2 | 108 ± 43.1 | 111 ± 41.1 |
| C20Cer | 100 ± 35.7 | 94.1 ± 48.3 | 116 ± 4.92 | 100 ± 59.5 | 104 ± 38.0 | 104 ± 52.8 |
| C24Cer | 100 ± 19.7 | 97.2 ± 24.4 | 92.8 ± 25.1 | 100 ± 37.0 | 101 ± 20.0 | 99.7 ± 24.1 |
| C24:1Cer | 100 ± 32.9 | 90.9 ± 25.9 | 89.9 ± 24.4 | 100 ± 36.4 | 103 ± 24.2 | 102 ± 29.0 |
| C24dhCer | 100 ± 17.0 | 92.6 ± 23.6 | 87.7 ± 24.1 | 100 ± 31.1 | 103 ± 21.9 | 103 ± 25.7 |
| C24:1dhCer | 100 ± 11.4 | 93.8 ± 13.2 | 91.0 ± 15.8 | 100 ± 22.1 | 103 ± 17.4 | 101 ± 14.0 |
| Freeze-Thaw Cycles | Storage (Months) | |||||
|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 0 | 18 | |
| C16Cer | 100 ± 13.8 | 105 ± 17.3 | 98.9 ± 23.6 | 107 ± 18.8 | 100 ± 35.3 | 92.6 ± 20.6 |
| C18Cer | 100 ± 24.7 | 103 ± 24.3 | 93.9 ± 21.3 | 106 ± 31.0 | 100 ± 26.5 | 117 ± 44.8 |
| C20Cer | 100 ± 21.4 | 106 ± 26.5 | 98.3 ± 17.5 | 109 ± 24.5 | 100 ± 27.9 | 89.9 ± 30.6 |
| C24Cer | 100 ± 33.1 | 108 ± 34.9 | 97.2 ± 34.3 | 110 ± 32.6 | 100 ± 28.7 | 96.9 ± 38.0 |
| C24:1Cer | 100 ± 27.1 | 106 ± 26.1 | 97.1 ± 21.3 | 108 ± 28.4 | 100 ± 21.6 | 98.6 ± 30.4 |
| C24dhCer | 100 ± 28.0 | 100 ± 29.3 | 93.2 ± 33.7 | 106 ± 32.2 | 100 ± 35.4 | 77.0 ± 37.3 |
| C24:1dhCer | 100 ± 26.0 | 105 ± 26.0 | 99.3 ± 28.9 | 113 ± 28.9 | 100 ± 25.0 | 97.1 ± 35.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brunkhorst, R.; Pfeilschifter, W.; Patyna, S.; Büttner, S.; Eckes, T.; Trautmann, S.; Thomas, D.; Pfeilschifter, J.; Koch, A. Preanalytical Biases in the Measurement of Human Blood Sphingolipids. Int. J. Mol. Sci. 2018, 19, 1390. https://doi.org/10.3390/ijms19051390
Brunkhorst R, Pfeilschifter W, Patyna S, Büttner S, Eckes T, Trautmann S, Thomas D, Pfeilschifter J, Koch A. Preanalytical Biases in the Measurement of Human Blood Sphingolipids. International Journal of Molecular Sciences. 2018; 19(5):1390. https://doi.org/10.3390/ijms19051390
Chicago/Turabian StyleBrunkhorst, Robert, Waltraud Pfeilschifter, Sammy Patyna, Stefan Büttner, Timon Eckes, Sandra Trautmann, Dominique Thomas, Josef Pfeilschifter, and Alexander Koch. 2018. "Preanalytical Biases in the Measurement of Human Blood Sphingolipids" International Journal of Molecular Sciences 19, no. 5: 1390. https://doi.org/10.3390/ijms19051390