Hanks-Type Serine/Threonine Protein Kinases and Phosphatases in Bacteria: Roles in Signaling and Adaptation to Various Environments



Abstract
:1. Introduction
2. Structure and Mechanism of Action of Bacterial STKs and STPs
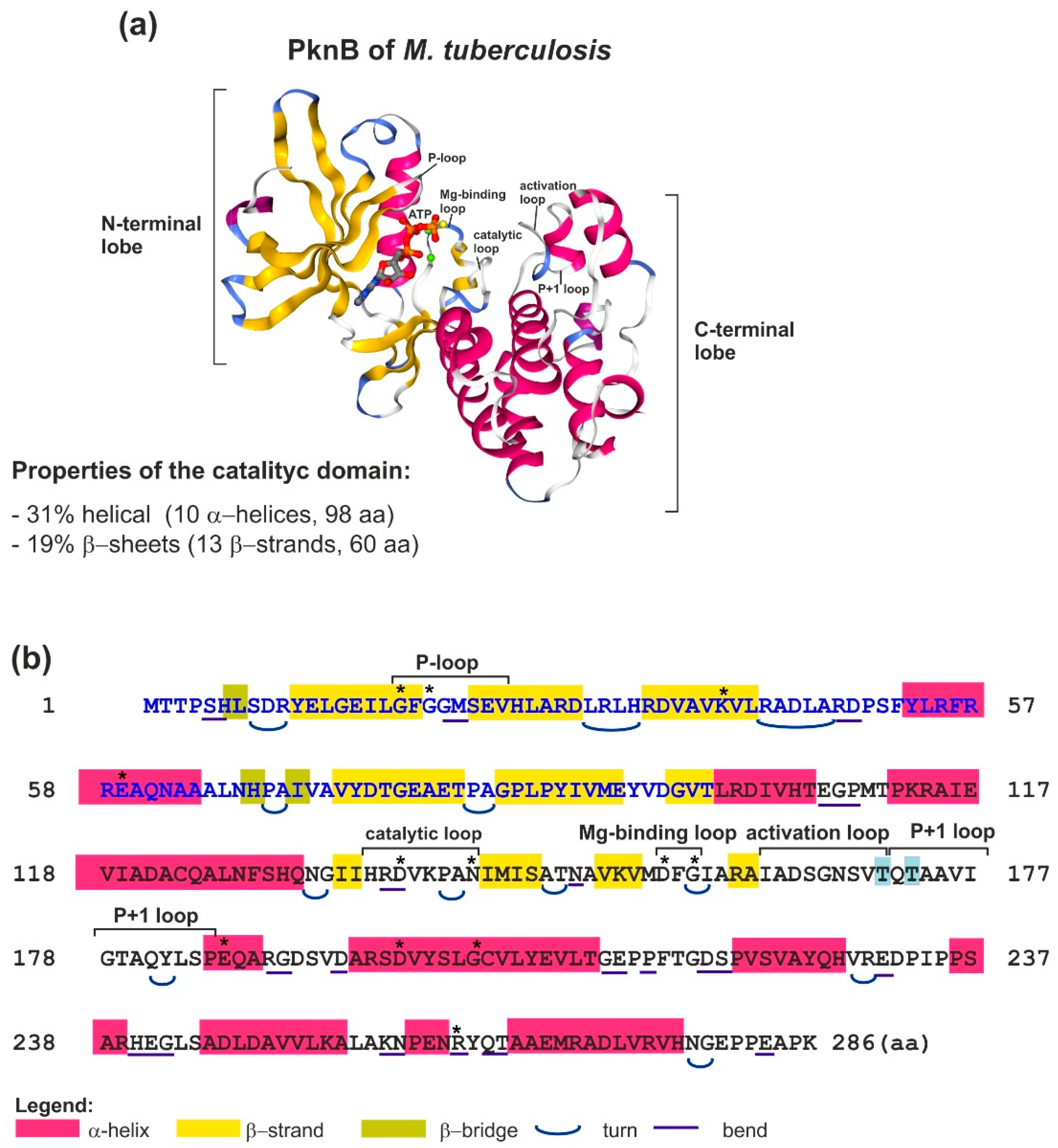
2.1. Structure and Mechanism of Action of Bacterial STKs
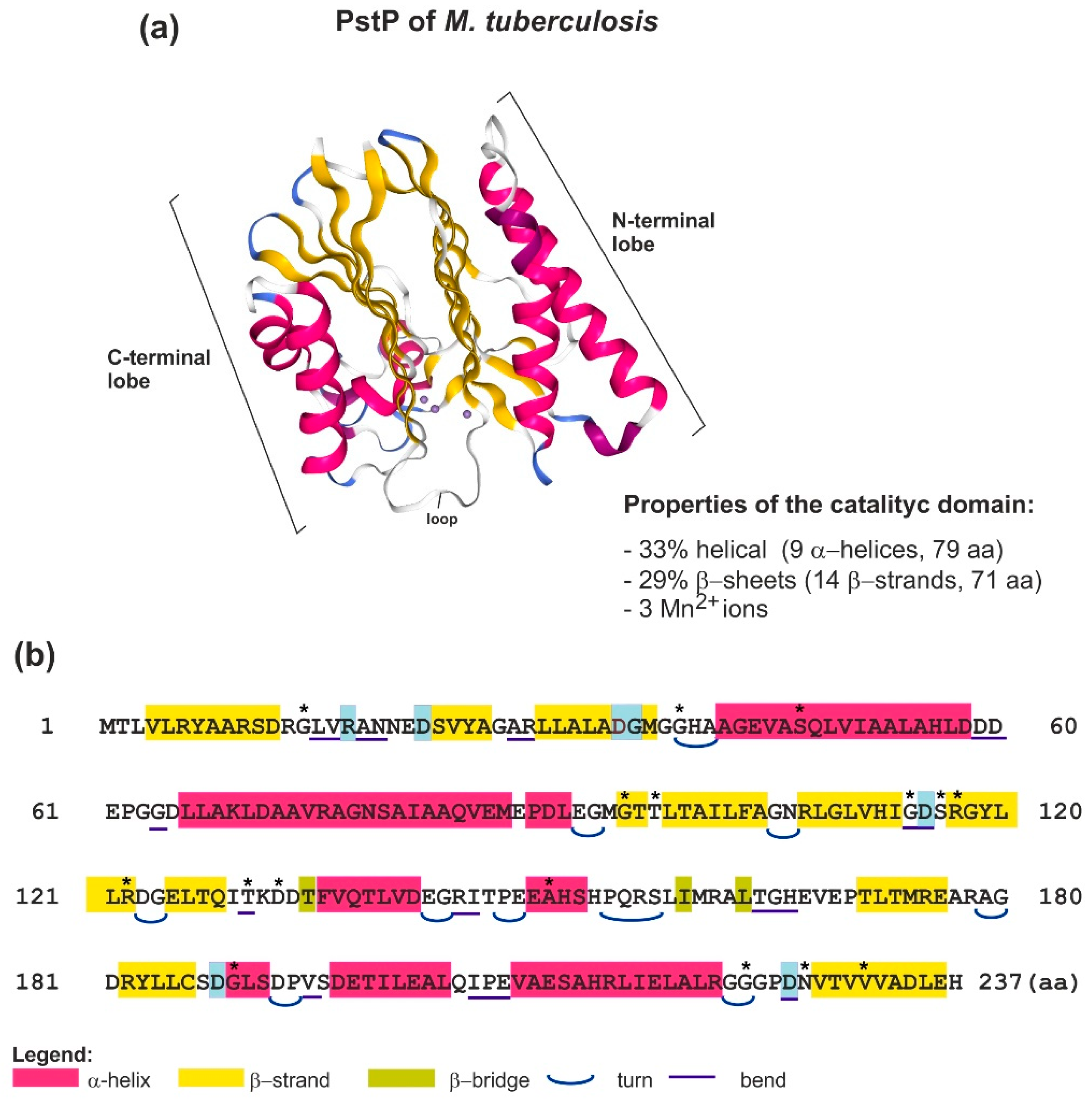
2.2. Structure and Function of Bacterial STPs
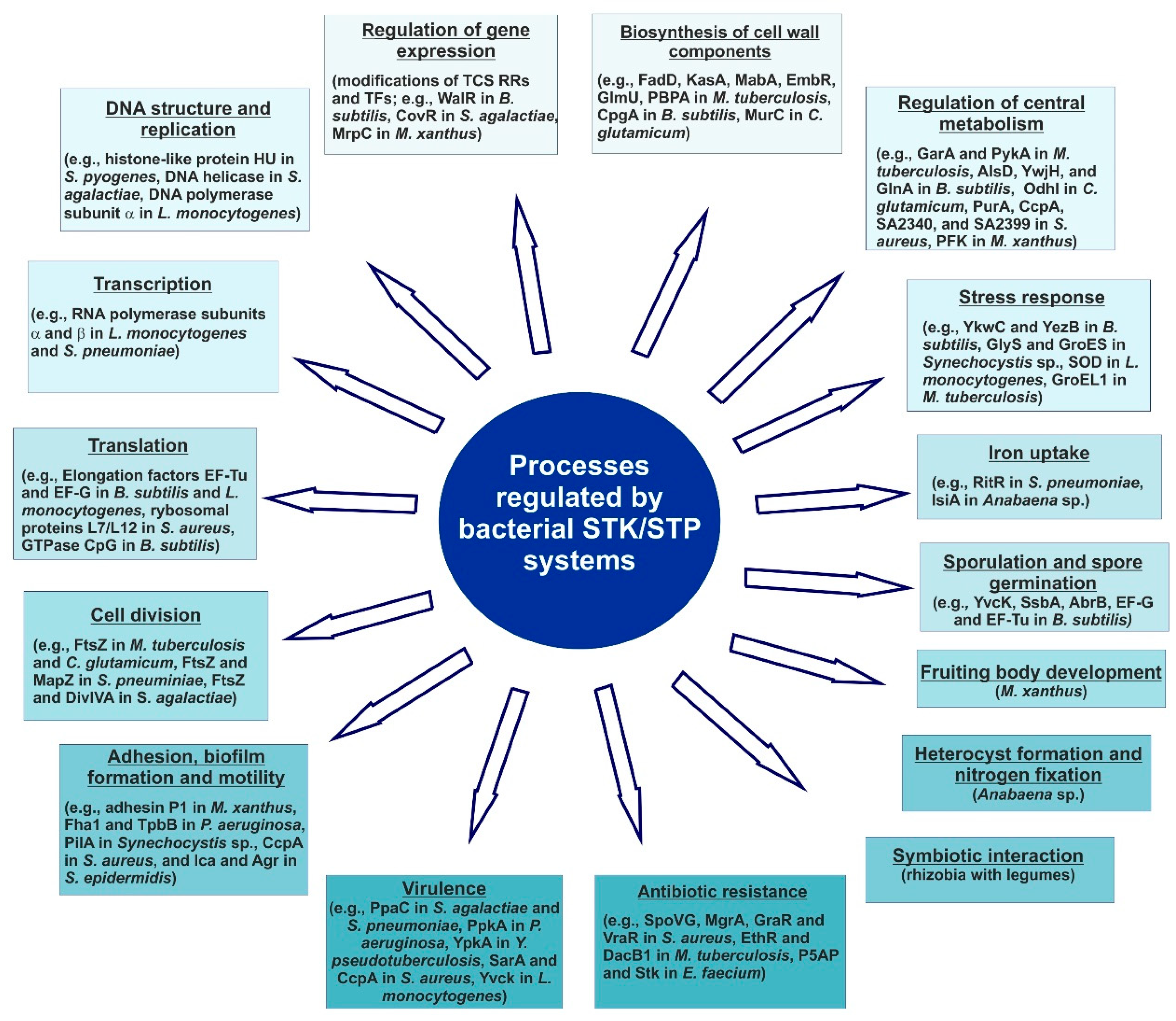
3. The Role of STKs and STPs in Bacterial Signaling and Physiology
3.1. Interactions of STKs and STPs with Transcriptional Regulators
3.2. The Role of STKs and STPs in Regulation of Transcription and Protein Biosynthesis
3.3. The Role of STKs and STPs in Cell Wall Architecture and Metabolism, Cellular Metabolism, Cell Division, and Adaptation to Stress Conditions
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| STK | serine/threonine kinase |
| STP | serine/threonine phosphatase |
| PPP | phosphoprotein phosphatase |
| PPM | metal-dependent phosphatase |
| TCS | two-component system |
| LUCA | last universal common ancestral |
| ATP | adenosine triphosphate |
| GTP | guanosine triphosphate |
| Ser | serine |
| Thr | threonine |
| Tyr | tyrosine |
| PASTA | penicillin-binding and Ser/Thr kinase-associated repeat |
| FHA | forkhead-associated domain |
| PBP | penicillin-binding protein |
| EF | elongation factor |
| MBP | myelin basic protein |
References
- DeVinney, R.; Steele-Mortimer, O.; Finlay, B.B. Phosphatases and kinases delivered to the host cell by bacterial pathogens. Trends Microbiol. 2000, 8, 29–33. [Google Scholar] [CrossRef]
- Rose, C.M.; Venkateshwaran, M.; Volkening, J.D.; Grimsrud, P.A.; Maeda, J.; Bailey, D.J.; Park, K.; Howes-Podoll, M.; den Os, D.; Yeun, L.H.; et al. Rapid phosphoproteomic and transcriptomic changes in the rhizobia-legume symbiosis. Mol. Cell Proteomics 2012, 11, 724–744. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I.; Grangeasse, C.; Turgay, K. Exploring the diversity of protein modifications: Special bacterial phosphorylation systems. FEMS Microbiol. Rev. 2016, 40, 398–417. [Google Scholar] [CrossRef] [PubMed]
- Francez-Charlot, A.; Kaczmarczyk, A.; Fischer, H.M.; Vorholt, J.A. The general stress response in Alphaproteobacteria. Trends Microbiol. 2015, 23, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, P.J. Protein kinases and protein phosphatases in prokaryotes: A genomic perspective. FEMS Microbiol. Lett. 2002, 206, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kennelly, P.J. Archaeal protein kinases and protein phosphatases: Insights from genomics and biochemistry. Biochem. J. 2003, 370, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.F.; Goss, L.; Dworkin, J. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. The origins of protein phosphorylation. Nat. Cell Biol. 2002, 4, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M. In the beginning, there was protein phosphorylation. J. Biol. Chem. 2014, 289, 9460–9462. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.A.; El Khoury, T.; Guiral, S.; Laaberki, M.H.; Candusso, M.; Galisson, F.; Foucher, A.E.; Kesraoui, S.; Ballut, L.; Vallet, S.; et al. Expanding the Kinome World: A New Protein Kinase Family Widely Conserved in Bacteria. J. Mol. Biol. 2017, 429, 3056–3074. [Google Scholar] [CrossRef] [PubMed]
- Libby, E.A.; Goss, L.A.; Dworkin, J. The Eukaryotic-Like Ser/Thr Kinase PrkC Regulates the Essential WalRK Two-Component System in Bacillus subtilis. PLoS Genet. 2015, 11, e1005275. [Google Scholar] [CrossRef] [PubMed]
- Brautigan, D.L. Protein Ser/Thr phosphatases—The ugly ducklings of cell signaling. FEBS J. 2013, 280, 324–345. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Serine/threonine phosphatases: Mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [PubMed]
- Mijakovic, I.; Macek, B. Impact of phosphoproteomics on studies of bacterial physiology. FEMS Microbiol. Rev. 2012, 36, 877–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dworkin, J. Ser/Thr phosphorylation as a regulatory mechanism in bacteria. Curr. Opin. Microbiol. 2015, 24, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennelly, P.J. Protein Ser/Thr/Tyr phosphorylation in the Archaea. J. Biol. Chem. 2014, 289, 9480–9487. [Google Scholar] [CrossRef] [PubMed]
- Esser, D.; Hoffmann, L.; Pham, T.K.; Bräsen, C.; Qiu, W.; Wright, P.C.; Albers, S.V.; Siebers, B. Protein phosphorylation and its role in archaeal signal transduction. FEMS Microbiol. Rev. 2016, 40, 625–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanks, S.K.; Quinn, A.M.; Hunter, T. The protein kinase family: Conserved features and deduced phylogeny of the catalytic domains. Science 1988, 241, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Dorado, J.; Inouye, S.; Inouye, M. A gene encoding a protein serine/threonine kinase is required for normal development of Myxococcus xanthus, a gram-negative bacterium. Cell 1991, 67, 995–1006. [Google Scholar] [CrossRef]
