The Coordinated Activities of nAChR and Wnt Signaling Regulate Intestinal Stem Cell Function in Mice




Abstract
:1. Introduction
2. Results
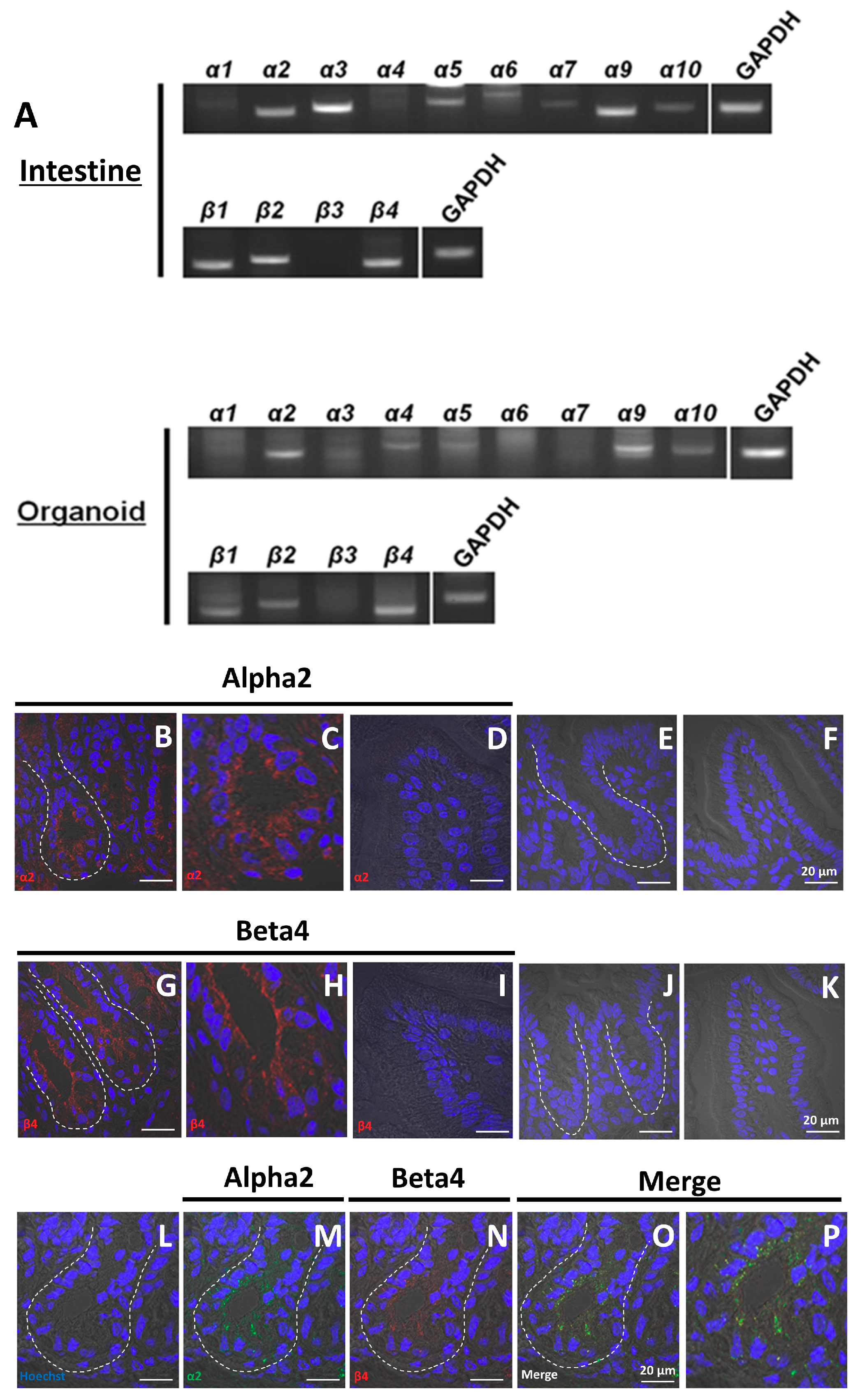
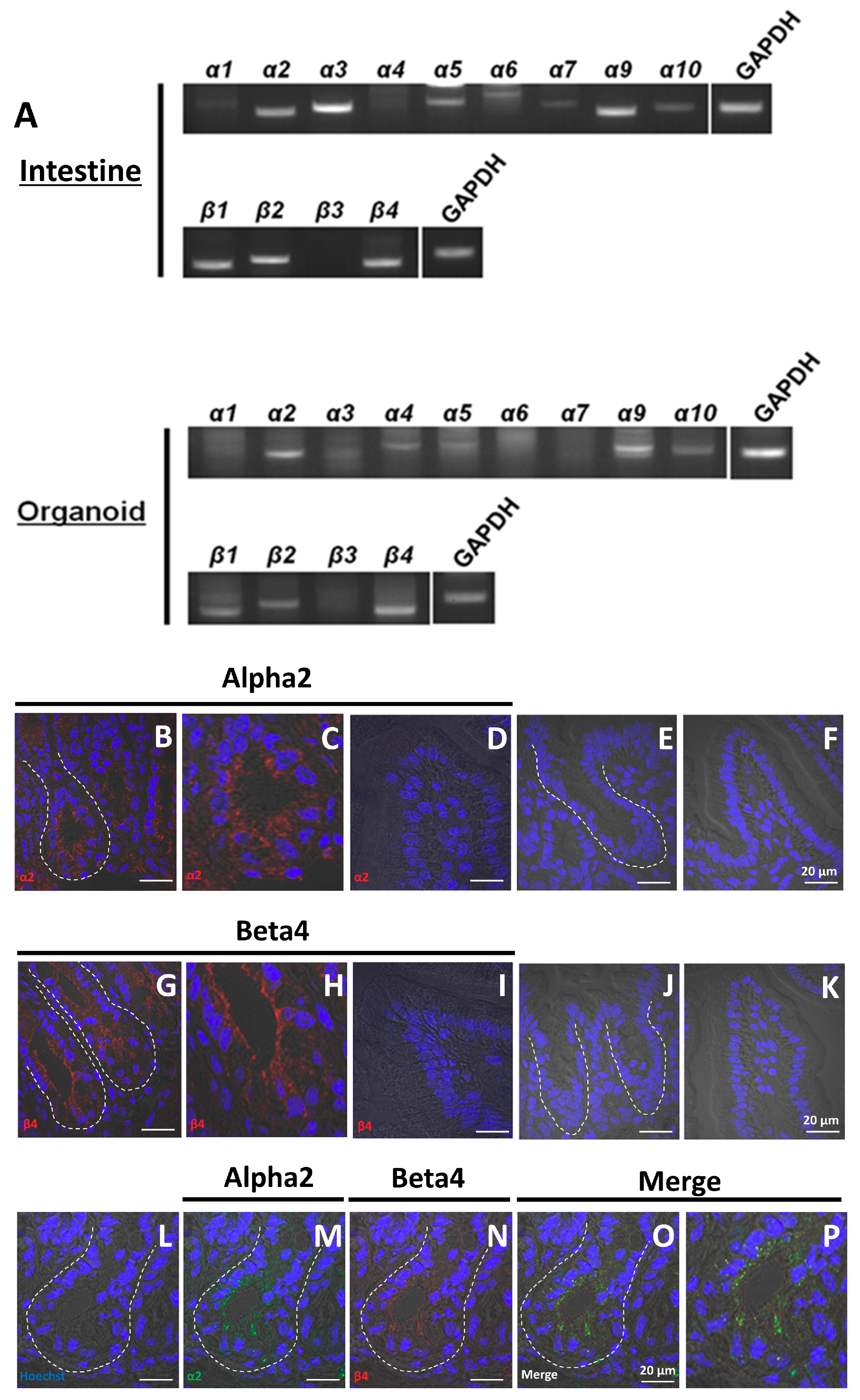
2.1. Organoids Contain Non-Neuronal nAChRs
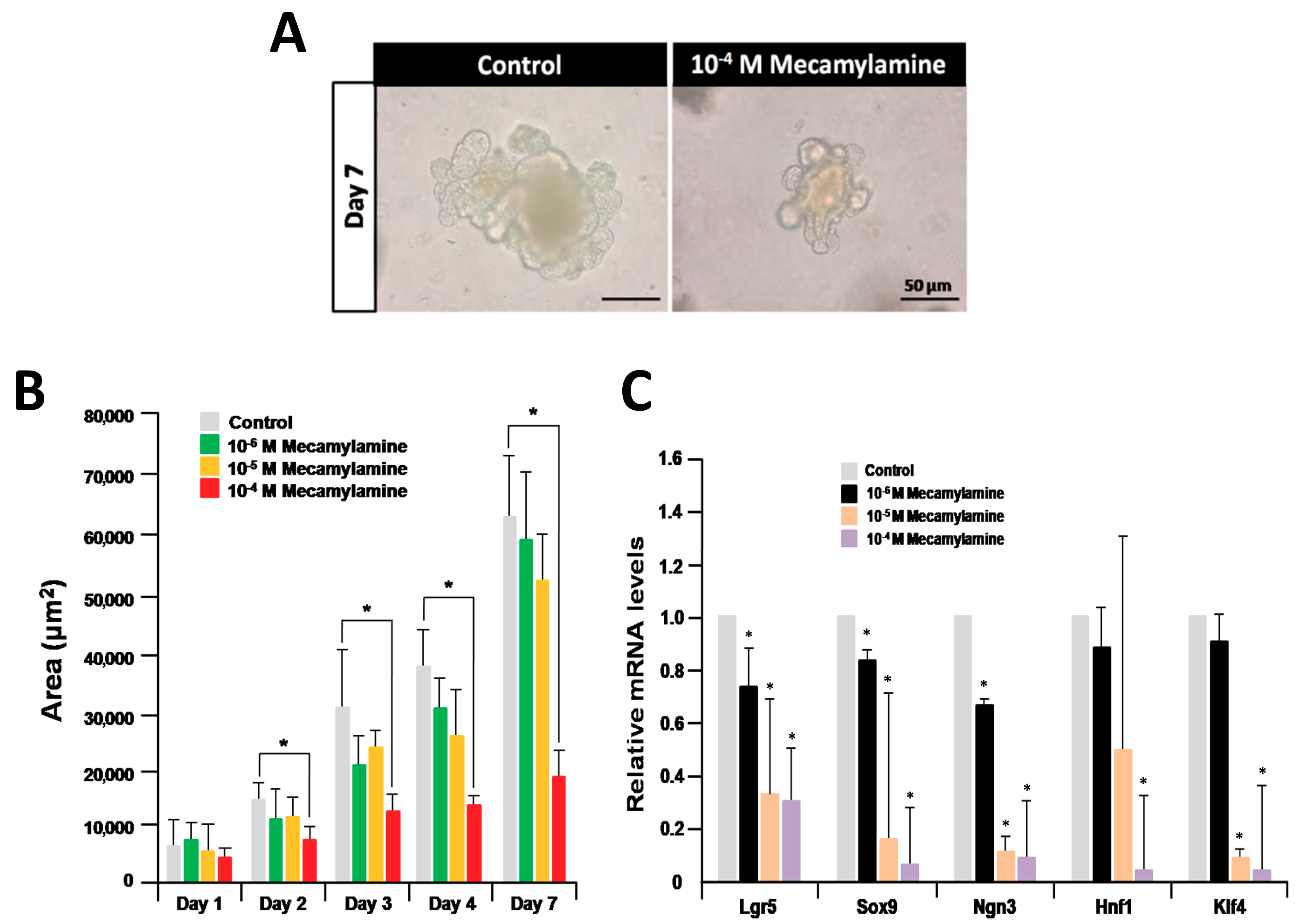
2.2. Inhibition of Organoid Growth and Differentiation by a nAChR Antagonist
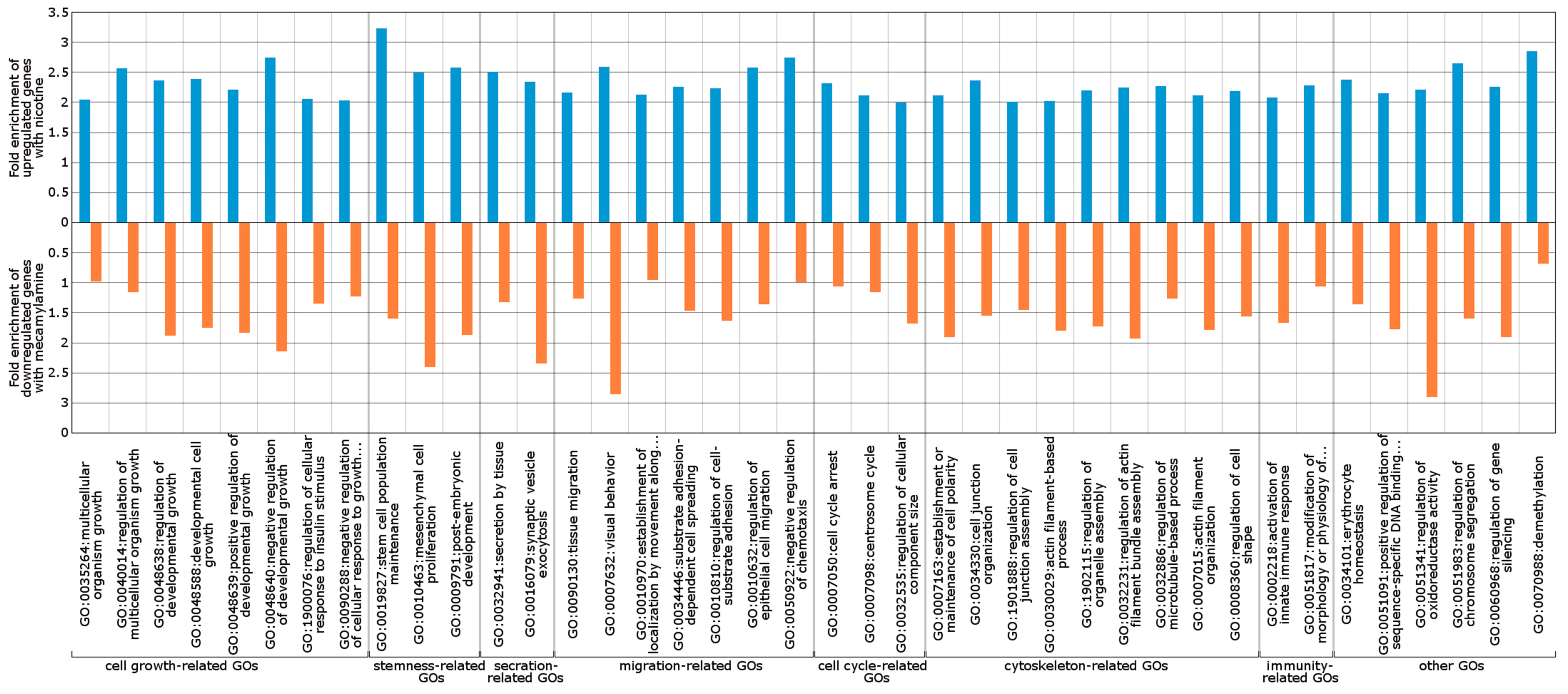
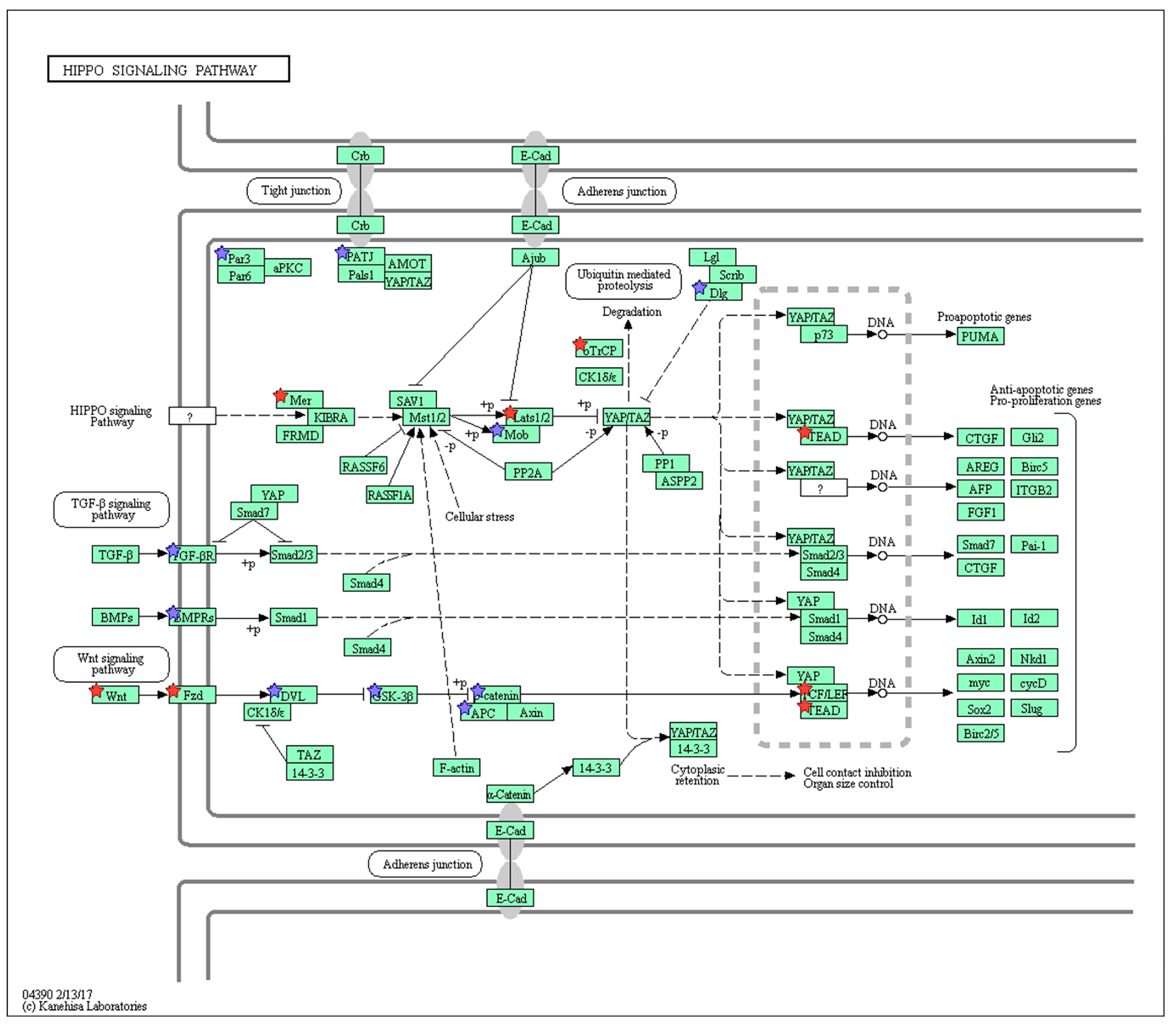
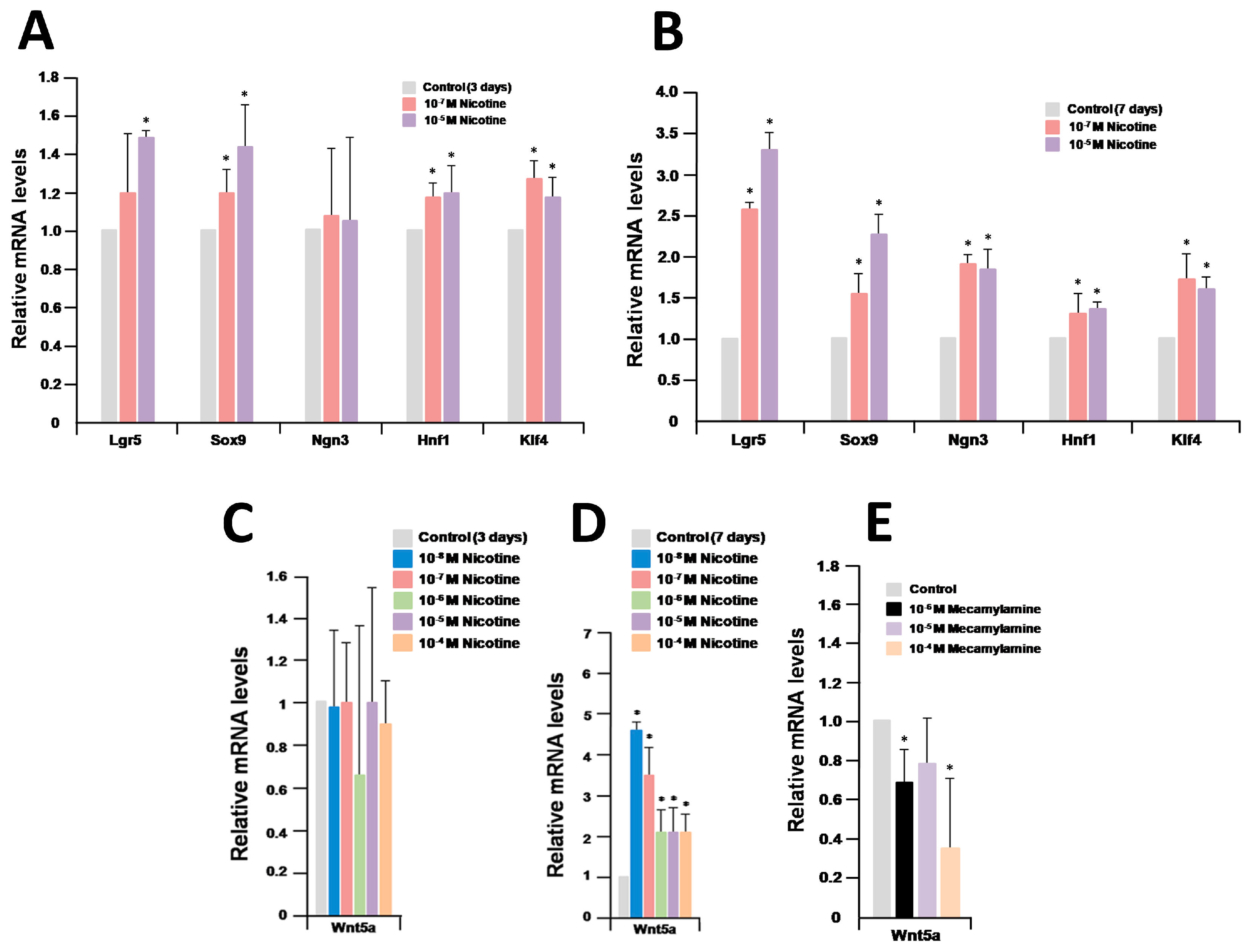
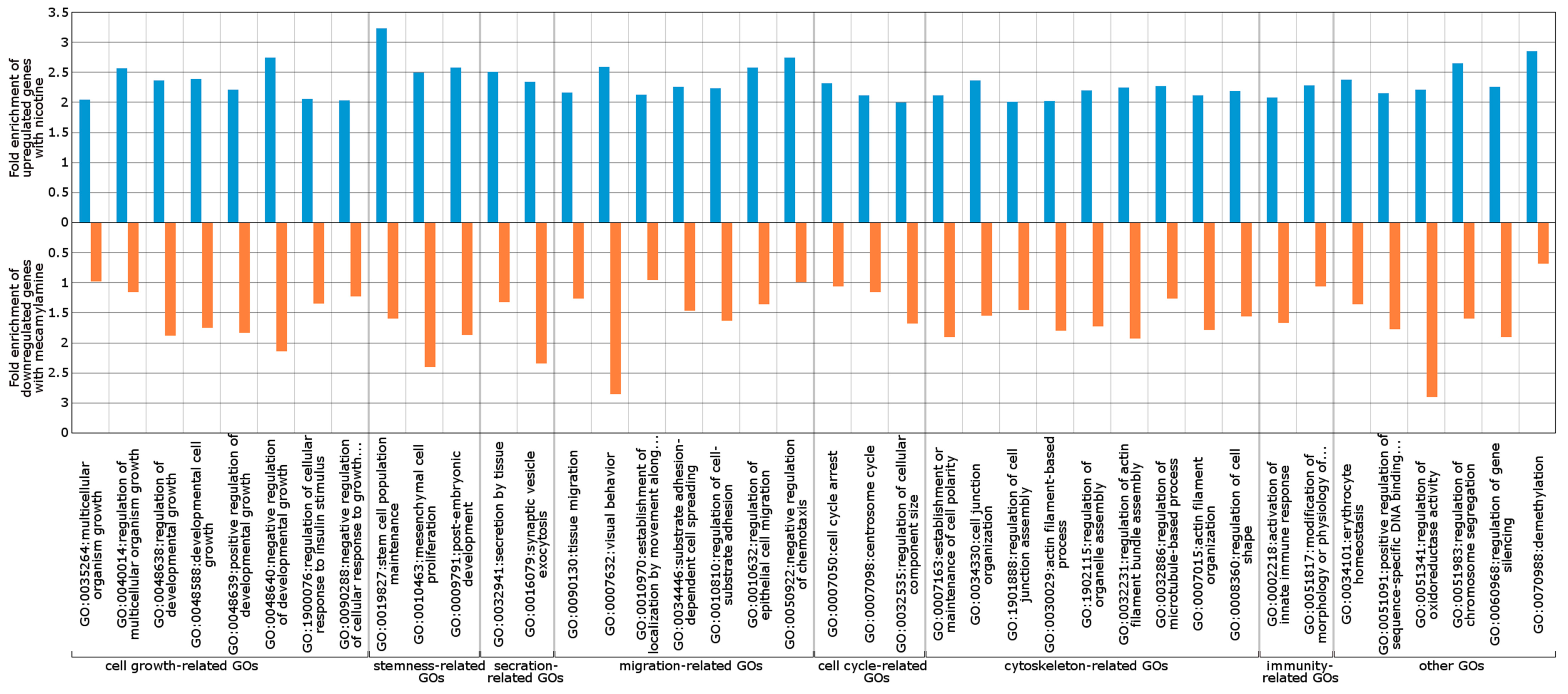
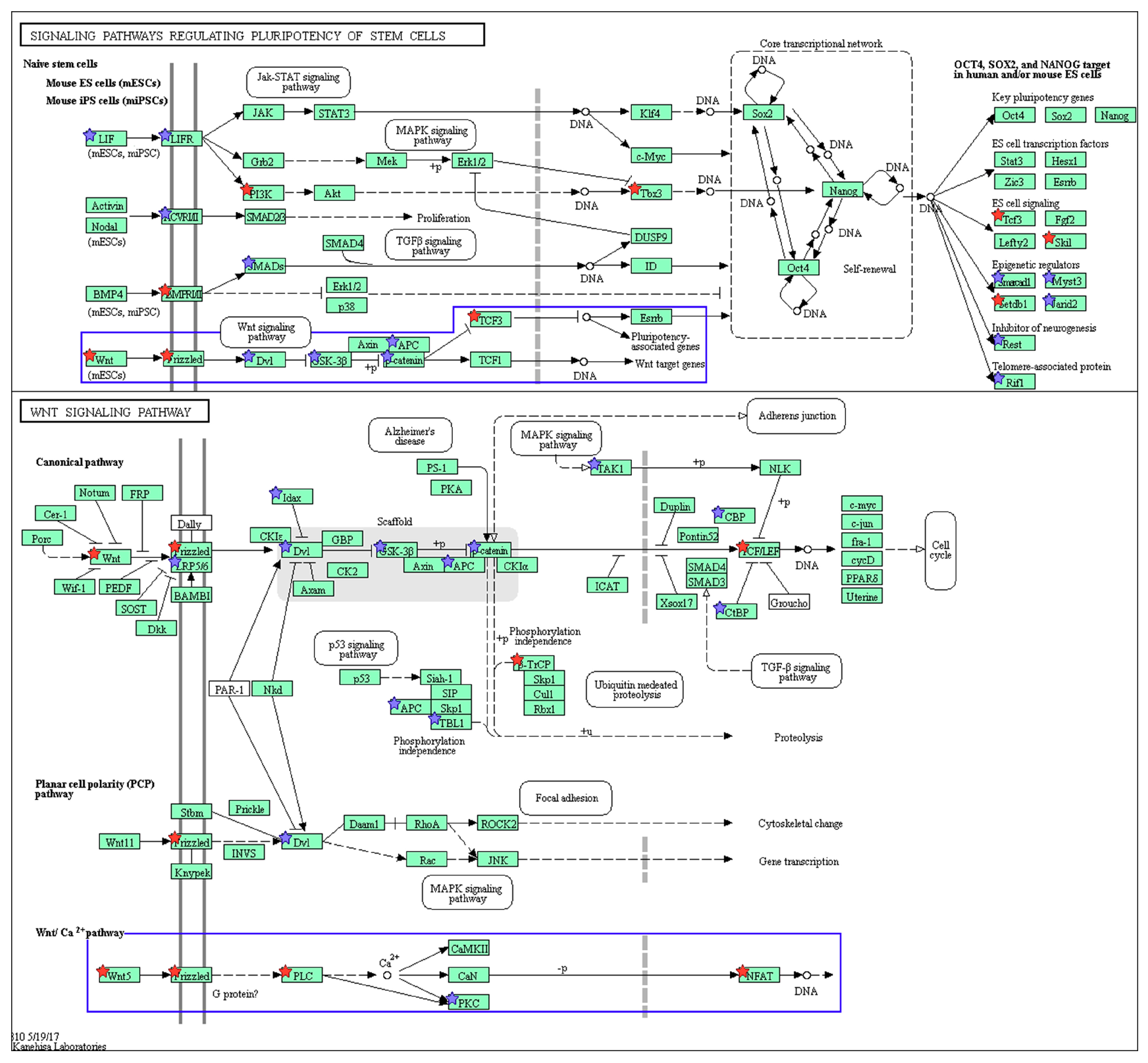
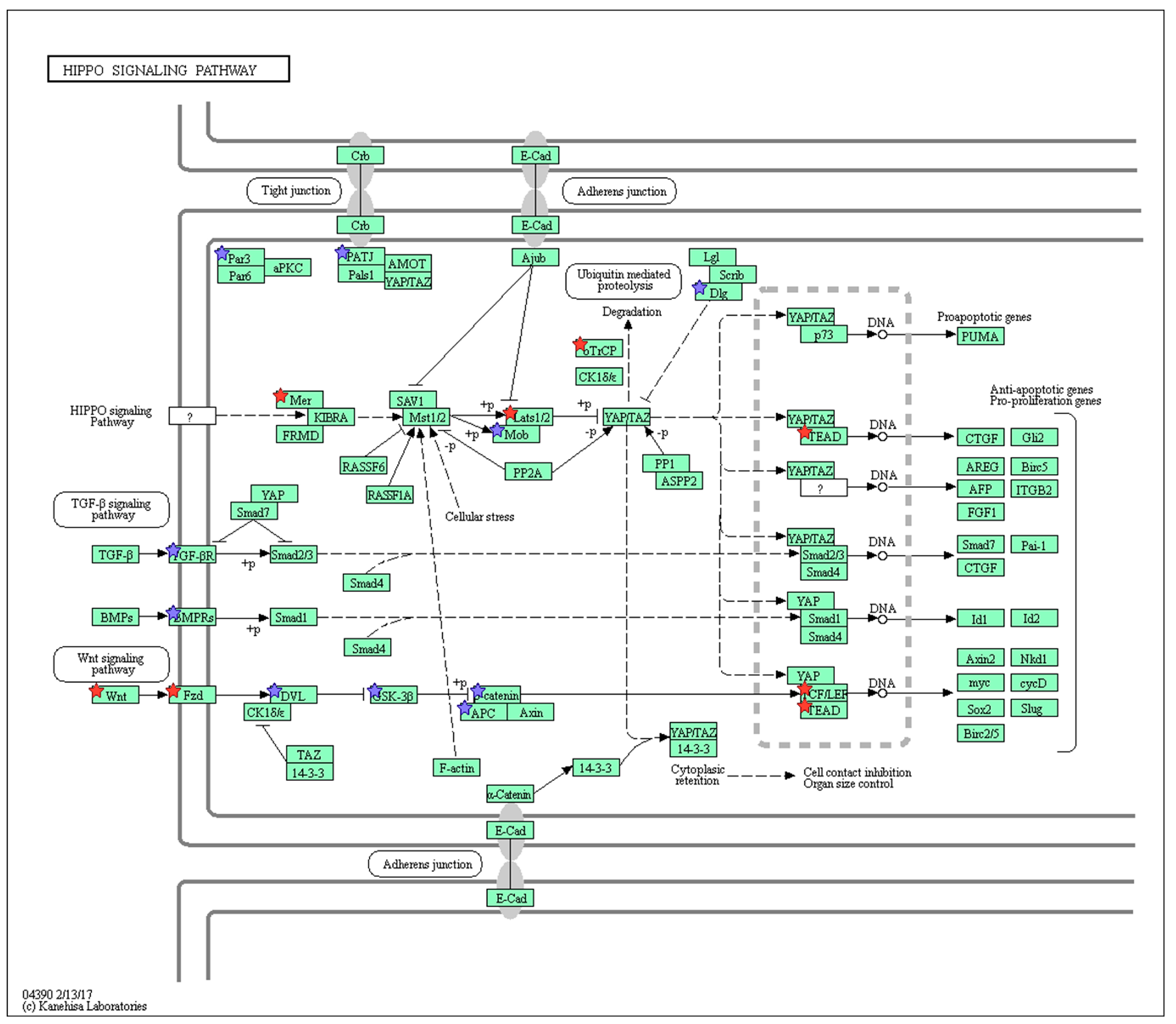
2.3. Wnt/Frizzled Pathway Activation Is Altered by Nicotine and Mecamylamine Treatment of Organoids
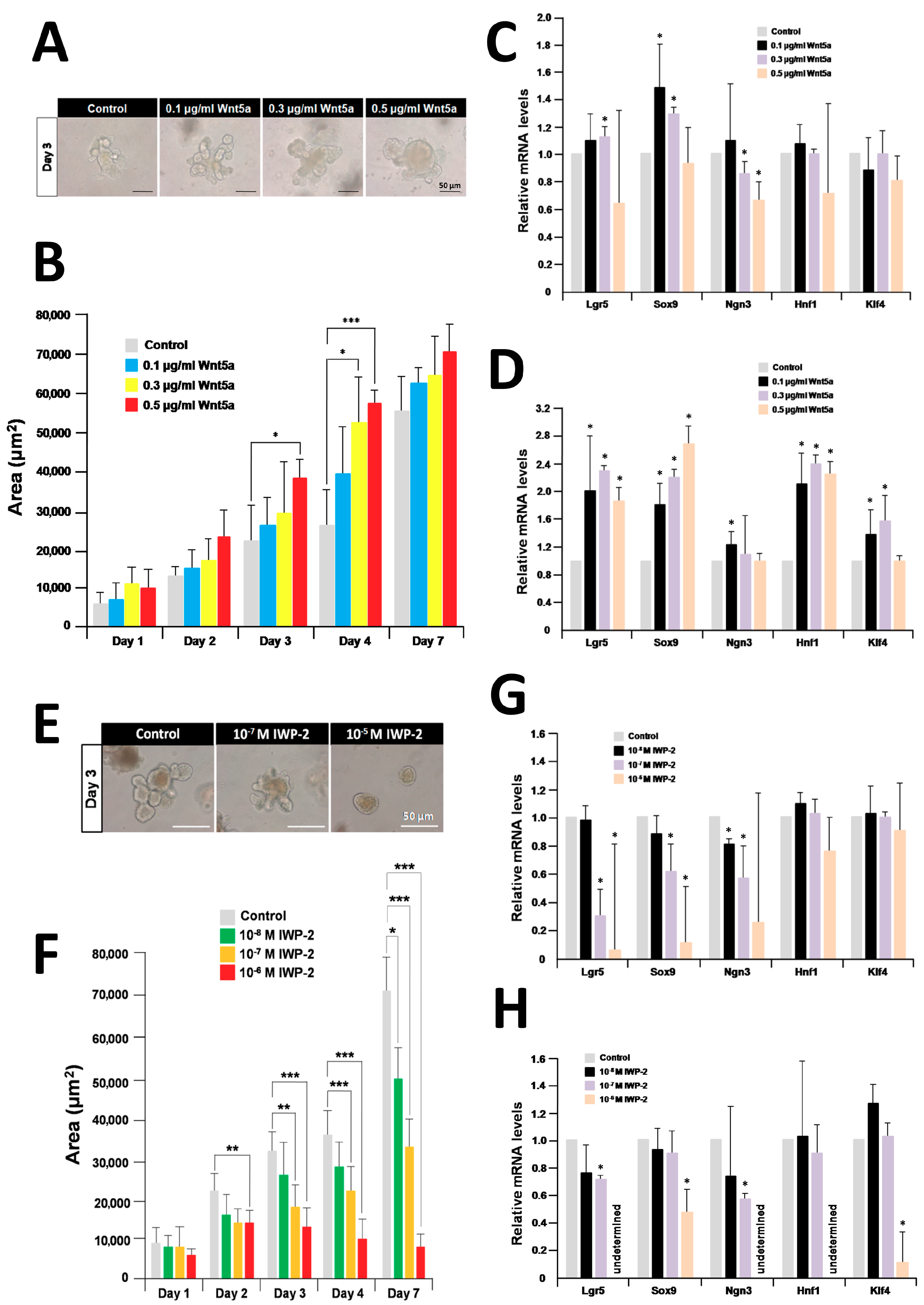
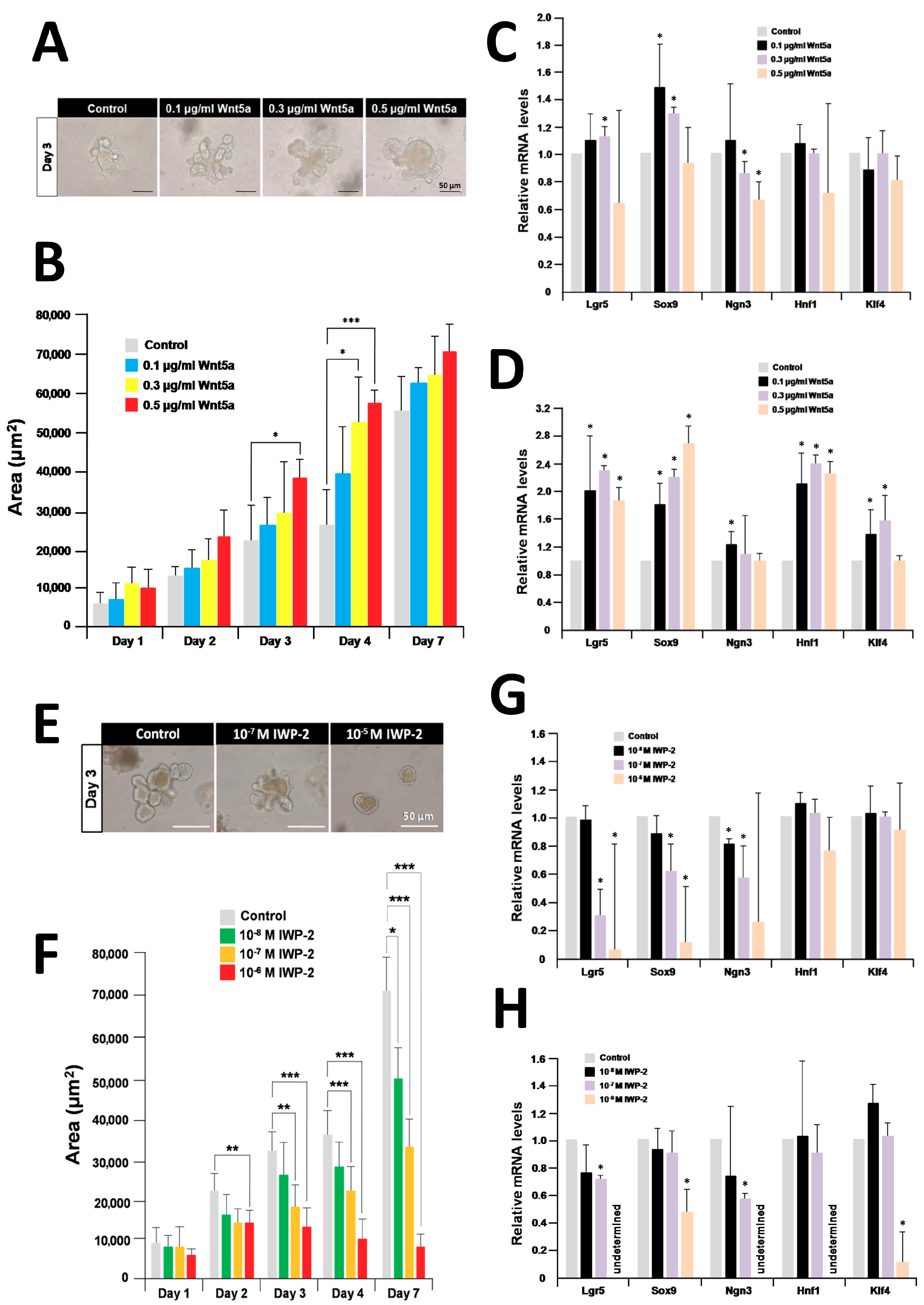
2.4. Recombinant Wnt5a Enhances Organoid Growth and Differentiation and Its Effect Is Inhibited By IWP-2
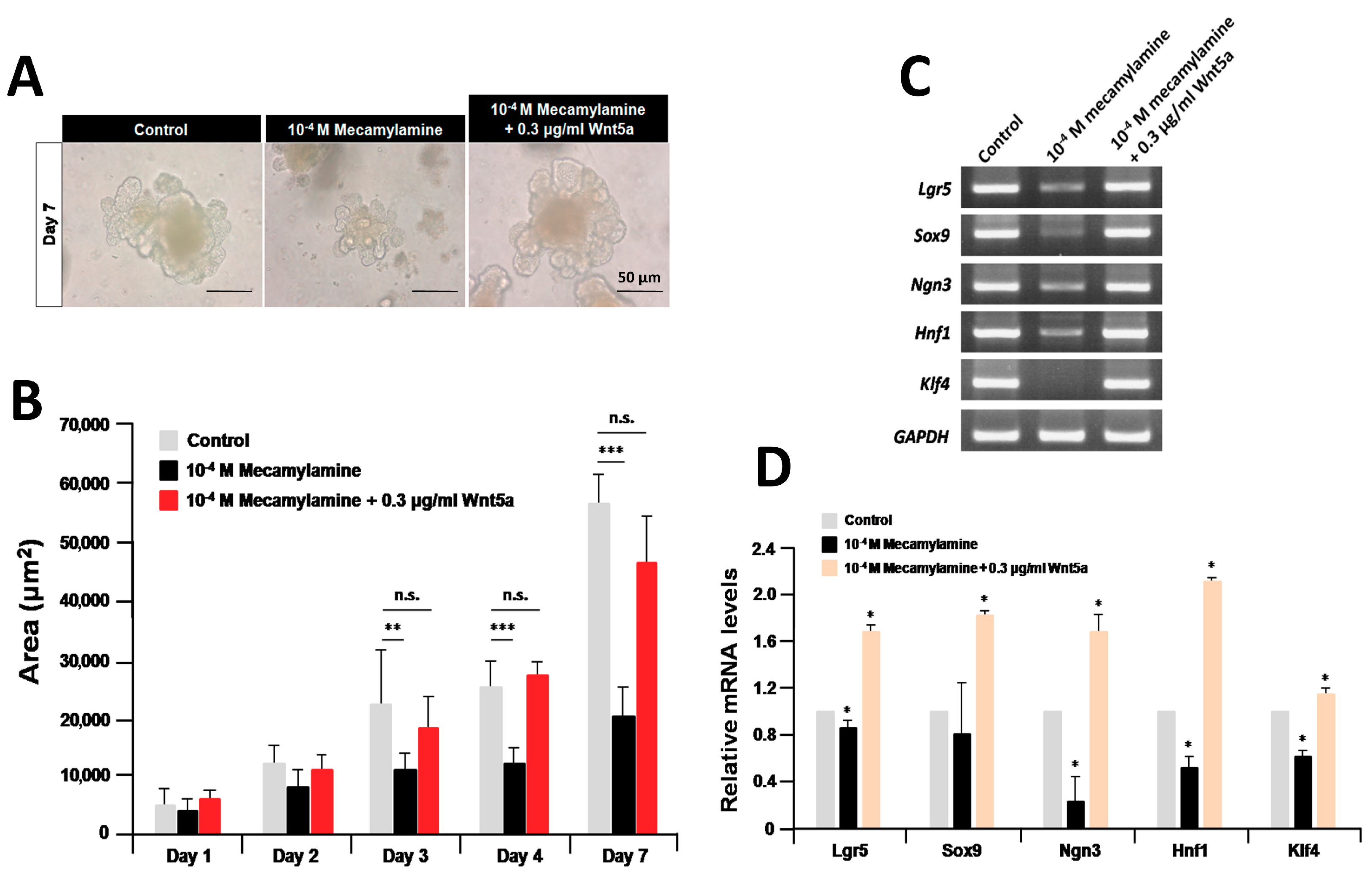
2.5. Recombinant Wnt5a Rescues the Organoids Treated with Mecamylamine
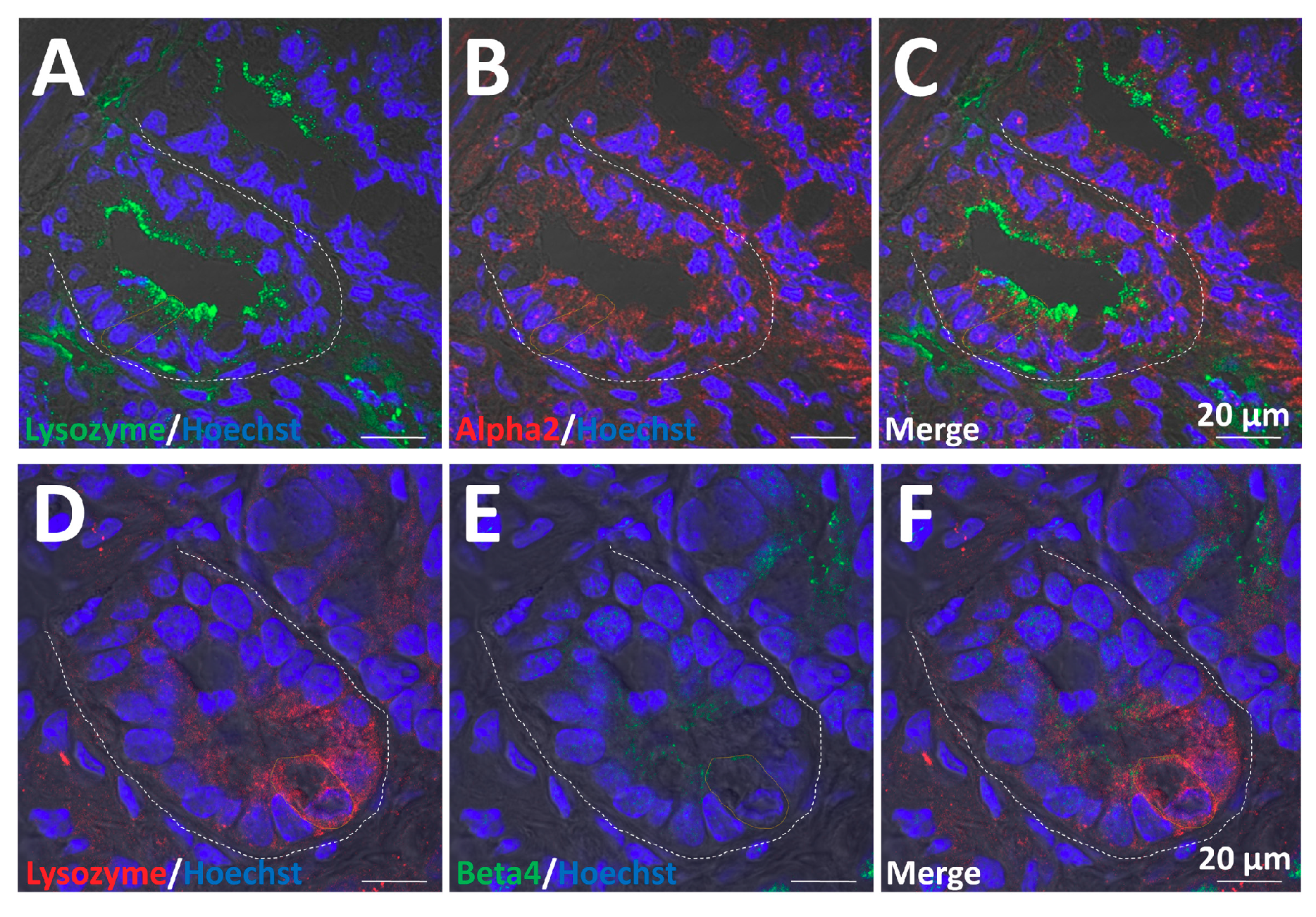
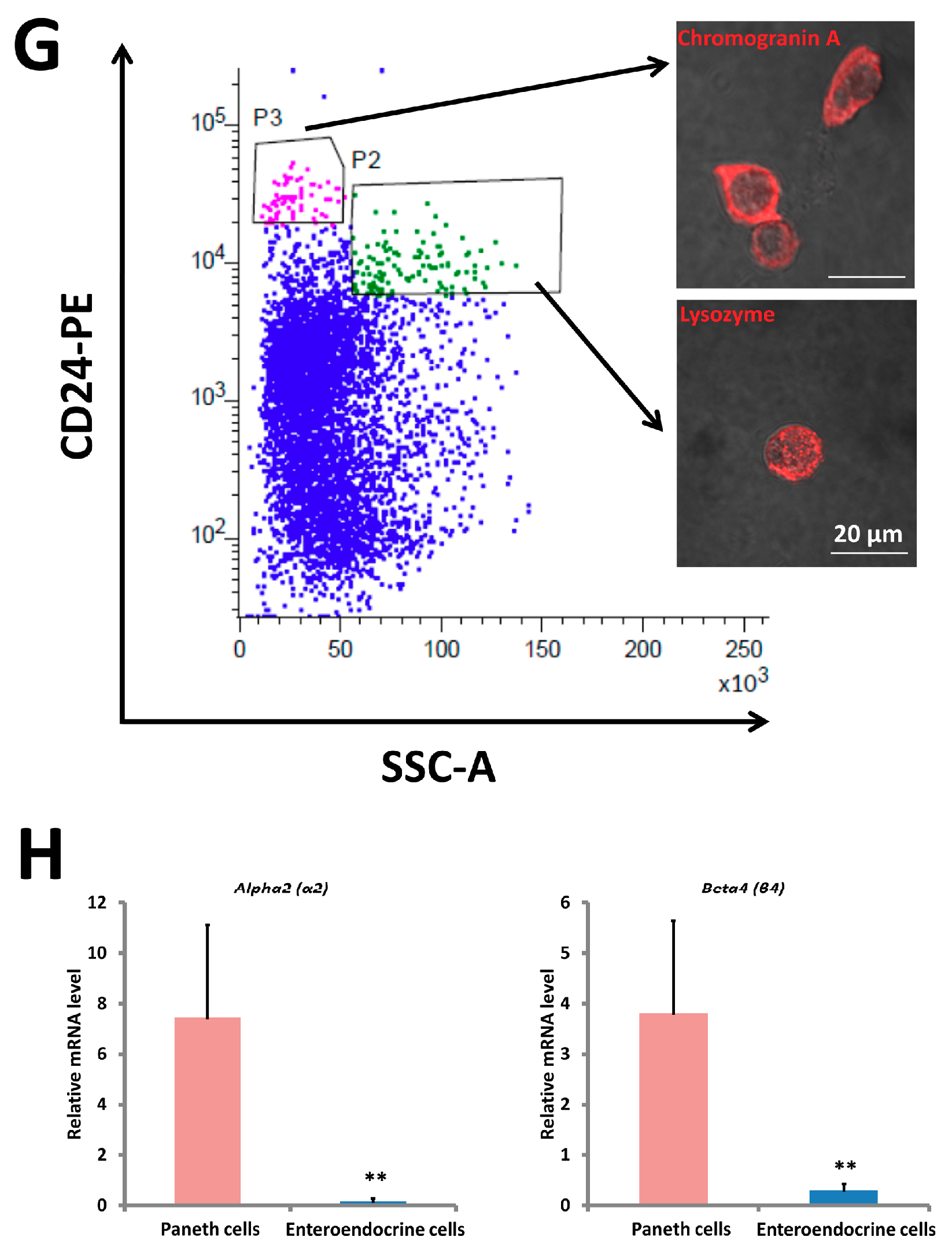
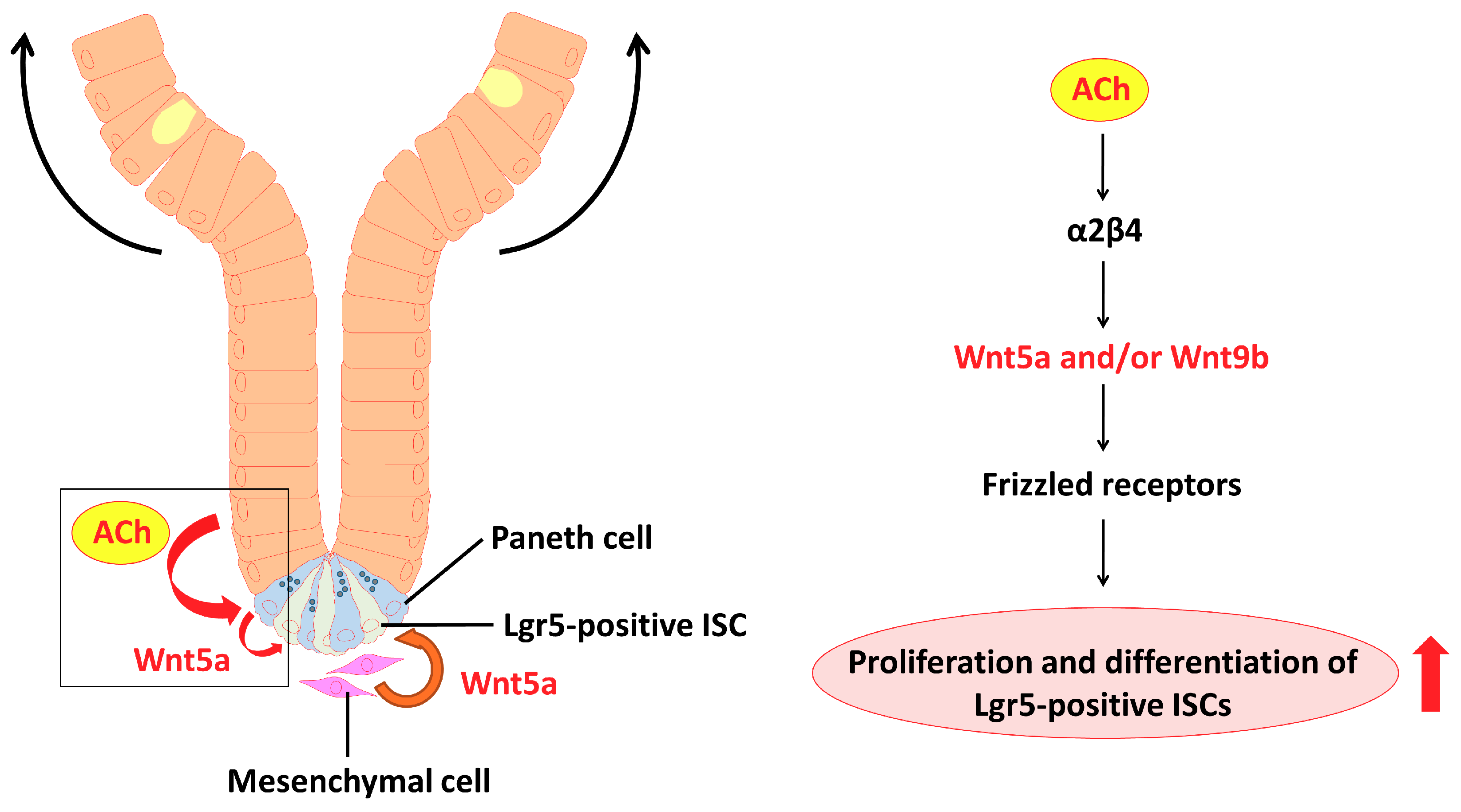
2.6. α2/β4 nAChR Is Localized in Paneth Cells
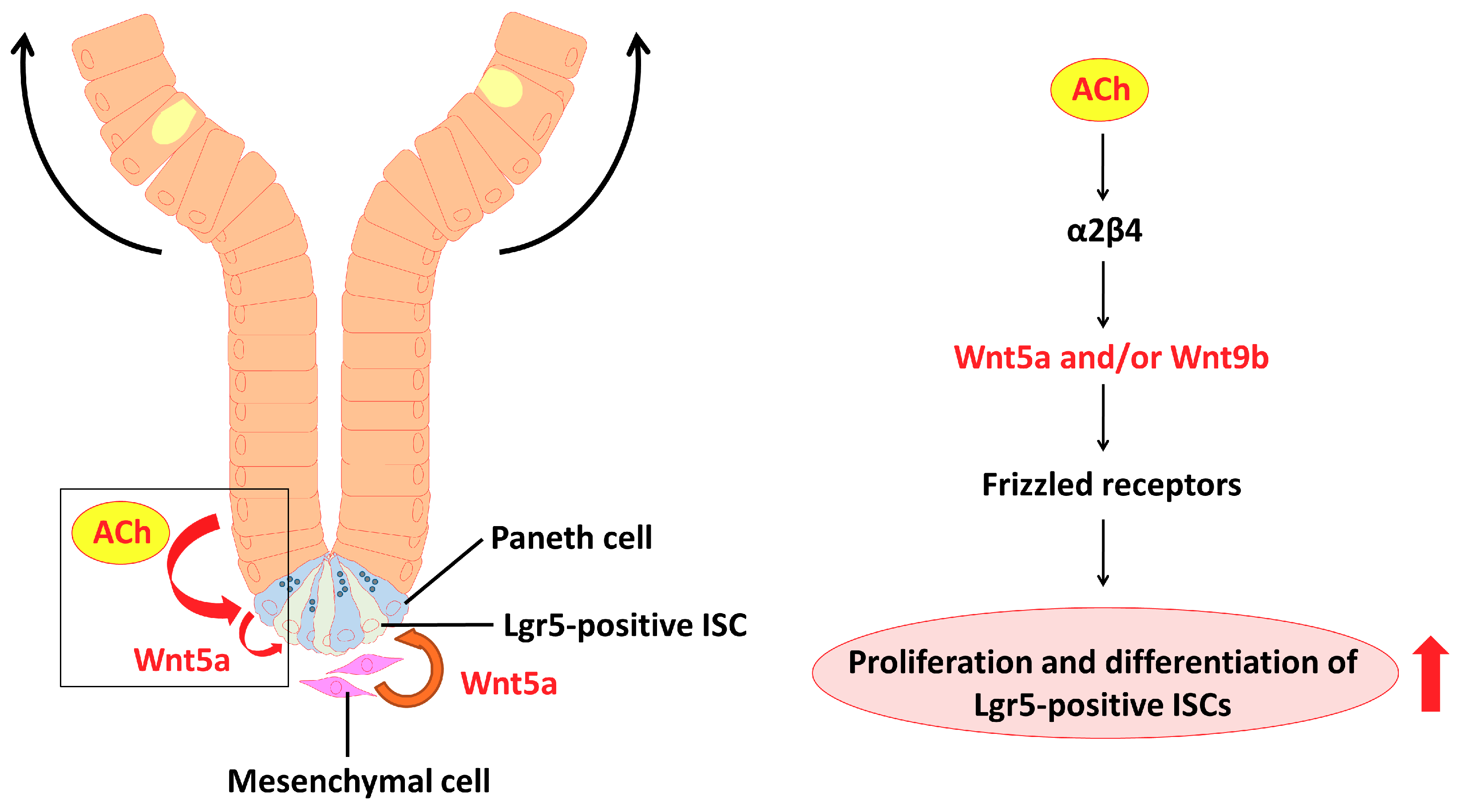
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Crypt Isolation and Crypt–Villus Organoid Culture
4.3. Subculture of Crypt–Villus Organoids
4.4. Pharmacological Assay
4.5. RT-PCR
4.6. Quantitative RT-PCR Analysis
4.7. Immunohistochemistry
4.8. RNA Sequencing
4.9. Flow Cytometry Analysis and Cell Sorting
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Clevers, H. The intestinal crypt, a prototype stem cell compartment. Cell 2013, 154, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- De Lau, W.; Barker, N.; Low, T.Y.; Koo, B.-K.; Li, V.S.; Teunissen, H.; Kujala, P.; Haegebarth, A.; Peters, P.J.; van de Wetering, M.; et al. Lgr5 homologues associate with Wnt receptors and mediate R-spondin signaling. Nature 2011, 476, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vris, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells bulid crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Grando, S.A.; Pitterlkow, M.R.; Schallreuter, K.U. Adrenergic and cholinergic control in the biology of epidermis: Physiological and clinical significance. J. Investig. Dermatol. 2006, 126, 1948–1965. [Google Scholar] [CrossRef] [PubMed]
