Notched Belly Grain 4, a Novel Allele of Dwarf 11, Regulates Grain Shape and Seed Germination in Rice (Oryza sativa L.)


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
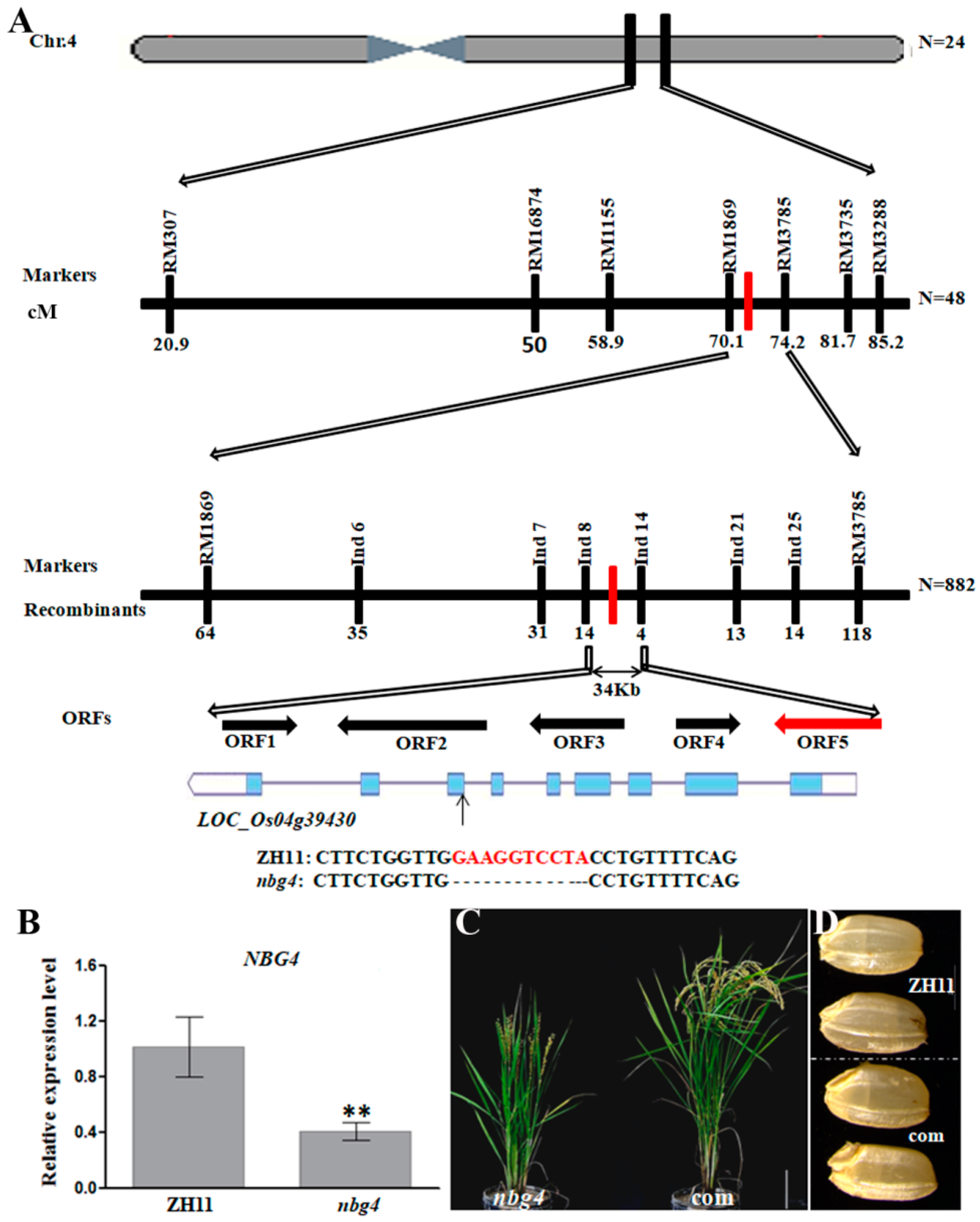
2.1. A31 Is an NBG Mutant
2.2. Map-Based Cloning of NBG4
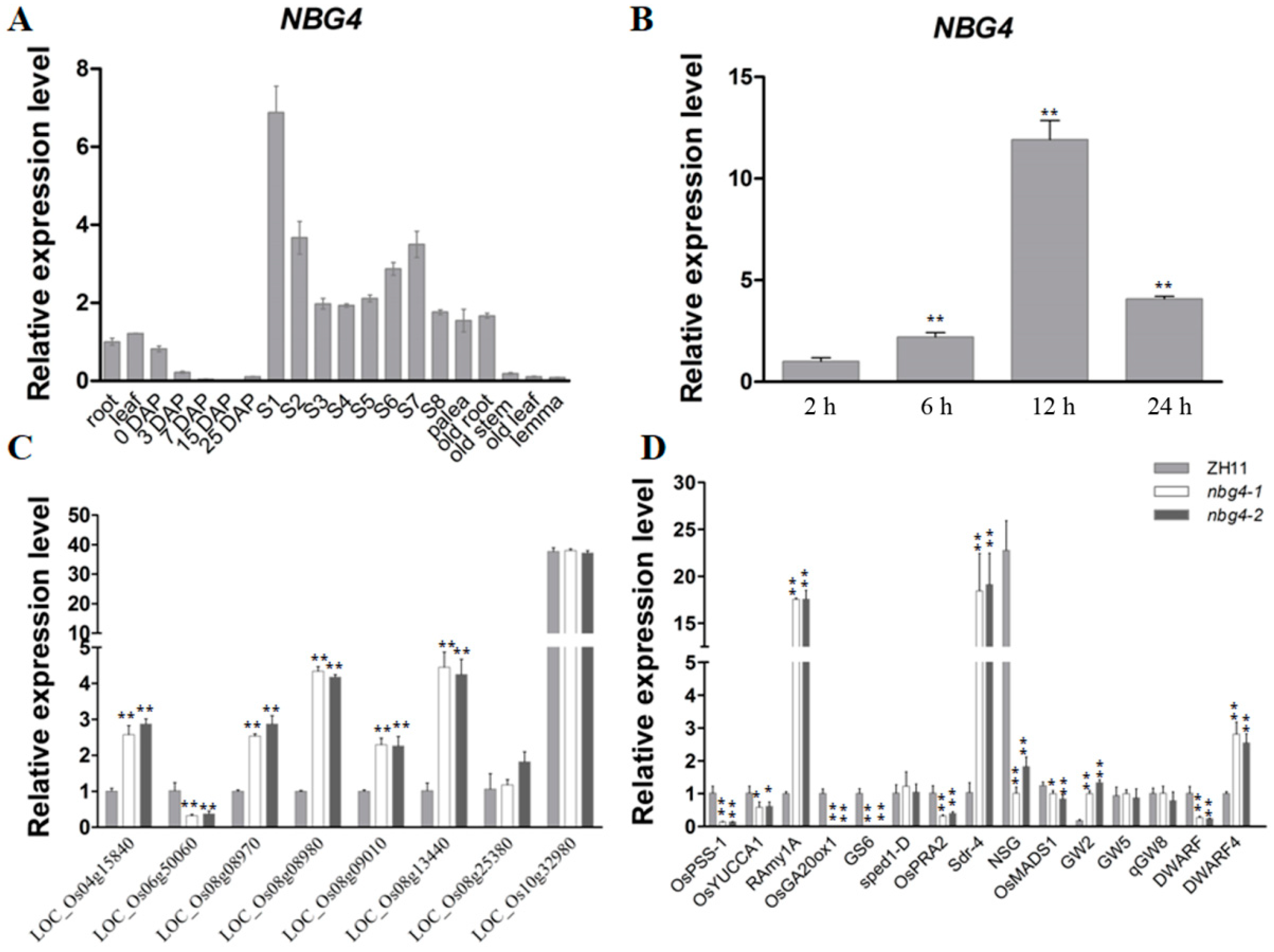
2.3. Downstream Genes Regulated by NBG4
3. Materials and Methods
3.1. Plant Materials and Growth Conditions
3.2. Map-Based Cloning, Plasmid Construction, and Plant Transformation
3.3. RNA Preparation and RT-PCR Analysis
3.4. RNA-seq Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Guo, D.; Chang, Y.; You, C.; Li, X.; Dai, X.; Weng, Q.; Zhang, J.; Chen, G.; Li, X.; et al. Non-random distribution of T-DNA insertions at various levels of the genome hierarchy as revealed by analyzing 13 804 T-DNA flanking sequences from an enhancer-trap mutant library. Plant J. Cell Mol. Boil. 2007, 49, 947–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Xu, R.; Duan, P.; Li, Y. Control of grain size in rice. Plant Reprod. 2018, 31, 237–251. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Li, J. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Annu. Rev. Genet. 2014, 48, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.R.; Yin, L.L.; Xue, H.W. Functional genomics based understanding of rice endosperm development. Curr. Opin. Plant Boil. 2013, 16, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Sobrizal, Y.A. Identification of a notched kernel gene associated with pre-harvest sprouting using oryza glumaepatula introgression lines in rice. Ipb 2007, 35. [Google Scholar] [CrossRef]
- Takeda, K.; Takahashi, M.E. Unbalanced growth in floral glumes and caryopsis in rice. I. varietal difference in the degree of unbalance and the occurrence of malformed grains. Jpn. J. Breed 1970, 20, 337–343. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, X.; Yang, X.; Li, G.; Tang, S.; Wang, S.; Ding, Y.; Liu, Z. Proteomic analysis of proteins related to rice grain chalkiness using iTRAQ and a novel comparison system based on a notched-belly mutant with white-belly. BMC Plant Boil. 2014, 14, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Zhang, X.; Wang, Z.; Jiang, Y.; Liu, Z.; Alexander, D.; Li, G.; Wang, S.; Ding, Y. Metabolomic analysis of pathways related to rice grain chalkiness by a notched-belly mutant with high occurrence of white-belly grains. BMC Plant Boil. 2017, 17, 39. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Z.; Min, S.; Kong, F.; Zhu, X. Genetic analysis of notched grain in rice. Chin. J. Rice Sci. 1986, 952. [Google Scholar] [CrossRef]
- Nagato, K.; Kobayashi, Y. Studies on the Occurence of Notched-Belly (Dogire-mai) in Rice Plants. Jpn. J. Crop. Sci. 2008, 26, 13–14. [Google Scholar] [CrossRef]
- Takeda, K. Temperature Response of Caryopsis Elongation in Relation to Grain Notching of Rice. Jpn. J. Crop. Sci. 1985, 54, 253–260. [Google Scholar] [CrossRef]
- Goto, E.; Kumagai, S. The effect of temperature and shading on notched-belly rice kernel in Hokkaido rice variety “Nanatsuboshi”. Jpn. J. Crop. Sci. 2009, 78, 35–42. [Google Scholar] [CrossRef]
- Pavithran, K. Inheritance and linkage relationship of notched kernel in rice (oryza). J. Can. Génétique Cytol. 1977, 19, 483–486. [Google Scholar] [CrossRef]
- Tanabe, S.; Ashikari, M.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Yano, M.; Yoshimura, A.; Kitano, H.; Matsuoka, M.; Fujisawa, Y. A Novel Cytochrome P450 Is Implicated in Brassinosteroid Biosynthesis via the Characterization of a Rice Dwarf Mutant, dwarf11, with Reduced Seed Length. Plant Cell 2005, 17, 776–790. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Rao, Y.; Xu, J.; Hu, S.; Fang, Y.; Yu, H.; Pan, J.; Liu, R.; Ren, D.; Wang, X. Characterization and cloning of SMALL GRAIN 4, a novel DWARF11 allele that affects brassinosteroid biosynthesis in rice. Sci. Bull. 2015, 60, 905. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, Y.; Zhao, S.; Gu, P.; Zhu, Z.; Sun, C.; Tan, L. Clustered primary branch 1, a new allele of DWARF11, controls panicle architecture and seed size in rice. Plant Biotechnol. J. 2015, 14, 377–386. [Google Scholar] [CrossRef]