- Stancik, I.A.; Šestak, M.S.; Ji, B.; Axelson-Fisk, M.; Franjevic, D.; Jers, C.; Domazet-Lošo, T.; Mijakovic, I. Serine/Threonine Protein Kinases from Bacteria, Archaea and Eukarya Share a Common Evolutionary Origin Deeply Rooted in the Tree of Life. J. Mol. Biol. 2018, 430, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Brown, A.K.; Besra, G.S.; Cozzone, A.J.; Kremer, L. The condensing activities of the Mycobacterium tuberculosis type II fatty acid synthase are differentially regulated by phosphorylation. J. Biol. Chem. 2006, 281, 30094–30103. [Google Scholar] [CrossRef] [PubMed]
- Veyron-Churlet, R.; Zanella-Cléon, I.; Cohen-Gonsaud, M.; Molle, V.; Kremer, L. Phosphorylation of the Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein reductase MabA regulates mycolic acid biosynthesis. J. Biol. Chem. 2010, 285, 12714–12725. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.; Verma, S.K.; Khan, S.; Prakash, B.; Nandicoori, V.K. PknB-mediated phosphorylation of a novel substrate, N-acetylglucosamine-1-phosphate uridyltransferase, modulates its acetyltransferase activity. J. Mol. Biol. 2009, 386, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Sureka, K.; Hossain, T.; Mukherjee, P.; Chatterjee, P.; Datta, P.; Kundu, M.; Basu, J. Novel role of phosphorylation-dependent interaction between FtsZ and FipA in mycobacterial cell division. PLoS ONE 2010, 5, e8590. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Chakraborti, P.K. GTPase activity of mycobacterial FtsZ is impaired due to its transphosphorylation by the eukaryotic-type Ser/Thr kinase, PknA. J. Biol. Chem. 2006, 281, 40107–40113. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Chakraborti, P.K. Ability of PknA, a mycobacterial eukaryotic-type serine/threonine kinase, to transphosphorylate MurD, a ligase involved in the process of peptidoglycan biosynthesis. Biochem. J. 2008, 415, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.M.; Abbott, D.W.; Park, S.T.; Dascher, C.C.; Cantley, L.C.; Husson, R.N. The Mycobacterium tuberculosis serine/threonine kinases PknA and PknB: Substrate identification and regulation of cell shape. Genes Dev. 2005, 19, 1692–1704. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Gupta, M.; Krupa, A.; Srinivasan, N.; Singh, Y. EmbR, a regulatory protein with ATPase activity, is a substrate of multiple serine/threonine kinases and phosphatase in Mycobacterium tuberculosis. FEBS J. 2006, 273, 2711–2721. [Google Scholar] [CrossRef] [PubMed]
- Canova, M.J.; Kremer, L.; Molle, V. The Mycobacterium tuberculosis GroEL1 chaperone is a substrate of Ser/Thr protein kinases. J. Bacteriol. 2009, 191, 2876–2883. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Datta, P.; Kundu, M.; Basu, J. The serine/threonine kinase PknB of Mycobacterium tuberculosis phosphorylates PBPA, a penicillin-binding protein required for cell division. Microbiology 2006, 152, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Park, S.T.; Kang, C.M.; Husson, R.N. Regulation of the SigH stress response regulon by an essential protein kinase in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13105–13110. [Google Scholar] [CrossRef] [PubMed]
- Villarino, A.; Duran, R.; Wehenkel, A.; Fernandez, P.; England, P.; Brodin, P.; Cole, S.T.; Zimny-Arndt, U.; Jungblut, P.R.; Cerveñansky, C.; et al. Proteomic identification of M. tuberculosis protein kinase substrates: PknB recruits GarA, a FHA domain-containing protein, through activation loop-mediated interactions. J. Mol. Biol. 2005, 350, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Veyron-Churlet, R.; Molle, V.; Taylor, R.C.; Brown, A.K.; Besra, G.S.; Zanella-Cléon, I.; Fütterer, K.; Kremer, L. The Mycobacterium tuberculosis beta-ketoacyl-acyl carrier protein synthase III activity is inhibited by phosphorylation on a single threonine residue. J. Biol. Chem. 2009, 284, 6414–6424. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.; Garcia, R.; Bach, H.; de Waard, J.H.; Jacobs, W.R.; Av-Gay, Y.; Bubis, J.; Takiff, H.E. Mycobacterium tuberculosis transporter MmpL7 is a potential substrate for kinase PknD. Biochem. Biophys. Res. Commun. 2006, 348, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.P.; Ulijasz, A.T. Regulation of transcription by eukaryotic-like serine-threonine kinases and phosphatases in Gram-positive bacterial pathogens. Virulence 2014, 5, 863–885. [Google Scholar] [CrossRef] [PubMed]
- Grundner, C.; Gay, L.M.; Alber, T. Mycobacterium tuberculosis serine/threonine kinases PknB, PknD, PknE, and PknF phosphorylate multiple FHA domains. Protein Sci. 2005, 14, 1918–1921. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Soulat, D.; Jault, J.M.; Grangeasse, C.; Cozzone, A.J.; Prost, J.F. Two FHA domains on an ABC transporter, Rv1747, mediate its phosphorylation by PknF, a Ser/Thr protein kinase from Mycobacterium tuberculosis. FEMS Microbiol. Lett. 2004, 234, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Leiba, J.; Carrère-Kremer, S.; Blondiaux, N.; Dimala, M.M.; Wohlkönig, A.; Baulard, A.; Kremer, L.; Molle, V. The Mycobacterium tuberculosis transcriptional repressor EthR is negatively regulated by Serine/Threonine phosphorylation. Biochem. Biophys. Res. Commun. 