- Morris, D. The cholinergic acetyltransferase of human placenta. Biochem. J. 1966, 98, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans. Br. J. Pharmacol. 2008, 154, 1558–1571. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.; Kilbinger, H.; Bittinger, F.; Kirkpatrick, C.J. The biological role of non-neuronal acetylcholine in plants and humans. Jpn. J. Pharmacol. 2001, 85, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.; Reinheimer, T.; Kilbinger, F.; Bittinger, F.; Kirkpatrick, C.J.; Saloga, J. Increased acetylcholine levels in skin biopsies of patients with atopic dermatitis. Life Sci. 2003, 72, 2169–2172. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J.; Racke, K. The cholinergic ‘pitfall’: Acetylcholine, a universal cell molecule in biological systems, including humans. Clin. Exp. Pharmacol. Physiol. 1999, 26, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.R. Wnt signaling in the mouse intestine. Oncogene 2006, 25, 7512–7521. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Pinto, D.; Clevers, H. Wnt control of stem cells and differentiation in the intestinal epithelium. Exp. Cell Res. 2005, 306, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, S.; Yamaguchi, T.P.; Hebrok, M. Wnt5a is essential for intestinal elongation in mice. Dev. Biol. 2009, 326, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Korinek, V.; Barker, N.; Moerer, P.; van Donselaar, E.; Huls, G.; Peters, P.J.; Clevers, H. Depletion of epithelial stem-cell compartments in the small intestine of mice acking Tcf-4. Nat. Genet. 1998, 19, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Zhao, J.; Andarmani, S.; Kakitani, M.; Oshima, T.; Binnerts, M.E.; Abo, A.; Tomizuka, K.; Funk, W.D. R-spondin proteins: A novel link to beta-catenin activation. Cell Cycle 2006, 5, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Farin, H.F.; Van Es, J.H.; Clevers, H. Redundant sources of Wnt regulate intestinal stem cells and promote formation of Paneth cells. Gastroenterology 2012, 143, 1518–1529.e7. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.P.; Bradley, A.; McMahon, A.P.; Jones, S. A Wnt5a pathway underlies outgrowth of multiple structures in the vertebrate embryo. Development 1999, 126, 1211–1223. [Google Scholar] [PubMed]
