Expanding the Utilization of Formalin-Fixed, Paraffin-Embedded Archives: Feasibility of miR-Seq for Disease Exploration and Biomarker Development from Biopsies with Clear Cell Renal Cell Carcinoma

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. RNA Yield and RNA Quality
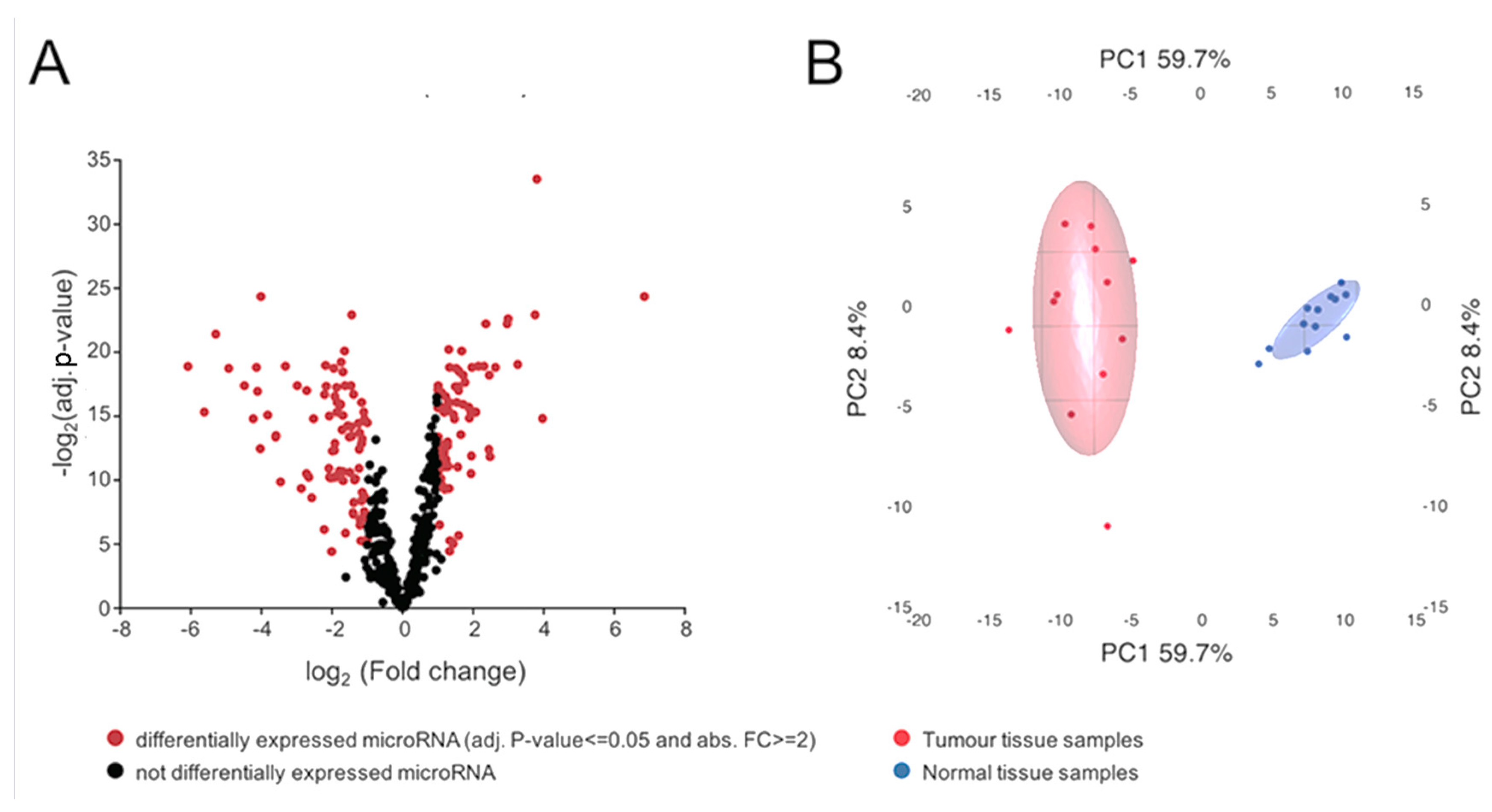
2.2. miRNA Expression Analysis and Data Visualization
2.3. Evaluation of Selected miRNAs as Potential Classifiers
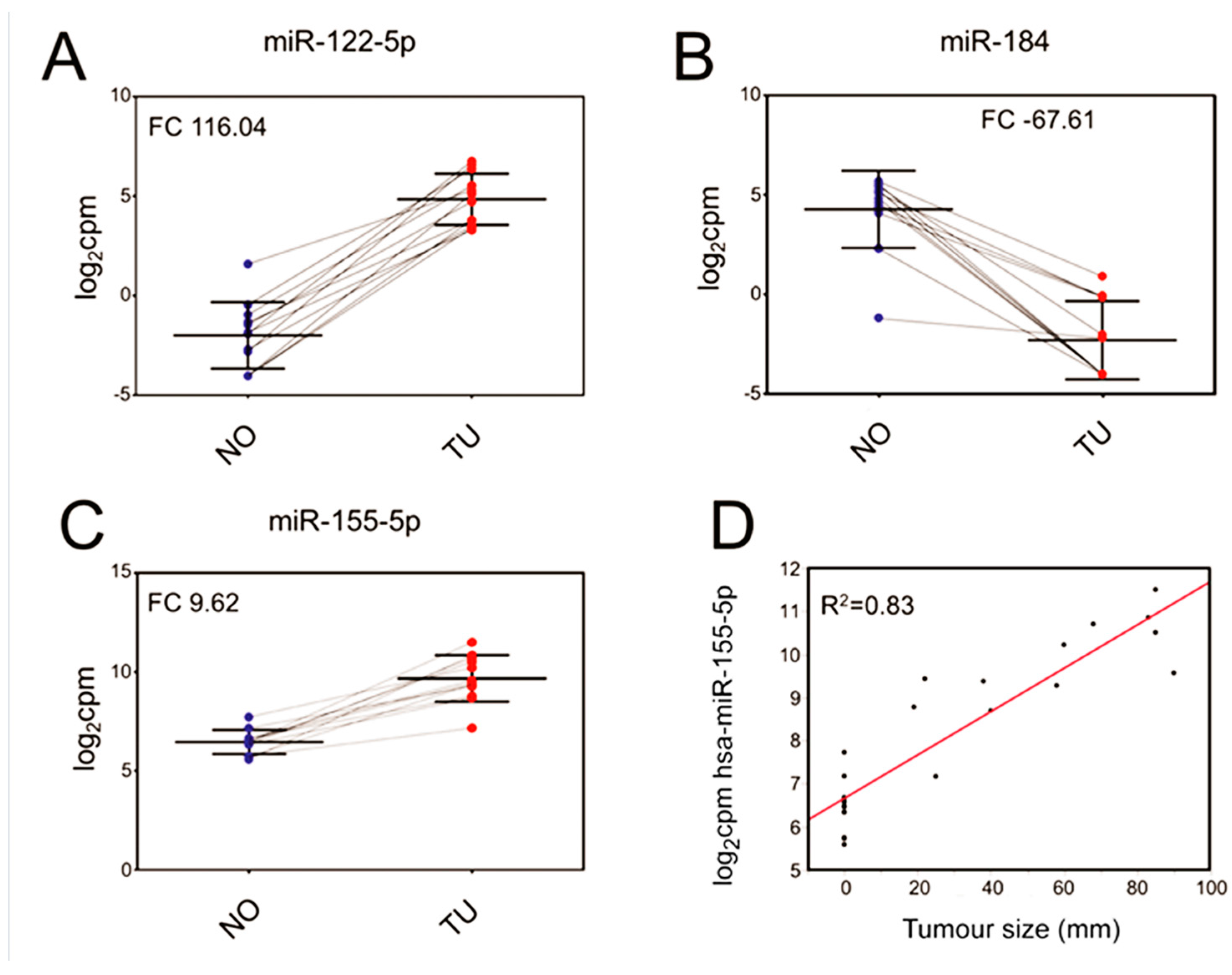
2.4. Correlation of miRNA with Tumour Size
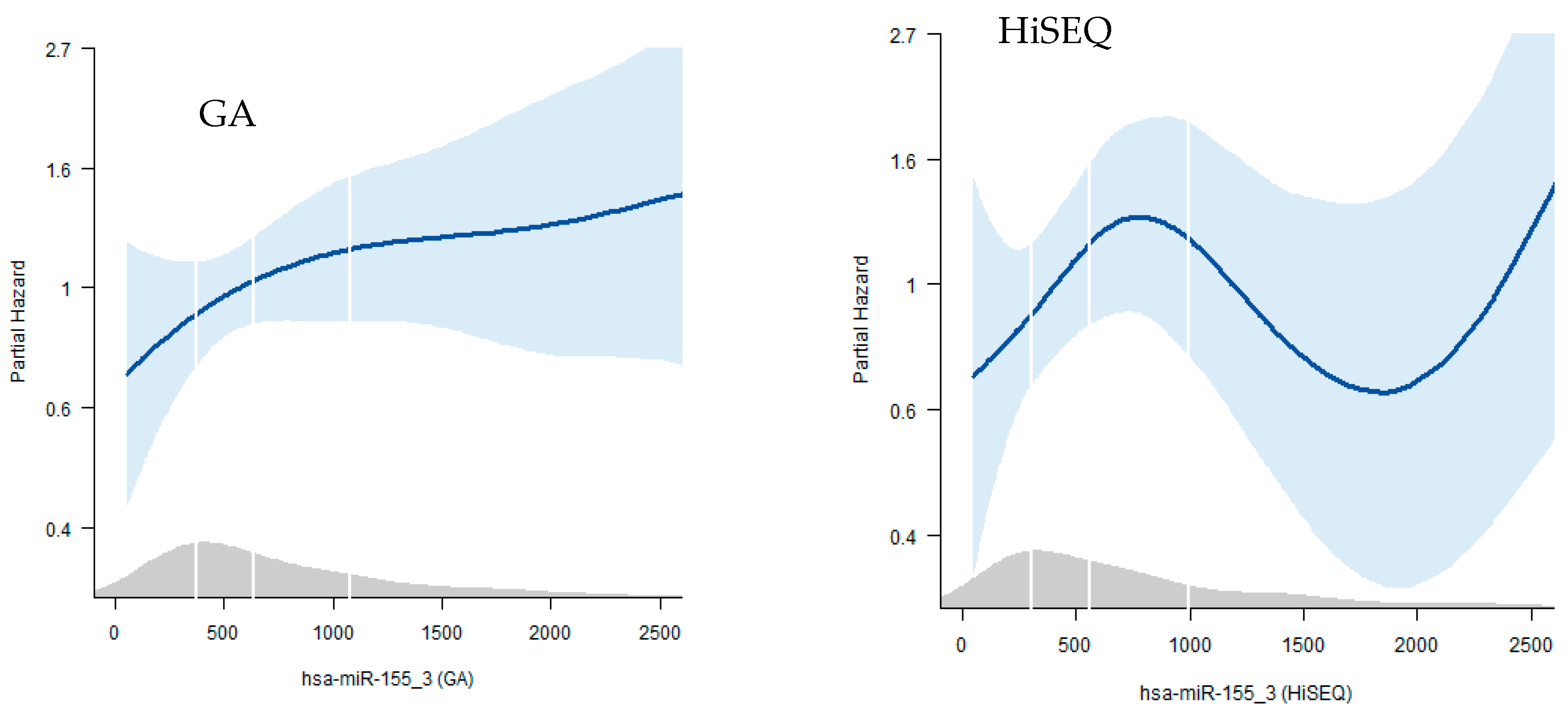
2.5. Survival Analysis
2.6. Correlation of miRNA Abundance to Body Mass Index (BMI)
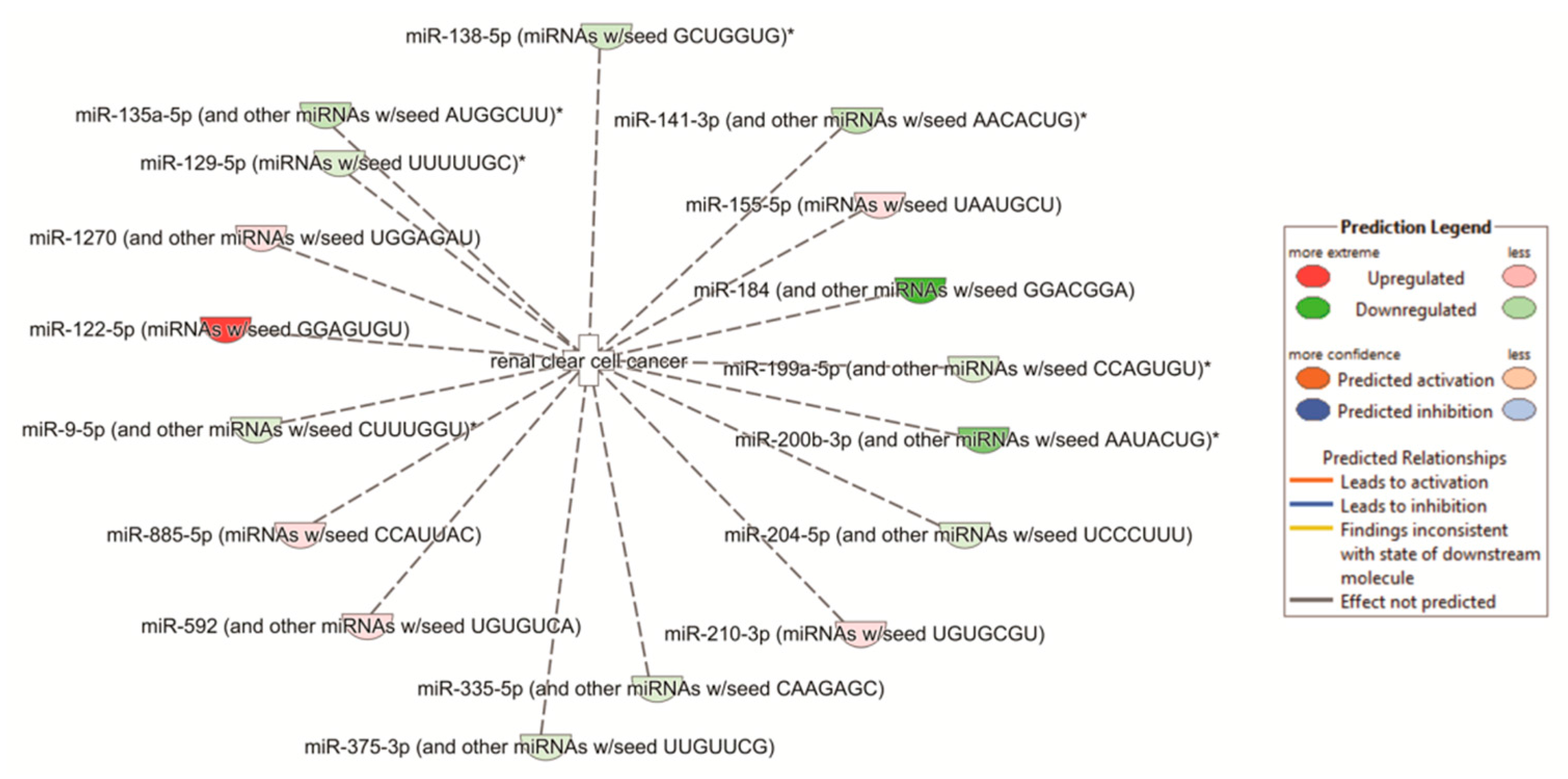
2.7. Pathway Analyses
2.8. Confirmation of Differentially Regulated miRNA
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Kidney Biopsies and RNA Extraction
4.3. Small RNA Library Preparation and Sequencing
4.4. Statistics and Next Generation Sequencing (NGS) Data Processing
4.5. Survival Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top miRNAs | Abs. Fold Change | Adj. p-Value | ||
|---|---|---|---|---|
| Tumour: BMI high vs. BMI low | miR-1251 | 10 | 0.440815 | |
| miR-483-3p | 9.5 | 0.740509 | ||
| mir-4792 | 8.7 | 0.151826 | ||
| miR-146b | 7.3 | 0.151826 | ||
| Normal: BMI high vs. low | mir-122 | 5.1 | 0.765518 | |
| miR-514a-5p | 4.7 | 0.185964 | ||
| miR-514a-3p | 4.9 | 0.085238 | ||
| 34c-3p | 3.9 | 0.765518 | ||
| BMI high: Tumour vs. Normal | miR-122-5p | 280 | 6.78 × 10−4 | |
| miR-184 | 129 | 3.15 × 10−5 | ||
| miR-122-3p | 50 | 1.95 × 10−4 | ||
| miR-891a-5p | 42.9 | 3.87 × 10−4 | ||
| BMI low: Tumour vs. Normal | mir-184 | 310.1 | 1.56 × 10−4 | |
| mir-891a | 137.5 | 2.19 × 10−3 | ||
| mir-141 | 63.9 | 2.30 × 10−4 | ||
| miR-122-5p | 58.8 | 7.11 × 10−4 | ||
| Tumour (BMI high vs. low) vs. Normal (BMI high vs. low) | mir-483-3p | 19.3 | 0.693293 | |
| mir-146b | 10.6 | 3.20 × 10−1 | ||
| mir-2277 | 9.9 | 3.20 × 10−1 | ||
| miR-192-5p | 6.1 | 4.39 × 10−1 |
| miRNA | Amongst 20 Most Deregulated miRNAs in Our Dataset | Evaluated as Classifier | Amongst the 4 miRNA with the Strongest Correlation to Tumour Size | Amongst the top 4 miRNAs with the Strongest Correlation to Tumour Size | Found amongst the Top 17 miRNAs Found by Shu et al. | Found amongst the Significant Differentially Expressed miRNAs from the Work of Shu et al. | Found amongst the Top 20 miRNAs Found by Osanto et al. | Found amongst the Significant Differentially Expressed miRNAs Found by Osanto et al. | Survival Analysis Using TCGA Dataset | Survival Analysis Using Illumina GA Dataset |
|---|---|---|---|---|---|---|---|---|---|---|
| (without Normal Samples) | (with Normal Samples) | |||||||||
| hsa-miR-122-5p | Yes | Yes | No | No | No | Yes | No | Yes | Yes | Yes |
| hsa-miR-885-5p | Yes | No | No | No | No | No | No | No | No | No |
| hsa-miR-210-3p | Yes | No | No | No | Yes | Yes | Yes | Yes | No | No |
| hsa-miR-210-5p | Yes | No | No | No | No | No | Yes | Yes | No | No |
| hsa-miR-138-5p | Yes | No | No | No | No | No | No | Yes | No | No |
| hsa-miR-187-3p | Yes | No | No | No | No | Yes | No | Yes | No | No |
| hsa-miR-4461 | Yes | No | No | No | No | No | No | No | No | No |
| hsa-miR-508-3p | Yes | No | No | No | No | No | No | No | No | No |