2014, 446, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- O’Hare, H.M.; Durán, R.; Cerveñansky, C.; Bellinzoni, M.; Wehenkel, A.M.; Pritsch, O.; Obal, G.; Baumgartner, J.; Vialaret, J.; Johnsson, K.; et al. Regulation of glutamate metabolism by protein kinases in mycobacteria. Mol. Microbiol. 2008, 70, 1408–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Papavinasasundaram, K.G.; Av-Gay, Y. Novel substrates of Mycobacterium tuberculosis PknH Ser/Thr kinase. Biochem. Biophys. Res. Commun. 2007, 355, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Kremer, L.; Girard-Blanc, C.; Besra, G.S.; Cozzone, A.J.; Prost, J.F. An FHA phosphoprotein recognition domain mediates protein EmbR phosphorylation by PknH, a Ser/Thr protein kinase from Mycobacterium tuberculosis. Biochemistry 2003, 42, 15300–15309. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.D.; Papavinasasundaram, K.G.; Zheng, X.; Chávez-Steenbock, A.; Wang, X.; Lee, G.Q.; Av-Gay, Y. Convergence of Ser/Thr and two-component signaling to coordinate expression of the dormancy regulon in Mycobacterium tuberculosis. J. Biol. Chem. 2010, 285, 29239–29246. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Stella, A.; Boudou, F.; Levillain, F.; Darthuy, E.; Vaubourgeix, J.; Wang, C.; Bardou, F.; Puzo, G.; Gilleron, M.; et al. Functional characterization of the Mycobacterium tuberculosis serine/threonine kinase PknJ. Microbiology 2010, 156, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, D.; Parikh, A.; Rananaware, D.; Gupta, M.; Singh, Y.; Nandicoori, V.K. The Mycobacterium tuberculosis protein kinase K modulates activation of transcription from the promoter of mycobacterial monooxygenase operon through phosphorylation of the transcriptional regulator VirS. J. Biol. Chem. 2009, 284, 11090–11099. [Google Scholar] [CrossRef] [PubMed]
- Canova, M.J.; Veyron-Churlet, R.; Zanella-Cleon, I.; Cohen-Gonsaud, M.; Cozzone, A.J.; Becchi, M.; Kremer, L.; Molle, V. The Mycobacterium tuberculosis serine/threonine kinase PknL phosphorylates Rv2175c: Mass spectrometric profiling of the activation loop phosphorylation sites and their role in the recruitment of Rv2175c. Proteomics 2008, 8, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zou, W.; Fang, J.; Huang, X.; Gao, F.; He, Z.; Zhang, K.; Zhao, N. Eukaryote-like Ser/Thr protein kinase PrkA modulates sporulation via regulating the transcriptional factor σ(K) in Bacillus subtilis. Front. Microbiol. 2015, 6, 382. [Google Scholar] [CrossRef] [PubMed]
- Absalon, C.; Obuchowski, M.; Madec, E.; Delattre, D.; Holland, I.B.; Séror, S.J. CpgA, EF-Tu and the stressosome protein YezB are substrates of the Ser/Thr kinase/phosphatase couple, PrkC/PrpC, in Bacillus subtilis. Microbiology 2009, 155, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Pietack, N.; Becher, D.; Schmidl, S.R.; Saier, M.H.; Hecker, M.; Commichau, F.M.; Stülke, J. In vitro phosphorylation of key metabolic enzymes from Bacillus subtilis: PrkC phosphorylates enzymes from different branches of basic metabolism. J. Mol. Microbiol. Biotechnol. 2010, 18, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Pompeo, F.; Foulquier, E.; Galinier, A. Impact of Serine/Threonine Protein Kinases on the Regulation of Sporulation in Bacillus subtilis. Front. Microbiol. 2016, 7, 568. [Google Scholar] [CrossRef] [PubMed]
- Kobir, A.; Poncet, S.; Bidnenko, V.; Delumeau, O.; Jers, C.; Zouhir, S.; Grenha, R.; Nessler, S.; Noirot, P.; Mijakovic, I. Phosphorylation of Bacillus subtilis gene regulator AbrB modulates its DNA-binding properties. Mol. Microbiol. 2014, 92, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, V.; Shi, L.; Krug, K.; Derouiche, A.; Jers, C.; Cousin, C.; Kobir, A.; Mijakovic, I.; Macek, B. Quantitative phosphoproteome analysis of Bacillus subtilis reveals novel substrates of the kinase PrkC and phosphatase PrpC. Mol. Cell Proteomics 2014, 13, 1965–1978. [Google Scholar] [CrossRef] [PubMed]
- Gaidenko, T.A.; Kim, T.J.; Price, C.W. The PrpC serine-threonine phosphatase and PrkC kinase have opposing physiological roles in stationary-phase Bacillus subtilis cells. J. Bacteriol. 2002, 184, 6109–6114. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.M.; Dworkin, J. Induction and regulation of a secreted peptidoglycan hydrolase by a membrane Ser/Thr kinase that detects muropeptides. Mol. Microbiol. 2010, 75, 1232–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidnenko, V.; Shi, L.; Kobir, A.; Ventroux, M.; Pigeonneau, N.; Henry, C.; Trubuil, A.; Noirot-Gros, M.F.; Mijakovic, I. Bacillus subtilis serine/threonine protein kinase YabT is involved in spore development via phosphorylation of a bacterial recombinase. Mol. Microbiol. 2013, 88, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Derouiche, A.; Petranovic, D.; Macek, B.; Mijakovic, I. Bacillus subtilis single-stranded DNA-binding protein SsbA is phosphorylated at threonine 38 by the serine/threonine kinase YabT. Periodicum Biologorum. 2016, 118, 399–404. [Google Scholar] [CrossRef]
- Pereira, S.F.; Gonzalez, R.L.; Dworkin, J. Protein synthesis during cellular quiescence is inhibited by phosphorylation of a translational elongation factor. Proc. Natl. Acad. Sci. USA 2015, 112, E3274–E3281. [Google Scholar] [CrossRef] [PubMed]