- Lickert, H.; kispert, A.; Kutsch, S.; Kemler, R. Expression pattern of Wnt genes in mouse gut development. Mech. Dev. 2001, 105, 181–184. [Google Scholar] [CrossRef]
- Miyoshi, H.; Ajima, R.; Luo, C.T.; Yamaguchi, T.P.; Stappenbeck, T.S. Wnt5a potentiates TGF-β signaling to promote colonic crypt regeneration after tissue injury. Science 2012, 338, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Ohnishi, H.; Sugiura, Y.; Honda, K.; Suematsu, M.; Kawasaki, T.; Deguchi, T.; Fujii, T.; Orihashi, K.; Hippo, Y.; et al. Non-neuronal acetylcholine as an endogenous regulator of proliferation and differentiation of Lgr5-positive stem cells in mice. FEBS J. 2014, 281, 4672–4690. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.X.; Pereira, E.F.R.; Alkondon, M.; Rogers, S.W. Mammalian nicotinic acetylcholine receptors: From structure to function. Physiol. Rev. 2009, 89, 73–120. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Daugherty, S.; de Groat, W.C. Effects of nicotine receptor agonists on bladder afferent nerve activity in an in vitro bladder-pelvic nerve preparation. Brain Res. 2016, 1637, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, D.; Barth, M.; Layer, P.G.; Sperling, L.E. Acetylcholine as a possible signaling molecule in embryonic stem cells: Studies on survival, proliferation and death. Chem. Biol. Interact. 2010, 187, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, T.; Ozawa, A.; Goshima, H.; Watanabe, Y. Involvement of nicotinic acetylcholine receptor in the proliferation of mouse induced plurpotent stem cells. Life Sci. 2012, 90, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Dwyer, J.B.; Mangold, J.E.; Wang, J.; Wei, J.; Leslie, F.M.; Li, M.D. Modulation of cell adhesion systems by prenatal nicotine exposure in limbic brain regions of adolescent female rats. Int. J. Neuropsychopharmacol. 2011, 14, 157–174. [Google Scholar] [CrossRef] [PubMed]
- Dennis, G.J.; Sherman, B.T.; Hosack, D.A.; Yang, J.; Gao, W.; Lane, H.C.; Lempicki, R.A. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Saint-Jeannet, J.P.; Wang, Y.; Nathans, J.; Dawid, I.; Varmus, H. A member of the Frizzled protein family mediating axis induction by Wnt-5A. Science 1997, 275, 1652–1654. [Google Scholar] [CrossRef] [PubMed]