| hsa-miR-135a-5p | Yes | No | No | No | No | Yes | No | Yes | No | No |
| hsa-miR-129-1-3p | Yes | No | No | No | No | Yes | No | Yes | No | No |
| hsa-miR-141-3p | Yes | No | No | No | Yes | Yes | No | Yes | No | No |
| hsa-miR-216b-5p | Yes | No | No | No | No | No | No | No | No | No |
| hsa-miR-514a-3p | Yes | No | No | No | No | Yes | No | No | Yes | Yes |
| hsa-miR-141-5p | Yes | No | No | No | No | Yes | No | No | No | No |
| hsa-miR-200c-3p | Yes | No | No | No | Yes | Yes | No | Yes | No | No |
| hsa-miR-891a-5p | Yes | No | No | No | No | No | Yes | yes | No | No |
| hsa-miR-184 | Yes | Yes | No | No | No | No | No | No | Yes | Yes |
| hsa-miR-1304 | No | No | Yes | No | No | No | No | No | No | No |
| hsa-miR-155 | Yes | No | Yes | Yes | Yes | Yes | Yes | Yes | No | No |
| hsa-miR-142-3p | No | No | Yes | No | No | Yes | No | No | No | No |
| Hsa-miRNA-616-5p | No | No | Yes | No | No | No | No | No | No | No |
| Hsa-miRNA-361-3p | No | No | No | Yes | No | No | No | No | No | No |
| Hsa-miRNA-10b-3p | No | No | No | Yes | No | No | No | No | No | No |
| Hsa-miRNA-10b-5p | No | No | No | Yes | Yes | Yes | No | Yes | No | No |
| Hsa-miRNA-146 | No | No | No | No | No | Yes | No | Yes | Yes | Yes |
| Hsa-miRNA-362 | Yes | No | No | No | No | Yes | No | Yes | No | No |
| Hsa-miRNA-1251 | Yes | No | No | No | No | No | No | No | No | No |
References
- Fitzmaurice, C.; Dicker, D.; Pain, A.; Hamavid, H.; Moradi-Lakeh, M.; MacIntyre, M.F.; Allen, C.; Hansen, G.; Woodbrook, R.; Wolfe, C.; et al. The Global Burden of Cancer 2013. JAMA Oncol. 2015, 1, 505–527. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.T.; McGovern, F.J. Renal-cell carcinoma. N. Engl. J. Med. 2005, 353, 2477–2490. [Google Scholar] [CrossRef] [PubMed]
- Smittenaar, C.R.; Petersen, K.A.; Stewart, K.; Moitt, N. Cancer incidence and mortality projections in the UK until 2035. Br. J. Cancer 2016, 115, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Cairns, P. Renal cell carcinoma. Cancer Biomark. 2010, 9, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, B.; Campbell, S.C.; Choi, H.Y.; Jacqmin, D.; Lee, J.E.; Weikert, S.; Kiemeney, L.A. The epidemiology of renal cell carcinoma. Eur. Urol. 2011, 60, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Gerlinger, M.; Horswell, S.; Larkin, J.; Rowan, A.J.; Salm, M.P.; Varela, I.; Fisher, R.; McGranahan, N.; Matthews, N.; Santos, C.R.; et al. Genomic architecture and evolution of clear cell renal cell carcinomas defined by multiregion sequencing. Nat Genet. 2014, 46, 225–233. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Wang, L.; Zhou, W.; Zhang, Z.; Wang, L.; Xu, S.; Wang, D.; Dong, J.; Tang, C.; Tang, H.; et al. MicroRNA Expression Profiling in Clear Cell Renal Cell Carcinoma: Identification and Functional Validation of Key miRNAs. PLoS ONE 2015, 10, e0125672. [Google Scholar] [CrossRef] [PubMed]