- Bryant-Hudson, K.M.; Shakir, S.M.; Ballard, J.D. Autoregulatory characteristics of a Bacillus anthracis serine/threonine kinase. J. Bacteriol. 2011, 193, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Arora, G.; Sajid, A.; Arulanandh, M.D.; Singhal, A.; Mattoo, A.R.; Pomerantsev, A.P.; Leppla, S.H.; Maiti, S.; Singh, Y. Unveiling the novel dual specificity protein kinases in Bacillus anthracis: Identification of the first prokaryotic dual specificity tyrosine phosphorylation-regulated kinase (DYRK)-like kinase. J. Biol. Chem. 2012, 287, 26749–26763. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, M.; Canova, M.J.; Patin, D.; Letek, M.; Zanella-Cléon, I.; Becchi, M.; Mateos, L.M.; Mengin-Lecreulx, D.; Molle, V.; Gil, J.A. The MurC ligase essential for peptidoglycan biosynthesis is regulated by the serine/threonine protein kinase PknA in Corynebacterium glutamicum. J. Biol. Chem. 2008, 283, 36553–36563. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.; Niebisch, A.; Schwaiger, A.; Viets, U.; Metzger, S.; Bramkamp, M.; Bott, M. Genetic and biochemical analysis of the serine/threonine protein kinases PknA, PknB, PknG and PknL of Corynebacterium glutamicum: Evidence for non-essentiality and for phosphorylation of OdhI and FtsZ by multiple kinases. Mol. Microbiol. 2009, 74, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, M.; Canova, M.J.; Zanella-Cléon, I.; Becchi, M.; Cozzone, A.J.; Mateos, L.M.; Kremer, L.; Gil, J.A.; Molle, V. From the characterization of the four serine/threonine protein kinases (PknA/B/G/L) of Corynebacterium glutamicum toward the role of PknA and PknB in cell division. J. Biol. Chem. 2008, 283, 18099–18112. [Google Scholar] [CrossRef] [PubMed]
- Silvestroni, A.; Jewell, K.A.; Lin, W.; Connelly, J.E.; Ivancic, M.M.; Tao, W.A.; Rajagopal, L. Identification of serine/threonine kinase substrates in the human pathogen group B. Streptococcus. J. Proteome Res. 2009, 8, 2563–2574. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.J.; Walthers, D.; Connelly, J.E.; Burnside, K.; Jewell, K.A.; Kenney, L.J.; Rajagopal, L. Threonine phosphorylation prevents promoter DNA binding of the Group B Streptococcus response regulator CovR. Mol. Microbiol. 2009, 71, 1477–1495. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, L.; Vo, A.; Silvestroni, A.; Rubens, C.E. Regulation of cytotoxin expression by converging eukaryotic-type and two-component signaling mechanisms in Streptococcus agalactiae. Mol. Microbiol. 2006, 62, 941–957. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, L.; Clancy, A.; Rubens, C.E. A eukaryotic type serine/threonine kinase and phosphatase in Streptococcus agalactiae reversibly phosphorylate an inorganic pyrophosphatase and affect growth, cell segregation, and virulence. J. Biol. Chem. 2003, 278, 14429–14441. [Google Scholar] [CrossRef] [PubMed]
- Burnside, K.; Lembo, A.; Harrell, M.I.; Gurney, M.; Xue, L.; BinhTran, N.T.; Connelly, J.E.; Jewell, K.A.; Schmidt, B.Z.; de los Reyes, M.; et al. Serine/threonine phosphatase Stp1 mediates post-transcriptional regulation of hemolysin, autolysis, and virulence of group B Streptococcus. J. Biol. Chem. 2011, 286, 44197–44210. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Agarwal, S.; Pancholi, P.; Pancholi, V. Role of serine/threonine phosphatase (SP-STP) in Streptococcus pyogenes physiology and virulence. J. Biol. Chem. 2011, 286, 41368–41380. [Google Scholar] [CrossRef] [PubMed]
- Nováková, L.; Bezousková, S.; Pompach, P.; Spidlová, P.; Sasková, L.; Weiser, J.; Branny, P. Identification of multiple substrates of the StkP Ser/Thr protein kinase in Streptococcus pneumoniae. J. Bacteriol. 2010, 192, 3629–3638. [Google Scholar] [CrossRef] [PubMed]
- Giefing, C.; Jelencsics, K.E.; Gelbmann, D.; Senn, B.M.; Nagy, E. The pneumococcal eukaryotic-type serine/threonine protein kinase StkP co-localizes with the cell division apparatus and interacts with FtsZ in vitro. Microbiology 2010, 156, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulijasz, A.T.; Falk, S.P.; Weisblum, B. Phosphorylation of the RitR DNA-binding domain by a Ser-Thr phosphokinase: Implications for global gene regulation in the streptococci. Mol. Microbiol. 2009, 71, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Falk, S.P.; Weisblum, B. Phosphorylation of the Streptococcus pneumoniae cell wall biosynthesis enzyme MurC by a eukaryotic-like Ser/Thr kinase. FEMS Microbiol. Lett. 2013, 340, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Arcondéguy, T.; Bastide, A.; Touriol, C.; Prats, H.; Trombe, M.C. The StkP/PhpP signaling couple in Streptococcus pneumoniae: Cellular organization and physiological characterization. J. Bacteriol. 2009, 191, 4943–4950. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Vasudhev, S.; DeOliveira, R.B.; Ram, S. Inhibition of the classical pathway of complement by meningococcal capsular polysaccharides. J. Immunol. 2014, 193, 1855–1863. [Google Scholar] [CrossRef] [PubMed]