- Wang, N.; Li, Y.F.; Sang, X.C.; Ling, Y.H.; Zhao, F.M.; Yang, Z.L.; He, G.H. nonstop glumes (nsg), a novel mutant affects spikelet development in rice. Genes Genom. 2013, 35, 149–157. [Google Scholar] [CrossRef]
- Trusov, Y.; Botella, J.R. New faces in plant innate immunity: Heterotrimeric G proteins. J. Plant Biochem. Biotechnol. 2012, 21, 40–47. [Google Scholar] [CrossRef]
- Song, X.J.; Huang, W.; Shi, M.; Zhu, M.Z.; Lin, H.X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.J.; Yu, X.; Zhou, K. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef]
- Sun, L.; Li, X.; Fu, Y.; Zhu, Z.; Tan, L.; Liu, F.; Sun, X.; Sun, X.; Sun, C. GS6, a member of the GRAS gene family, negatively regulates grain size in rice. J. Integr. Plant Boil. 2013, 55, 938–949. [Google Scholar] [CrossRef]
- Wang, S.; Li, S.; Liu, Q.; Wu, K.; Zhang, J.; Wang, S.; Wang, Y.; Chen, X.; Zhang, Y.; Gao, C. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Sci. Found. China 2015, 47, 949. [Google Scholar] [CrossRef]
- Jiang, G.; Xiang, Y.; Zhao, J.; Yin, D.; Zhao, X.; Zhu, L.; Zhai, W. Regulation of inflorescence branch development in rice through a novel pathway involving the pentatricopeptide repeat protein sped1-D. Genetics 2014, 197, 1395–1407. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, H.; Zhu, K.; Fang, Y.; Gao, Z.; He, Y.; Zhang, G.; Guo, L.; Zeng, D.; Dong, G. Identification and characterization of shortened uppermost internode 1, a gene negatively regulating uppermost internode elongation in rice. Plant Mol. Boil. 2011, 77, 475–487. [Google Scholar]
- Zhang, G.; Song, X.; Guo, H.; Wu, Y.; Chen, X.; Fang, R. A Small G Protein as a Novel Component of the Rice Brassinosteroid Signal Transduction. Mol. Plant 2016, 9, 1260–1271. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin biosynthesis by the YUCCA genes in rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef]