- Guethmundsson, E.; Hellborg, H.; Lundstam, S.; Erikson, S.; Ljungberg, B.; Swedish Kidney Cancer Quality Register, G. Metastatic potential in renal cell carcinomas </=7 cm: Swedish Kidney Cancer Quality Register data. Eur. Urol. 2011, 60, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Thorstenson, A.; Harmenberg, U.; Lindblad, P.; Holmstrom, B.; Lundstam, S.; Ljungberg, B. Cancer Characteristics and Current Treatments of Patients with Renal Cell Carcinoma in Sweden. BioMed Res. Int. 2015, 2015, 456040. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, B.; Bensalah, K.; Canfield, S.; Dabestani, S.; Hofmann, F.; Hora, M.; Kuczyk, M.A.; Lam, T.; Marconi, L.; Merseburger, A.S.; et al. EAU guidelines on renal cell carcinoma: 2014 update. Eur. Urol. 2015, 67, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Coppin, C.; Kollmannsberger, C.; Le, L.; Porzsolt, F.; Wilt, T.J. Targeted therapy for advanced renal cell cancer (RCC): A Cochrane systematic review of published randomised trials. BJU Int. 2011, 108, 1556–1563. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chang, Y.; Xu, L.; Liu, Z.; Fu, Q.; Yang, Y.; Lin, Z.; Xu, J. The Presence of Vascular Mimicry Predicts High Risk of Clear Cell Renal Cell Carcinoma after Radical Nephrectomy. J. Urol. 2016, 196, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, D.R.; McCormack, R.T.; Keating, S.M.; Gutman, S.I.; Hamilton, S.R.; Mansfield, E.A.; Piper, M.A.; Deverka, P.; Frueh, F.W.; Jessup, J.M.; et al. Evidence of clinical utility: An unmet need in molecular diagnostics for patients with cancer. Clin. Cancer Res. 2014, 20, 1428–1444. [Google Scholar] [CrossRef] [PubMed]
- Heinzelmann, J.; Unrein, A.; Wickmann, U.; Baumgart, S.; Stapf, M.; Szendroi, A.; Grimm, M.O.; Gajda, M.R.; Wunderlich, H.; Junker, K. MicroRNAs with prognostic potential for metastasis in clear cell renal cell carcinoma: A comparison of primary tumors and distant metastases. Ann. Surg. Oncol. 2014, 21, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, T.; Seki, N.; Inoguchi, S.; Yoshino, H.; Tatarano, S.; Yamada, Y.; Itesako, T.; Goto, Y.; Nishikawa, R.; Nakagawa, M.; et al. Expression of the tumor suppressive miRNA-23b/27b cluster is a good prognostic marker in clear cell renal cell carcinoma. J. Urol. 2014, 192, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, I.Z.; Tan, Y.C.; Morozov, P.; Wilson, P.D.; Rennert, H.; Blumenfeld, J.D.; Tuschl, T. Urine microRNA as potential biomarkers of autosomal dominant polycystic kidney disease progression: Description of miRNA profiles at baseline. PLoS ONE 2014, 9, e86856. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Szabo, P.M.; Nofech-Mozes, R.; Rotondo, F.; Kovacs, K.; Mirham, L.; Girgis, H.; Boles, D.; Patocs, A.; Yousef, G.M. Integrative bioinformatics analysis reveals new prognostic biomarkers of clear cell renal cell carcinoma. Clin. Chem. 2014, 60, 1314–1326. [Google Scholar] [CrossRef] [PubMed]
- Fritz, H.K.; Lindgren, D.; Ljungberg, B.; Axelson, H.; Dahlback, B. The miR(21/10b) ratio as a prognostic marker in clear cell renal cell carcinoma. Eur. J. Cancer 2014, 50, 1758–1765. [Google Scholar] [CrossRef] [PubMed]
- Munari, E.; Marchionni, L.; Chitre, A.; Hayashi, M.; Martignoni, G.; Brunelli, M.; Gobbo, S.; Argani, P.; Allaf, M.; Hoque, M.O.; et al. Clear cell papillary renal cell carcinoma: Micro-RNA expression profiling and comparison with clear cell renal cell carcinoma and papillary renal cell carcinoma. Hum. Pathol. 2014, 45, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.; Wonnacott, A.; Fraser, D.J.; Bowen, T. MicroRNAs in Diabetic Nephropathy: From Biomarkers to Therapy. Curr. Diabetes Rep. 2016, 16, 35. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Anglicheau, D.; Muthukumar, T.; Suthanthiran, M. MicroRNAs: Small RNAs with big effects. Transplantation 2010, 90, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, K.; Kato, M.; Natarajan, R. Emerging Roles of microRNAs in the Pathophysiology of Renal Diseases. Am. J. Physiol. Ren. Physiol. 2015. [Google Scholar] [CrossRef]