- Mikels, A.J.; Nusse, R. Purified Wnt5a protein activates or inhibits beta-catenin-TCF signaling depending on receptor context. PLoS Biol. 2006, 4, e115. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; ten Berge, D.; Brown, J.; Ahn, S.; Hu, L.A.; Miller, W.E.; Caron, M.G.; Barak, L.S.; Nusse, R.; Lefkowitz, R.J. Dishevelled 2 recruits beta-arrestin 2 to mediate Wnt5a-stimulated endocytosis of Frizzled 4. Science 2003, 301, 1391–1394. [Google Scholar] [CrossRef] [PubMed]
- Dejmek, J.; Sӓfholm, A.; Nielsen, C.K.; Andersson, T.; Leandersson, K. Wnt-5a/Ca2+-induced NFAT activity is counteracted by Wnt-5a/Yes-Cdc42-casein kinase 1α signaling in human mammary epithelial cells. Mol. Cell. Biol. 2006, 26, 6024–6036. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Dodge, M.E.; Tang, W.; Lu, J.; Ma, Z.; Fan, C.W.; Wei, S.; Hao, W.; Kilgore, J.; Williams, N.S.; et al. Small molecule-mediated disruption of Wnt-dependent signaling in tissue regeneration and cancer. Nat. Chem. Biol. 2009, 5, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Conti-Tronconi, B.M.; McLane, K.E.; Raftery, M.A.; Grando, S.; Protti, M.P. The nicotinic acetylcholine receptor: Structure and auto immune pathology. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 69–123. [Google Scholar] [CrossRef] [PubMed]
- Changeux, J.P. Thudichum medal lecture. The acetylcholine receptor: A model for allosteric membrane proteins. Biochem. Soc. Trans. 1995, 23, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, J.M. Nicotinic acetylcholine receptors. In Handbook of Receptors and Channels: Ligand-and Voltage-Gated Ion Channels; North, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 153–175. [Google Scholar]
- Albuquerque, E.X.; Alkondon, M.; Pereira, E.F.; Castro, N.G.; Schrattenholz, A.; Barbosa, C.T.; Bonfante-Cabarcas, R.; Aracava, Y.; Eisenberg, H.M.; Maelicke, A. Properties of neuronal nicotinic acetylcholine receptors: Pharmacological characterization and modulation of synaptic function. J. Pharmacol. Exp. Ther. 1997, 280, 1117–1136. [Google Scholar] [PubMed]
- Grando, S.A.; Horton, R.M.; Pereira, E.F.; Diethelm-Okita, B.M.; Gorge, P.M.; Albuquerque, E.X.; Conti-Fine, B.M. A nicotinic acetylcholine receptor regulating cell adhesion and motility is expressed in human keratinocytes. J. Investig. Dermatol. 1995, 105, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Conti-Fine, B.M.; Navaneetham, D.; Lei, S.; Maus, A.D. Neuronal nicotinic receptors in non-neuronal cells: New mediators of tobacco toxicity? Eur. J. Pharmacol. 2000, 393, 279–294. [Google Scholar] [CrossRef]