- Nováková, L.; Sasková, L.; Pallová, P.; Janecek, J.; Novotná, J.; Ulrych, A.; Echenique, J.; Trombe, M.C.; Branny, P. Characterization of a eukaryotic type serine/threonine protein kinase and protein phosphatase of Streptococcus pneumoniae and identification of kinase substrates. FEBS J. 2005, 272, 1243–1254. [Google Scholar] [CrossRef] [PubMed]
- Banu, L.D.; Conrads, G.; Rehrauer, H.; Hussain, H.; Allan, E.; van der Ploeg, J.R. The Streptococcus mutans serine/threonine kinase, PknB, regulates competence development, bacteriocin production, and cell wall metabolism. Infect. Immun. 2010, 78, 2209–2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomas-Lopez, R.; Paracuellos, P.; Riberty, M.; Cozzone, A.J.; Duclos, B. Several enzymes of the central metabolism are phosphorylated in Staphylococcus aureus. FEMS Microbiol. Lett. 2007, 272, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Truong-Bolduc, Q.C.; Ding, Y.; Hooper, D.C. Posttranslational modification influences the effects of MgrA on norA expression in Staphylococcus aureus. J. Bacteriol. 2008, 190, 7375–7381. [Google Scholar] [CrossRef] [PubMed]
- Didier, J.P.; Cozzone, A.J.; Duclos, B. Phosphorylation of the virulence regulator SarA modulates its ability to bind DNA in Staphylococcus aureus. FEMS Microbiol. Lett. 2010, 306, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Ding, Y.; Ji, Q.; Liang, Z.; Deng, X.; Wong, C.C.; Yi, C.; Zhang, L.; Xie, S.; Alvarez, S.; et al. Protein cysteine phosphorylation of SarA/MgrA family transcriptional regulators mediates bacterial virulence and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 15461–15466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiba, J.; Hartmann, T.; Cluzel, M.E.; Cohen-Gonsaud, M.; Delolme, F.; Bischoff, M.; Molle, V. A novel mode of regulation of the Staphylococcus aureus catabolite control protein A (CcpA) mediated by Stk1 protein phosphorylation. J. Biol. Chem. 2012, 287, 43607–43619. [Google Scholar] [CrossRef] [PubMed]
- Canova, M.J.; Baronian, G.; Brelle, S.; Cohen-Gonsaud, M.; Bischoff, M.; Molle, V. A novel mode of regulation of the Staphylococcus aureus Vancomycin-resistance-associated response regulator VraR mediated by Stk1 protein phosphorylation. Biochem. Biophys. Res. Commun. 2014, 447, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Fridman, M.; Williams, G.D.; Muzamal, U.; Hunter, H.; Siu, K.W.; Golemi-Kotra, D. Two unique phosphorylation-driven signaling pathways crosstalk in Staphylococcus aureus to modulate the cell-wall charge: Stk1/Stp1 meets GraSR. Biochemistry 2013, 52, 7975–7986. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Brelle, S.; Minatelli, S.; Molle, V. Stk1-mediated phosphorylation stimulates the DNA-binding properties of the Staphylococcus aureus SpoVG transcriptional factor. Biochem. Biophys. Res. Commun. 2016, 473, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Fan, J.; Niu, C.; Wang, D.; Wang, J.; Wang, X.; Villaruz, A.E.; Li, M.; Otto, M.; Gao, Q. The Eukaryotic-Type Serine/Threonine Protein Kinase Stk Is Required for Biofilm Formation and Virulence in Staphylococcus epidermidis. PLoS ONE 2011, 6, e25380. [Google Scholar] [CrossRef] [PubMed]
- Schmidl, S.R.; Gronau, K.; Hames, C.; Busse, J.; Becher, D.; Hecker, M.; Stülke, J. The stability of cytadherence proteins in Mycoplasma pneumoniae requires activity of the protein kinase PrkC. Infect. Immun. 2010, 78, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Gocayne, J.D.; White, O.; Adams, M.D.; Clayton, R.A.; Fleischmann, R.D.; Bult, C.J.; Kerlavage, A.R.; Sutton, G.; Kelley, J.M.; et al. The minimal gene complement of Mycoplasma genitalium. Science 1995, 270, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Sacco, E.; Cortes, M.; Josseaume, N.; Rice, L.B.; Mainardi, J.L.; Arthur, M. Serine/threonine protein phosphatase-mediated control of the peptidoglycan cross-linking l,d-transpeptidase pathway in Enterococcus faecium. mBio 2014, 5, e01414–e01446. [Google Scholar] [CrossRef] [PubMed]
- Desbonnet, C.; Tait-Kamradt, A.; Garcia-Solache, M.; Dunman, P.; Coleman, J.; Arthur, M.; Rice, L.B. Involvement of the Eukaryote-Like Kinase-Phosphatase System and a Protein That Interacts with Penicillin-Binding Protein 5 in Emergence of Cephalosporin Resistance in Cephalosporin-Sensitive Class A Penicillin-Binding Protein Mutants in Enterococcus faecium. mBio 2016, 7, e02115–e02188. [Google Scholar] [CrossRef]
- Udo, H.; Lam, C.K.; Mori, S.; Inouye, M.; Inouye, S. Identification of a substrate for Pkn2, a protein Ser/Thr kinase from Myxococcus xanthus by a novel method for substrate identification. J. Mol. Microbiol. Biotechnol. 2000, 2, 557–563. [Google Scholar] [PubMed]