- Li, M.; Wang, Y.; Song, Y.; Bu, R.; Yin, B.; Fei, X.; Guo, Q.; Wu, B. MicroRNAs in renal cell carcinoma: A systematic review of clinical implications (Review). Oncol. Rep. 2015, 33, 1571–1578. [Google Scholar] [CrossRef] [PubMed]
- Nassirpour, R.; Mathur, S.; Gosink, M.M.; Li, Y.; Shoieb, A.M.; Wood, J.; O’Neil, S.P.; Homer, B.L.; Whiteley, L.O. Identification of tubular injury microRNA biomarkers in urine: Comparison of next-generation sequencing and qPCR-based profiling platforms. BMC Genom. 2014, 15, 485. [Google Scholar] [CrossRef] [PubMed]
- Osanto, S.; Qin, Y.; Buermans, H.P.; Berkers, J.; Lerut, E.; Goeman, J.J.; van Poppel, H. Genome-wide microRNA expression analysis of clear cell renal cell carcinoma by next generation deep sequencing. PLoS ONE 2012, 7, e38298. [Google Scholar] [CrossRef] [PubMed]
- Weng, L.; Wu, X.; Gao, H.; Mu, B.; Li, X.; Wang, J.H.; Guo, C.; Jin, J.M.; Chen, Z.; Covarrubias, M.; et al. MicroRNA profiling of clear cell renal cell carcinoma by whole-genome small RNA deep sequencing of paired frozen and formalin-fixed, paraffin-embedded tissue specimens. J. Pathol. 2010, 222, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, J.; Thorsen, K.; Lund, M.K.; Hein, A.M.; Hamilton-Dutoit, S.J.; Vang, S.; Nordentoft, I.; Birkenkamp-Demtroder, K.; Kruhoffer, M.; Hager, H.; et al. Next-generation sequencing of RNA and DNA isolated from paired fresh-frozen and formalin-fixed paraffin-embedded samples of human cancer and normal tissue. PLoS ONE 2014, 9, e98187. [Google Scholar] [CrossRef] [PubMed]
- Eikrem, O.; Beisland, C.; Hjelle, K.; Flatberg, A.; Scherer, A.; Landolt, L.; Skogstrand, T.; Leh, S.; Beisvag, V.; Marti, H.P. Transcriptome Sequencing (RNAseq) Enables Utilization of Formalin-Fixed, Paraffin-Embedded Biopsies with Clear Cell Renal Cell Carcinoma for Exploration of Disease Biology and Biomarker Development. PLoS ONE 2016, 11, e0149743. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, C.P.; Feddersen, R.M.; Rakhshan, F.; Grill, D.E.; Simon, G.; Middha, S.; Jang, J.S.; Simon, V.; Schultz, D.A.; Zschunke, M.; et al. Multi-platform analysis of microRNA expression measurements in RNA from fresh frozen and FFPE tissues. PLoS ONE 2013, 8, e52517. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Xu, H. Prognostic value of meta-signature miRNAs in renal cell carcinoma: An integrated miRNA expression profiling analysis. Sci. Rep. 2015, 5, 10272. [Google Scholar] [CrossRef] [PubMed]
- Muciaccia, B.; Vico, C.; Aromatario, M.; Fazi, F.; Cecchi, R. Molecular analysis of different classes of RNA molecules from formalin-fixed paraffin-embedded autoptic tissues: A pilot study. Int. J. Legal Med. 2015, 129, 11–21. [Google Scholar] [CrossRef] [PubMed]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M.; Statistics Subcommittee of the, N.C.I.E.W.G.o.C.D. REporting recommendations for tumour MARKer prognostic studies (REMARK). Br. J. Cancer 2005, 93, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Park, B.; Jeong, B.C.; Seo, S.I.; Jeon, S.S.; Choi, H.Y.; Adami, H.O.; Lee, J.E.; Lee, H.M. Body mass index and survival in patients with renal cell carcinoma: A clinical-based cohort and meta-analysis. Int. J. Cancer 2013, 132, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Rogde, A.J.; Gudbrandsdottir, G.; Hjelle, K.M.; Sand, K.E.; Bostad, L.; Beisland, C. Obesity is associated with an improved cancer-specific survival, but an increased rate of postoperative complications after surgery for renal cell carcinoma. Scand. J. Urol. Nephrol. 2012, 46, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Hildebrandt, M.A.; Gu, J.; Tannir, N.M.; Matin, S.F.; Karam, J.A.; Wood, C.G.; Wu, X. MicroRNA profiling in clear cell renal cell carcinoma tissues potentially links tumorigenesis and recurrence with obesity. Br. J. Cancer 2017, 116, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Mollenkopf, H.J.; Grimm, C.; Wagner, I.; Albrecht, M.; Waller, T.; Pilarsky, C.; Johannsen, M.; Stephan, C.; Lehrach, H.; et al. MicroRNA profiling of clear cell renal cell cancer identifies a robust signature to define renal malignancy. J. Cell. Mol. Med. 2009, 13, 3918–3928. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.U.; Park, J.M.; Shin, S.J.; Lee, J.; Song, C.; Go, H.; Cho, N.H.; Ro, J.Y.; Cho, Y.M. Prognostic Significance of Macroscopic Appearance in Clear Cell Renal Cell Carcinoma and Its Metastasis-Predicting Model. Pathol. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Juan, D.; Alexe, G.; Antes, T.; Liu, H.; Madabhushi, A.; Delisi, C.; Ganesan, S.; Bhanot, G.; Liou, L.S. Identification of a microRNA panel for clear-cell kidney cancer. Urology 2010, 75, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Wotschofsky, Z.; Busch, J.; Jung, M.; Kempkensteffen, C.; Weikert, S.; Schaser, K.D.; Melcher, I.; Kilic, E.; Miller, K.; Kristiansen, G.; et al. Diagnostic and prognostic potential of differentially expressed miRNAs between metastatic and non-metastatic renal cell carcinoma at the time of nephrectomy. Clin. Chim. Acta 2013, 416, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Merhautova, J.; Hezova, R.; Poprach, A.; Kovarikova, A.; Radova, L.; Svoboda, M.; Vyzula, R.; Demlova, R.; Slaby, O. miR-155 and miR-484 Are Associated with Time to Progression in Metastatic Renal Cell Carcinoma Treated with Sunitinib. BioMed Res. Int. 2015, 2015, 941980. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ma, X.; Yao, Y.; Li, H.; Fan, Y.; Zhang, Y.; Zhao, C.; Wang, L.; Ma, M.; Lei, Z.; et al. miR-155 regulates the proliferation and invasion of clear cell renal cell carcinoma cells by targeting E2F2. Oncotarget 2016, 7, 20324–20337. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, T.; Zhong, Z.; Wang, Y.; Li, Y.; Zhao, X. microRNA-155 silencing inhibits proliferation and migration and induces apoptosis by upregulating BACH1 in renal cancer cells. Mol. Med. Rep. 2012, 5, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Tian, D.; Zhang, B.; Zhang, Y.; Yan, D.; Wu, S. Overexpression of miR-155 in clear-cell renal cell carcinoma and its oncogenic effect through targeting FOXO3a. Exp. Ther. Med. 2017, 13, 2286–2292. [Google Scholar] [CrossRef] [PubMed]