- Wessler, I.; Kirkpatrick, C.J.; Racké, K. Non-neuronal acetylcholine, a locally acting molecule, widely distributed in biological systems: Expression and function in humans. Pharmacol. Ther. 1998, 77, 59–79. [Google Scholar] [CrossRef]
- Macklin, K.D.; Maus, A.D.J.; Pereira, E.F.R.; Albuquerque, E.X.; Conti-Fine, B.M. Human vascular endothelial cells express functional nicotinic acetylcholine receptors. J. Pharmacol. Exp. Ther. 1998, 287, 435–439. [Google Scholar] [PubMed]
- Veeman, M.T.; Axelrod, J.D.; Moon, R.T. A second canon. Functions and mechanisms of β-catenin-independent Wnt signaling. Dev. Cell 2003, 5, 367–377. [Google Scholar] [CrossRef]
- Kohn, A.D.; Moon, R.T. Wnt and calcium signaling: β-catenin-independent pathways. Cell Calcium 2005, 38, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The Wnt/Ca2+ pathway: A new vertebrate Wnt signaling pathway takes shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Sheldahl, L.C.; Slusarski, D.C.; Pandur, P.; Miller, J.R.; Kuhl, M.; Moon, R.T. Dishevelled activates Ca2+ flux, PKC, and CamKII in vertebrate embryos. J. Cell Biol. 2003, 161, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, F.M.P.; Santos, A.K.; Gomes, D.A.; da Silva, S.L.; Gomes, K.N.; Ladeira, L.O.; Resende, R.R. Stem cells and calcium signaling. Adv. Exp. Med. Biol. 2012, 740, 891–916. [Google Scholar] [PubMed]
- Crabtree, G.R.; Olson, E.N. NFAT signaling: Choreographing the social lives of cells. Cell 2002, 109 (Suppl. 1), S67–S79. [Google Scholar] [CrossRef]
- Rao, A.; Luo, C.; Hogan, P.G. Transcription factors of the NFAT family: Regulation and function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef] [PubMed]
- Mo, J.S.; Park, H.W.; Guan, K.L. The Hippo signaling pathway in stem cell biology and cancer. EMBO Rep. 2014, 15, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Bakker, E.R.M.; Raghoebir, L.; Franken, P.F.; Helvensteijn, W.; van Gurp, L.; Meijlink, F.; van der Valk, M.A.; Rottier, R.J.; Kuipers, E.J.; van Veelen, W.; et al. Induced Wnt5a expression perturbs embryonic outgrowth and intestinal elongation, but is well-tolerated in adult mice. Dev. Biol. 2012, 369, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.W.; Yeh, J.C.; Fan, T.P.; Smith, S.K.; Charnock-Jones, D.S. Wnt5a-mediated non-canonical Wnt signaling regulates human endothelial cell proliferation and migration. Biochem. Biophys. Res. Commun. 