- Mori, M.; Nomura, T.; Ooka, H.; Yokota, T.; Sugimoto, K.; Satoh, K.; Yamamoto, K.; Hirochika, H.; Kikuchi, S. Isolation and Characterization of a Rice Dwarf Mutant with a Defect in Brassinosteroid Biosynthesis. Plant Physiol. 2002, 130, 1152–1161. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Li, S.; Jiao, G.; Sheng, Z.; Wu, Y.; Shao, G.; Xie, L.; Peng, C.; Xu, J.; Tang, S. OsPK2 encodes a plastidic pyruvate kinase involved in rice endosperm starch synthesis, compound granule formation and grain filling. Plant Biotechnol. J. 2018, 16, 1878–1891. [Google Scholar] [CrossRef] [Green Version]
- Hiei, Y.; Komari, T.; Kubo, T. Transformation of rice mediated by Agrobacterium tumefaciens. Plant Mol. Boil. 1997, 35, 205–218. [Google Scholar] [CrossRef]
- Leng, N.; Dawson, J.A.; Thomson, J.A.; Ruotti, V.; Rissman, A.I.; Smits, B.M.; Haag, J.D.; Gould, M.N.; Stewart, R.M.; Kendziorski, C. EBSeq: An empirical Bayes hierarchical model for inference in RNA-seq experiments. Bioinformatics 2013, 29, 1035–1043. [Google Scholar] [CrossRef]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, X.; Wang, Y.; Sun, A.; Bello, B.K.; Ni, S.; Zhang, J. Notched Belly Grain 4, a Novel Allele of Dwarf 11, Regulates Grain Shape and Seed Germination in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2018, 19, 4069. https://doi.org/10.3390/ijms19124069
Tong X, Wang Y, Sun A, Bello BK, Ni S, Zhang J. Notched Belly Grain 4, a Novel Allele of Dwarf 11, Regulates Grain Shape and Seed Germination in Rice (Oryza sativa L.). International Journal of Molecular Sciences. 2018; 19(12):4069. https://doi.org/10.3390/ijms19124069
Chicago/Turabian StyleTong, Xiaohong, Yifeng Wang, Aiqun Sun, Babatunde Kazeem Bello, Shen Ni, and Jian Zhang. 2018. "Notched Belly Grain 4, a Novel Allele of Dwarf 11, Regulates Grain Shape and Seed Germination in Rice (Oryza sativa L.)" International Journal of Molecular Sciences 19, no. 12: 4069. https://doi.org/10.3390/ijms19124069