- Senbabaoglu, Y.; Gejman, R.S.; Winer, A.G.; Liu, M.; Van Allen, E.M.; de Velasco, G.; Miao, D.; Ostrovnaya, I.; Drill, E.; Luna, A.; et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Ma, X.; Li, H.; Gao, Y.; Huang, Q.; Zhang, Y.; Bao, X.; Du, Q.; Luo, G.; Liu, K.; et al. miR-122 Promotes Metastasis of Clear-Cell Renal Cell Carcinoma by Downregulating Dicer. Int. J. Cancer 2017. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yi, B.O.; Ding, S.; Sun, J.; Cao, W.; Liu, M. Demethylation drug 5-Aza-2′-deoxycytidine-induced upregulation of miR-200c inhibits the migration, invasion and epithelial-mesenchymal transition of clear cell renal cell carcinoma in vitro. Oncol. Lett. 2016, 11, 3167–3172. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, H.; Yonemori, M.; Miyamoto, K.; Tatarano, S.; Kofuji, S.; Nohata, N.; Nakagawa, M.; Enokida, H. microRNA-210-3p depletion by CRISPR/Cas9 promoted tumorigenesis through revival of TWIST1 in renal cell carcinoma. Oncotarget 2017, 8, 20881–20894. [Google Scholar] [CrossRef] [PubMed]
- Gjerdrum, C.; Tiron, C.; Hoiby, T.; Stefansson, I.; Haugen, H.; Sandal, T.; Collett, K.; Li, S.; McCormack, E.; Gjertsen, B.T.; et al. Axl is an essential epithelial-to-mesenchymal transition-induced regulator of breast cancer metastasis and patient survival. Proc. Natl. Acad. Sci. USA 2010, 107, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Hong, Q.; Li, O.; Zheng, W.; Xiao, W.Z.; Zhang, L.; Wu, D.; Cai, G.Y.; He, J.C.; Chen, X.M. LncRNA HOTAIR regulates HIF-1alpha/AXL signaling through inhibition of miR-217 in renal cell carcinoma. Cell Death Dis. 2017, 8, e2772. [Google Scholar] [CrossRef] [PubMed]
- Trivanovic, D.; Jaukovic, A.; Krstic, J.; Nikolic, S.; Okic Djordjevic, I.; Kukolj, T.; Obradovic, H.; Mojsilovic, S.; Ilic, V.; Santibanez, J.F.; et al. Inflammatory cytokines prime adipose tissue mesenchymal stem cells to enhance malignancy of MCF-7 breast cancer cells via transforming growth factor-beta1. IUBMB Life 2016, 68, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.Y.; Tang, S.J.; Chuang, M.J.; Cha, T.L.; Li, J.Y.; Sun, G.H.; Sun, K.H. TNF-alpha induces epithelial-mesenchymal transition of renal cell carcinoma cells via a GSK3beta-dependent mechanism. Mol. Cancer Res. 2012, 10, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.Y.; Han, Z.P.; Sun, K.; Zhang, S.S.; Hou, J.; Liu, Y.; Li, R.; Gao, L.; Zhao, X.; Zhao, Q.D.; et al. Toll-like receptor 4 signaling promotes epithelial-mesenchymal transition in human hepatocellular carcinoma induced by lipopolysaccharide. BMC Med. 2012, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tong, J.; He, F.; Yu, X.; Fan, L.; Hu, J.; Tan, J.; Chen, Z. miR-141 regulates TGF-beta1-induced epithelial-mesenchymal transition through repression of HIPK2 expression in renal tubular epithelial cells. Int. J. Mol. Med. 2015, 35, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.H.; Kao, A.P.; Lin, T.C.; Chang, C.C.; Kuo, T.C. Promotion of epithelial-mesenchymal transition and tumor growth by 17beta-estradiol in an ER(+)/HER2(+) cell line derived from human breast epithelial stem cells. Biotechnol. Appl. Biochem. 