- Nariya, H.; Inouye, S. Activation of 6-phosphofructokinase via phosphorylation by Pkn4, a protein Ser/Thr kinase of Myxococcus xanthus. Mol. Microbiol. 2002, 46, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Inouye, M.; Inouye, S. Reciprocal regulation of the differentiation of Myxococcus xanthus by Pkn5 and Pkn6, eukaryotic-like Ser/Thr protein kinases. Mol. Microbiol. 1996, 20, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Nariya, H.; Inouye, S. Identification of a protein Ser/Thr kinase cascade that regulates essential transcriptional activators in Myxococcus xanthus development. Mol. Microbiol. 2005, 58, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Kamei, A.; Yuasa, T.; Orikawa, K.; Geng, X.X.; Ikeuchi, M. A eukaryotic-type protein kinase, SpkA, is required for normal motility of the unicellular Cyanobacterium synechocystis sp. strain PCC 6803. J. Bacteriol. 2001, 183, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Panichkin, V.B.; Arakawa-Kobayashi, S.; Kanaseki, T.; Suzuki, I.; Los, D.A.; Shestakov, S.V.; Murata, N. Serine/threonine protein kinase SpkA in Synechocystis sp. strain PCC 6803 is a regulator of expression of three putative pilA operons, formation of thick pili, and cell motility. J. Bacteriol. 2006, 188, 7696–7699. [Google Scholar] [CrossRef] [PubMed]
- Mata-Cabana, A.; García-Domínguez, M.; Florencio, F.J.; Lindahl, M. Thiol-based redox modulation of a cyanobacterial eukaryotic-type serine/threonine kinase required for oxidative stress tolerance. Antioxid. Redox Signal 2012, 17, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Zorina, A.; Stepanchenko, N.; Novikova, G.V.; Sinetova, M.; Panichkin, V.B.; Moshkov, I.E.; Zinchenko, V.V.; Shestakov, S.V.; Suzuki, I.; Murata, N.; et al. Eukaryotic-like Ser/Thr protein kinases SpkC/F/K are involved in phosphorylation of GroES in the Cyanobacterium synechocystis. DNA Res. 2011, 18, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Jang, J.; Janicki, A.; Zhang, C.C.; Bédu, S. Inactivation of spkD, encoding a Ser/Thr kinase, affects the pool of the TCA cycle metabolites in Synechocystis sp. strain PCC 6803. Microbiology 2008, 154, 2161–2167. [Google Scholar] [CrossRef] [PubMed]
- Zorina, A.A.; Bedbenov, V.S.; Novikova, G.V.; Panichkin, V.B.; Los, D.A. Involvement of serine/threonine protein kinases in cold stress response in the cyanobacterium Synechocystis sp. PCC 6803: Functional characterization of a protein kinase Spke. Mol. Biol. (Mosk) 2014, 48, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhang, X.; Chi, X.; Guan, X.; Li, Y.; Qin, S.; Shao, H.B. Serine/threonine protein kinase SpkG is a candidate for high salt resistance in the unicellular cyanobacterium Synechocystis sp. PCC 6803. PLoS ONE 2011, 6, e18718. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.; Phalip, V.; Zhang, C.C. Characterization of PknC, a Ser/Thr kinase with broad substrate specificity from the cyanobacterium Anabaena sp. strain PCC 7120. Eur. J. Biochem. 2001, 268, 1869–7185. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Golden, J.W. Overexpression of pknE blocks heterocyst development in Anabaena sp. strain PCC 7120. J. Bacteriol. 2011, 193, 2619–2629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.C.; Libs, L. Cloning and characterization of the pknD gene encoding an eukaryotic-type protein kinase in the cyanobacterium Anabaena sp. PCC7120. Mol. Gen. Genet. 1998, 258, 26–33. [Google Scholar] [PubMed]
- Zhang, C.C.; Friry, A.; Peng, L. Molecular and genetic analysis of two closely linked genes that encode, respectively, a protein phosphatase 1/2A/2B homolog and a protein kinase homolog in the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 1998, 180, 2616–2622. [Google Scholar]
- Ehira, S.; Ohmori, M. The pknH gene restrictively expressed in heterocysts is required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2012, 158, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.; Durán, R.; Schujman, G.E.; Marchissio, M.J.; Portela, M.M.; Obal, G.; Pritsch, O.; de Mendoza, D.; Cerveñansky, C. Serine/threonine protein kinase PrkA of the human pathogen Listeria monocytogenes: Biochemical characterization and identification of interacting partners through proteomic approaches. J. Proteomics 2011, 74, 1720–1734. [Google Scholar] [CrossRef] [PubMed]
- Pensinger, D.A.; Boldon, K.M.; Chen, G.Y.; Vincent, W.J.; Sherman, K.; Xiong, M.; Schaenzer, A.J.; Forster, E.R.; Coers, J.; Striker, R.; et al. The Listeria monocytogenes PASTA kinase PrkA and its substrate YvcK are required for cell wall homeostasis, metabolism, and virulence. PLoS Pathog. 2016, 12, e1006001. [Google Scholar] [CrossRef] [PubMed]
- Mougous, J.D.; Gifford, C.A.; Ramsdell, T.L.; Mekalanos, J.J. Threonine phosphorylation post-translationally regulates protein secretion in Pseudomonas aeruginosa. Nat. Cell. Biol. 2007, 9, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Kapatral, V.; Xu, W.; Chakrabarty, A.M. Characterization of a Hank’s type serine/threonine kinase and serine/threonine phosphoprotein phosphatase in Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 6615–6622. [Google Scholar] [PubMed]
- Juris, S.J.; Rudolph, A.E.; Huddler, D.; Orth, K.; Dixon, J.E. A distinctive role for the Yersinia protein kinase: Actin binding, kinase activation, and cytoskeleton disruption. Proc. Natl. Acad. Sci. USA 2000, 97, 9431–9436. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, Z.; Chen, F.; Liu, X.; Ning, N.; Huang, J.; Wang, H. Eukaryotic-like Kinase Expression in Enterohemorrhagic Escherichia coli: Potential for Enhancing Host Aggressive Inflammatory Response. J. Infect. Dis. 2017, 216, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Rachwał, K.; Matczyńska, E.; Janczarek, M. Transcriptome profiling of a Rhizobium leguminosarum bv. trifolii rosR mutant reveals the role of the transcriptional regulator RosR in motility, synthesis of cell-surface components, and other cellular processes. BMC Genomics 2015, 16, 1111. [Google Scholar] [CrossRef]