2008, 365, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, A.; Yamamoto, H.; Sato, A.; Matsumoto, S. Wnt5a: Its signaling, functions and implication in disease. Acta. Physiol. (Oxf.) 2012, 204, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Nishita, M.; Enomoto, M.; Yamagata, K.; Minami, Y. Cell/tissue-tropic functions of Wnt5a signaling in normal and cancer cells. Trends Cell Biol. 2010, 20, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Pukrop, T.; Binder, C. The complex pathways of Wnt5a in cancer progression. J. Mol. Med. 2008, 86, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Gregorieff, A.; Pinto, D.; Begthel, H.; Destree, O.; Kielman, M.; Clevers, H. Expression pattern of Wnt signaling components in the adult intestine. Gastroenterology 2005, 129, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Udagawa, J.; Matsumoto, A.; Hashimoto, R.; Hatta, T.; Nishita, M.; Minami, Y.; Otani, H. Ror2 is required for midgut elongation during mouse development. Dev. Dyn. 2010, 239, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Bakker, E.R.; Das, A.M.; Helvensteijn, W.; Franken, P.F.; Swagemakers, S.; van der Valk, M.A.; ten Hagen, T.L.; Kuipers, E.J.; van Veelen, W.; Smits, R. Wnt5a promotes human colon cancer cell migration and invasion but does not augment intestinal tumorigenesis in Apc1638N mice. Carcinogenesis 2013, 34, 2629–2638. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Sun, B.; Liu, Z.; Zhao, X.; Qi, L.; Li, Y.; Gu, Q. Wnt5a suppresses colon cancer by inhibiting cell proliferation and epithelial-mesenchymal transition. J. Cell. Physiol. 2014, 229, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Posadas, I.; Lópes-Hernández, B.; Ceňa, V. Nicotinic receptors in neurodegeneration. Curr. Neuropharmacol. 2013, 11, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Eglen, R.M.; Randle, D.H. Drug discovery goes three-dimensional: Goodbye to flat high-throughput screening? Assay Drug Dev. Technol. 