2012, 59, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Eikrem, O.S.; Strauss, P.; Beisland, C.; Scherer, A.; Landolt, L.; Flatberg, A.; Leh, S.; Beisvag, V.; Skogstrand, T.; Hjelle, K.; et al. Development and confirmation of potential gene classifiers of human clear cell renal cell carcinoma using next-generation RNA sequencing. Scand. J. Urol. 2016, 50, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Landolt, L.; Marti, H.P.; Beisland, C.; Flatberg, A.; Eikrem, O.S. RNA extraction for RNA sequencing of archival renal tissues. Scand. J. Clin. Lab. Investig. 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Walther, C.; Hofvander, J.; Nilsson, J.; Magnusson, L.; Domanski, H.A.; Gisselsson, D.; Tayebwa, J.; Doyle, L.A.; Fletcher, C.D.; Mertens, F. Gene fusion detection in formalin-fixed paraffin-embedded benign fibrous histiocytomas using fluorescence in situ hybridization and RNA sequencing. Lab. Investig. 2015, 95, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Goldfischer, M.; Babyeva, S.; Mao, Y.; Volyanskyy, K.; Dimitrova, N.; Fallon, J.T.; Zhong, M. Identification of a novel PARP14-TFE3 gene fusion from 10-year-old FFPE tissue by RNA-seq. Genes Chromosomes Cancer 2015. [Google Scholar] [CrossRef] [PubMed]
- Illumina. Evaluating RNA Quality from FFPE Samples. Available online: https://www.illumina.com/content/dam/illumina-marketing/documents/products/technotes/evaluating-rna-quality-from-ffpe-samples-technical-note-470-2014-001.pdf (accessed on 26 October 2016).
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]

| Patient Number | Age (Year) | Gender | BMI | Nephrectomy Type | eGFR | TNM-Stage | Size (mm) | Fuhrman Grade | Leibovich Score | Stage |
|---|---|---|---|---|---|---|---|---|---|---|
| 39 | 71 | Male | 25 | Radical | 59 | pT3AcN0cM0 | 90 | 4 | 8 | III |
| 44 | 74 | Female | 23 | Radical | >60 | pT3AcN0cM0 | 58 | 4 | 4 | III |
| 46 | 53 | Female | 24 | Partial | >60 | pT1AcN0cM0 | 38 | 1 | 0 | I |
| 50 | 72 | Female | 19 | Radical | >60 | pT1BcN0cM0 | 68 | 2 | 3 | I |
| 53 | 46 | Female | 44 | Radical | >60 | pT2AcN0cM0 | 83 | 2 | 3 | II |
| 55 | 44 | Female | 23 | Radical | >60 | pT3AcN0cM0 | 85 | 3 | 5 | III |
| 57 | 63 | Female | 28 | Radical | >60 | pT1AcN0cM0 | 25 | 2 | 0 | I |
| 59 | 52 | Female | 29 | Partial | >60 | pT1AcN0cM0 | 40 | 2 | 0 | I |
| 63a | 55 | Male | 28 | Partial | >60 | pT1AcN0cM0 | 19 | 3 | 1 | I |
| 63b | 44 | Male | 20 | Partial | >60 | pT1AcN0cM0 | 22 | 2 | 0 | I |
| 64 | 52 | Male | 26 | Radical | >60 | pT1BcN0cM0 | 60 | 3 | 4 | I |
| 65 | 57 | Male | 24 | Radical | >60 | pT2AcN0cM0 | 85 | 3 | 5 | II |
| Mature microRNA | Precursor microRNA | Fold Change (TU/NO) | p-Value | Adjusted p-Value |
|---|---|---|---|---|
| hsa-miR-122-5p | hsa-miR-122 | 116.04 | 2.60 × 10−10 | 4.68 × 10−8 |
| hsa-miR-184 | hsa-miR-184 | −67.61 | 8.22 × 10−8 | 2.05 × 10−6 |
| hsa-miR-891a-5p | hsa-miR-891a | −49.12 | 3.95 × 10−6 | 2.43 × 10−5 |
| hsa-miR-200c-3p | hsa-miR-200c | −39.12 | 6.84 × 10−9 | 3.59 × 10−7 |
| hsa-miR-141-5p | hsa-miR-141 | −30.31 | 1.25 × 10−7 | 2.29 × 10−6 |
| hsa-miR-514a-3p | hsa-miR-514a-2 | −22.31 | 4.82 × 10−7 | 5.87 × 10−6 |
| hsa-miR-216b-5p | hsa-miR-216b | −18.72 | 6.35 × 10−6 | 3.50 × 10−5 |
| hsa-miR-141-3p | hsa-miR-141 | −17.64 | 1.08 × 10−7 | 2.18 × 10−6 |
| hsa-miR-129-1-3p | hsa-miR-129-1 | −17.21 | 7.85 × 10−7 | 7.91 × 10−6 |
| hsa-miR-135a-5p | hsa-miR-135a-2 | −16.31 | 4.17 × 10−5 | 1.77 × 10−4 |
| hsa-miR-508-3p | hsa-miR-508 | −16.13 | 3.20 × 10−10 | 4.68 × 10−8 |