- Hanks, S.K.; Hunter, T. Protein kinases 6. The eukaryotic protein kinase superfamily: Kinase (catalytic) domain structure and classification. FASEB J. 1995, 9, 576–596. [Google Scholar] [CrossRef] [PubMed]
- Kornev, A.P.; Taylor, S.S. Defining the conserved internal architecture of a protein kinase. Biochim. Biophys. Acta 2010, 1804, 440–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krupa, A.; Srinivasan, N. Diversity in domain architectures of Ser/Thr kinases and their homologues in prokaryotes. BMC Genomics 2005, 6, 129. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Lombardía, M.; Pompeo, F.; Boitel, B.; Alzari, P.M. Crystal structure of the catalytic domain of the PknB serine/threonine kinase from Mycobacterium tuberculosis. J. Biol. Chem. 2003, 278, 13094–13100. [Google Scholar] [CrossRef] [PubMed]
- Young, K.D. Bacterial shape. Mol. Microbiol. 2003, 49, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Young, T.A.; Delagoutte, B.; Endrizzi, J.A.; Falick, A.M.; Alber, T. Structure of Mycobacterium tuberculosis PknB supports a universal activation mechanism for Ser/Thr protein kinases. Nat. Struct. Biol. 2003, 10, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, A.E.; Echols, N.; Lombana, T.N.; King, D.S.; Alber, T. Allosteric activation by dimerization of the PknD receptor Ser/Thr protein kinase from Mycobacterium tuberculosis. J. Biol. Chem. 2007, 282, 11427–11435. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, A.E.; Grundner, C.; Echols, N.; Gay, L.M.; Lombana, T.N.; Miecskowski, C.A.; Pullen, K.E.; Sung, P.Y.; Alber, T. Structure/function studies of Ser/Thr and Tyr protein phosphorylation in Mycobacterium tuberculosis. J. Mol. Microbiol. Biotechnol. 2005, 9, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Mieczkowski, C.; Iavarone, A.T.; Alber, T. Auto-activation mechanism of the Mycobacterium tuberculosis PknB receptor Ser/Thr kinase. EMBO J. 2008, 27, 3186–3197. [Google Scholar] [CrossRef] [PubMed]
- Pérez, J.; Castañeda-García, A.; Jenke-Kodama, H.; Müller, R.; Muñoz-Dorado, J. Eukaryotic-like protein kinases in the prokaryotes and the myxobacterial kinome. Proc. Natl. Acad. Sci. USA 2008, 105, 15950–15955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrícková, K.; Petrícek, M. Eukaryotic-type protein kinases in Streptomyces coelicolor: Variations on a common theme. Microbiology 2003, 149, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Yeats, C.; Finn, R.D.; Bateman, A. The PASTA domain: A beta-lactam-binding domain. Trends Biochem. Sci. 2002, 27, 438–440. [Google Scholar] [CrossRef]
- Dessen, A.; Mouz, N.; Gordon, E.; Hopkins, J.; Dideberg, O. Crystal structure of PBP2x from a highly penicillin-resistant Streptococcus pneumoniae clinical isolate: A mosaic framework containing 83 mutations. J. Biol. Chem. 2001, 276, 45106–45112. [Google Scholar] [CrossRef] [PubMed]
- Good, M.C.; Greenstein, A.E.; Young, T.A.; Ng, H.L.; Alber, T. Sensor domain of the Mycobacterium tuberculosis receptor Ser/Thr protein kinase, PknD, forms a highly symmetric beta propeller. J. Mol. Biol. 2004, 339, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Barthe, P.; Mukamolova, G.V.; Roumestand, C.; Cohen-Gonsaud, M. The structure of PknB extracellular PASTA domain from Mycobacterium tuberculosis suggests a ligand-dependent kinase activation. Structure 2010, 18, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Paracuellos, P.; Ballandras, A.; Robert, X.; Kahn, R.; Hervé, M.; Mengin-Lecreulx, D.; Cozzone, A.J.; Duclos, B.; Gouet, P. The extended conformation of the 2.9-Å crystal structure of the three-PASTA domain of a Ser/Thr kinase from the human pathogen Staphylococcus aureus. J. Mol. Biol. 2010, 404, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Shah, I.M.; Laaberki, M.H.; Popham, D.L.; Dworkin, J. A eukaryotic-like Ser/Thr kinase signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell 2008, 135, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Cozzone, A.J. Role of protein phosphorylation on serine/threonine and tyrosine in the virulence of bacterial pathogens. J. Mol. Microbiol. Biotechnol. 2005, 9, 198–213. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Wells, C.L.; Dunny, G.M. A eukaryotic-type Ser/Thr kinase in Enterococcus faecalis mediates antimicrobial resistance and intestinal persistence. Proc. Natl. Acad. Sci. USA 2007, 104, 3508–3513. [Google Scholar] [CrossRef] [PubMed]
- Wehenkel, A.; Bellinzoni, M.; Graña, M.; Duran, R.; Villarino, A.; Fernandez, P.; Andre-Leroux, G.; England, P.; Takiff, H.; Cerveñansky, C.; et al. Mycobacterial Ser/Thr protein kinases and phosphatases: Physiological roles and therapeutic potential. Biochim. Biophys. Acta 2008, 1784, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Molle, V.; Kremer, L. Division and cell envelope regulation by Ser/Thr phosphorylation: Mycobacterium shows the way. Mol. Microbiol. 2010, 75, 1064–1077. [Google Scholar] [CrossRef] [PubMed]