2015, 13, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Martin, U. Pluripotent stem cells for disease modeling and drug screening: New perspectives for treatment of cystic fibrosis? Mol. Cell. Pediatr. 2015, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Sampaziotis, F.; Cardoso De Brito, M.; Madrigal, P.; Bertero, A.; Saeb-Parsy, K.; Soares, F.A.C.; Schrumpf, E.; Melum, E.; Karlsen, T.H.; Bradley, J.A.; et al. Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation. Nat. Biotechnol. 2015, 33, 845–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedhia, P.H.; Bertaux-Skeirik, N.; Zavros, Y.; Spence, J.R. Organoid models of human gastrointestinal development and disease. Gastroenterology 2016, 150, 1098–1112. [Google Scholar] [CrossRef] [PubMed]
- Hynds, R.E.; Giangreco, A. Concise review: The relevance of human stem cell-derived organoids models for epithelial translational medicine. Stem Cells 2013, 31, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Walsh, A.J.; Cook, R.S.; Sanders, M.E.; Arteaga, C.L.; Skala, M.C. Drug response in organoids generated from frozen primary tumor tissues. Sci. Rep. 2016, 6, 18889. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway ID | KEGG Pathway Designation | Number of Genes | Fold Enrichment |
|---|---|---|---|
| mmu04550 | Signaling pathways regulating pluripotency of stem cells | 26 | 2.88 |
| mmu04919 | Thyroid hormone signaling pathway | 21 | 2.84 |
| mmu04390 | Hippo signaling pathway | 21 | 2.13 |
| mmu04310 | Wnt signaling pathway | 19 | 2.06 |
| mmu04010 | MAPK signaling pathway | 27 | 1.63 |
| mmu04144 | Endocytosis | 29 | 1.59 |
| mmu04015 | Rap1 signaling pathway | 25 | 1.79 |
| mmu04810 | Regulation of actin cytoskeleton | 22 | 1.57 |
| Accession | Gene Name | Nicotine (Fold Change *) | Mecamylamine (Fold Change *) | |||
|---|---|---|---|---|---|---|
| 10−7 M(3 Day) | 10−5 M(3 Day) | 10−7 M(7 Day) | 10−5 M(7 Day) | 10−6 M (7 Days) | ||
| NM_019827 | Gsk3β | 1.09 | 1.05 | 1.64 | 1.71 | 1.00 |
| NM_007614 | Ctnnb1 | 0.91 | 0.78 | 1.61 | 1.88 | 0.84 |
| NM_007561 | Bmpr2 | 1.01 | 1.06 | 1.60 | 1.70 | 0.98 |
| NM_007889 | Dvl3 | 1.08 | 1.02 | 1.74 | 1.73 | 1.10 |
| NM_001005784 | Patj | 0.39 | 0.62 | 3.74 | 3.88 | 1.06 |
| NM_026735 | Mob1b | 1.15 | 1.10 | 2.27 | 2.37 | 0.83 |
| NM_007862 | Dlg1 | 1.19 | 1.18 | 1.74 | 1.96 | 1.66 |