| hsa-miR-4461 | hsa-miR-4461 | −16.04 | 3.57 × 10−10 | 4.68 × 10−8 |
| hsa-miR-885-5p | hsa-miR-885 | 15.65 | 6.08 × 10−6 | 3.46 × 10−5 |
| hsa-miR-187-3p | hsa-miR-187 | −14.12 | 4.83 × 10−6 | 2.88 × 10−5 |
| hsa-miR-210-3p | hsa-miR-210 | 14.02 | 1.55 × 10−13 | 8.12 × 10−11 |
| hsa-miR-210-5p | hsa-miR-210 | 13.47 | 1.38 × 10−9 | 1.26 × 10−7 |
| hsa-miR-138-5p | hsa-miR-138-2 | −12.09 | 2.01 × 10−5 | 9.40 × 10−5 |
| hsa-miR-1251-5p | hsa-miR-1251 | −10.98 | 3.74 × 10−4 | 1.08 × 10−3 |
| hsa-miR-362-5p | hsa-miR-362 | −9.99 | 7.65 × 10−8 | 2.04 × 10−6 |
| hsa-miR-155-5p | hsa-miR-155 | 9.62 | 5.31 × 10−8 | 1.86 × 10−6 |
| A | GA | |||
| Model 1 | Model 2 | |||
| HR (95% CI) | p | HR | p | |
| Per SD | 1.18 (1.04, 1.35) | 0.013 | 1.0 (0.82, 1.21) | 0.97 |
| Vs Q1 | ||||
| Q2 | 1.88 (0.92, 3.83) | 0.082 | 1.39 (0.7, 2.77) | 0.342 |
| Q3 | 2.19 (1.12, 4.28) | 0.022 | 1.22 (0.62, 2.39) | 0.567 |
| Q4 | 3.03 (1.57, 5.85) | 0.001 | 1.46 (0.76, 2.8) | 0.261 |
| Model 1 | age, sex | |||
| model 2 | +stage, grade | |||
| B | HiSEQ | |||
| Model 1 | Model 2 | |||
| HR (95% CI) | p | HR | p | |
| Per SD | 1.27 (1.00, 1.62) | 0.046 | 1.1 (0.85–1.43) | 0.449 |
| Vs Q1 | ||||
| Q2 | 0.98 (0.42, 2.27) | 0.955 | 1.0 (0.42, 2.39) | 0.996 |
| Q3 | 1.84 (0.84, 4.01) | 0.128 | 2.4 (1.07, 5.37) | 0.033 |
| Q4 | 1.67 (0.79, 3.53) | 0.181 | 1.07 (0.5, 2.3) | 0.862 |
| Model 1 | age, sex | |||
| model 2 | +stage, grade | |||
| Pathways | p-Value | Overlap |
| Th2 pathway | 6.23 × 10−11 | 17.3% 26/150 |
| Th1 and Th2 activation pathway | 3.10 × 10−10 | 15.1% 28/185 |
| Th1 pathway | 3.12 × 10−8 | 15.6% 21/135 |
| Antigen presentation pathway | 9.02 × 10−8 | 28.9% 11/38 |
| Hepatic fibrosis/hepatic stellate cell activation | 3.88 × 10−7 | 12.6% 23/183 |
| Upstream Regulators | p-Value | |
| IFNG | 3.41 × 10−20 | |
| TNF | 7.26 × 10−16 | |
| Lipopolysaccharide TGG | 2.01 × 10−15 | |
| TGFβ1 | 1.03 × 10−14 | |
| Beta-estradiol | 1.12 × 10−14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strauss, P.; Marti, H.-P.; Beisland, C.; Scherer, A.; Lysne, V.; Leh, S.; Flatberg, A.; Koch, E.; Beisvag, V.; Landolt, L.; et al. Expanding the Utilization of Formalin-Fixed, Paraffin-Embedded Archives: Feasibility of miR-Seq for Disease Exploration and Biomarker Development from Biopsies with Clear Cell Renal Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 803. https://doi.org/10.3390/ijms19030803
Strauss P, Marti H-P, Beisland C, Scherer A, Lysne V, Leh S, Flatberg A, Koch E, Beisvag V, Landolt L, et al. Expanding the Utilization of Formalin-Fixed, Paraffin-Embedded Archives: Feasibility of miR-Seq for Disease Exploration and Biomarker Development from Biopsies with Clear Cell Renal Cell Carcinoma. International Journal of Molecular Sciences. 2018; 19(3):803. https://doi.org/10.3390/ijms19030803
Chicago/Turabian StyleStrauss, Philipp, Hans-Peter Marti, Christian Beisland, Andreas Scherer, Vegard Lysne, Sabine Leh, Arnar Flatberg, Even Koch, Vidar Beisvag, Lea Landolt, and et al. 2018. "Expanding the Utilization of Formalin-Fixed, Paraffin-Embedded Archives: Feasibility of miR-Seq for Disease Exploration and Biomarker Development from Biopsies with Clear Cell Renal Cell Carcinoma" International Journal of Molecular Sciences 19, no. 3: 803. https://doi.org/10.3390/ijms19030803