| NM_001109752 | Dlg4 | 16.37 | 7.20 | 2.28 | 2.62 | 0.80 |
| NM_145571 | Mob1a | 1.04 | 1.05 | 1.64 | 1.84 | 0.96 |
| NM_001166585 | Tead1 | 1.65 | 1.30 | 1.95 | 1.64 | 1.13 |
| NM_010690 | Lats1 | 1.10 | 1.08 | 1.64 | 1.62 | 0.98 |
| NM_001164112 | Nfatc1 | 1.20 | 0.87 | 1.60 | 1.99 | 0.00 |
| NM_009521 | Wnt3 | 1.10 | 1.13 | 1.10 | 1.03 | 1.05 |
| NM_001256224 | Wnt5a | 1.30 | 1.37 | 4624.5 | 3885.7 | 0.00 |
| NM_011719 | Wnt9b | 1.30 | 1.25 | 1.84 | 1.41 | 0.29 |
| NM_020510 | Frizzled-2 | 1.16 | 1.14 | 1.11 | 0.95 | 0.89 |
| NM_008055 | Frizzled-4 | 0.98 | 0.94 | 1.82 | 1.99 | 0.83 |
| NM_022721 | Frizzled-5 | 1.05 | 0.89 | 1.84 | 1.80 | 0.97 |
| NM_001162494 | Frizzled-6 | 1.02 | 1.08 | 2.01 | 1.88 | 1.02 |
| NM_010195 | Lgr5 | 1.16 | 1.09 | 1.46 | 1.78 | 0.53 |
| NM_011448 | Sox9 | 1.07 | 1.33 | 1.25 | 1.48 | 0.27 |
| NM_009719 | Ngn3 | 1.30 | 1.59 | 0.97 | 1.39 | 0.28 |
| NM_009327 | Hnf1 | 1.10 | 1.17 | 1.19 | 1.31 | 0.15 |
| NM_010637 | Klf4 | 1.03 | 1.04 | 1.31 | 1.44 | 0.17 |
| KEGG Pathway ID | KEGG Pathway Designation | Number of Genes | Fold Enrichment |
|---|---|---|---|
| mmu04550 | Signaling pathways regulating pluripotency of stem cells | 16 | 2.20 |
| mmu04919 | Thyroid hormone signaling pathway | 9 | 1.50 |
| mmu04390 | Hippo signaling pathway | 14 | 1.70 |
| mmu04310 | Wnt signaling pathway | 13 | 1.70 |
| mmu04010 | MAPK signaling pathway | 25 | 1.80 |
| mmu04144 | Endocytosis | 27 | 1.80 |
| mmu04015 | Rap1 signaling pathway | 20 | 1.70 |
| mmu04810 | Regulation of actin cytoskeleton | 20 | 1.70 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, T.; Shiraishi, A.; Murata, J. The Coordinated Activities of nAChR and Wnt Signaling Regulate Intestinal Stem Cell Function in Mice. Int. J. Mol. Sci. 2018, 19, 738. https://doi.org/10.3390/ijms19030738
Takahashi T, Shiraishi A, Murata J. The Coordinated Activities of nAChR and Wnt Signaling Regulate Intestinal Stem Cell Function in Mice. International Journal of Molecular Sciences. 2018; 19(3):738. https://doi.org/10.3390/ijms19030738
Chicago/Turabian StyleTakahashi, Toshio, Akira Shiraishi, and Jun Murata. 2018. "The Coordinated Activities of nAChR and Wnt Signaling Regulate Intestinal Stem Cell Function in Mice" International Journal of Molecular Sciences 19, no. 3: 738. https://doi.org/10.3390/ijms19030738