- Ohlsen, K.; Donat, S. The impact of serine/threonine phosphorylation in Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Sajid, A.; Arora, G.; Singhal, A.; Kalia, V.C.; Singh, Y. Protein Phosphatases of Pathogenic Bacteria: Role in Physiology and Virulence. Annu. Rev. Microbiol. 2015, 69, 527–547. [Google Scholar] [CrossRef] [PubMed]
- Stock, J.B.; Stock, A.M.; Mottonen, J.M. Signal transduction in bacteria. Nature 1990, 344, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.; Arora, D.; Singh, L.K.; Gangwal, A.; Sajid, A.; Molle, V.; Singh, Y.; Nandicoori, V.K. Serine/Threonine Protein Phosphatase PstP of Mycobacterium tuberculosis Is Necessary for Accurate Cell Division and Survival of Pathogen. J. Biol. Chem. 2016, 291, 24215–24230. [Google Scholar] [CrossRef] [PubMed]
- Boitel, B.; Ortiz-Lombardía, M.; Durán, R.; Pompeo, F.; Cole, S.T.; Cerveñansky, C.; Alzari, P.M. PknB kinase activity is regulated by phosphorylation in two Thr residues and dephosphorylation by PstP, the cognate phospho-Ser/Thr phosphatase, in Mycobacterium tuberculosis. Mol. Microbiol. 2003, 49, 1493–1508. [Google Scholar] [CrossRef] [PubMed]
- Arora, G.; Sajid, A.; Gupta, M.; Bhaduri, A.; Kumar, P.; Basu-Modak, S.; Singh, Y. Understanding the role of PknJ in Mycobacterium tuberculosis: Biochemical characterization and identification of novel substrate pyruvate kinase A. PLoS ONE 2010, 5, e10772. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Arora, G.; Sajid, A.; Maji, A.; Bhat, A.; Virmani, R.; Upadhyay, S.; Nandicoori, V.K.; Sengupta, S.; Singh, Y. Regulation of homocysteine metabolism by Mycobacterium tuberculosis S-adenosylhomocysteine hydrolase. Sci. Rep. 2013, 3, 2264. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Sajid, A.; Arora, G.; Tandon, V.; Singh, Y. Forkhead-associated domain-containing protein Rv0019c and polyketide-associated protein PapA5, from substrates of serine/threonine protein kinase PknB to interacting proteins of Mycobacterium tuberculosis. J. Biol. Chem. 2009, 284, 34723–34734. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.D.; Halbedel, S.; Görke, B.; Stülke, J. Control of the phosphorylation state of the HPr protein of the phosphotransferase system in Bacillus subtilis: Implication of the protein phosphatase PrpC. J. Mol. Microbiol. Biotechnol. 2007, 13, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kang, C.M.; Brody, M.S.; Price, C.W. Opposing pairs of serine protein kinases and phosphatases transmit signals of environmental stress to activate a bacterial transcription factor. Genes Dev. 1996, 10, 2265–2275. [Google Scholar] [CrossRef] [PubMed]
- Obuchowski, M.; Madec, E.; Delattre, D.; Boël, G.; Iwanicki, A.; Foulger, D.; Séror, S.J. Characterization of PrpC from Bacillus subtilis, a member of the PPM phosphatase family. J. Bacteriol. 2000, 182, 5634–5638. [Google Scholar] [CrossRef] [PubMed]
- Shakir, S.M.; Bryant, K.M.; Larabee, J.L.; Hamm, E.E.; Lovchik, J.; Lyons, C.R.; Ballard, J.D. Regulatory Interactions of a Virulence-Associated Serine/Threonine Phosphatase-Kinase Pair in Bacillus anthracis. J. Bacteriol. 2010, 192, 400–409. [Google Scholar] [CrossRef] [PubMed]
- Fleurie, A.; Lesterlin, C.; Manuse, S.; Zhao, C.; Cluzel, C.; Lavergne, J.P.; Franz-Wachtel, M.; Macek, B.; Combet, C.; Kuru, E.; et al. MapZ marks the division sites and positions FtsZ rings in Streptococcus pneumoniae. Nature 2014, 516, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Agarwal, S.; Pancholi, P.; Pancholi, V. Strain-specific regulatory role of eukaryote-like serine/threonine phosphatase in pneumococcal adherence. Infect. Immun. 2012, 80, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, K.; Nováková, L.; Fadda, D.; Branny, P.; Massidda, O.; Veening, J.W. Control of cell division in Streptococcus pneumoniae by the conserved Ser/Thr protein kinase StkP. Proc. Natl. Acad. Sci. USA 2012, 109, E905–E913. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Pancholi, V. Identification and biochemical characterization of a eukaryotic-type serine/threonine kinase and its cognate phosphatase in Streptococcus pyogenes: Their biological functions and substrate identification. J. Mol. Biol. 2006, 357, 1351–1372. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Branny, P.; Allan, E. A eukaryotic-type serine/threonine protein kinase is required for biofilm formation, genetic competence, and acid resistance in Streptococcus mutans. J. Bacteriol. 2006, 188, 1628–1632. [Google Scholar] [CrossRef] [PubMed]
- Beltramini, A.M.; Mukhopadhyay, C.D.; Pancholi, V. Modulation of cell wall structure and antimicrobial susceptibility by a Staphylococcus aureus eukaryote-like serine/threonine kinase and phosphatase. Infect. Immun. 2009, 77, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Burnside, K.; Lembo, A.; de Los Reyes, M.; Iliuk, A.; Binhtran, N.T.; Connelly, J.E.; Lin, W.J.; Schmidt, B.Z.; Richardson, A.R.; Fang, F.C.; et al. Regulation of hemolysin expression and virulence of Staphylococcus aureus by a serine/threonine kinase and phosphatase. PLoS ONE 2010, 5, e11071. [Google Scholar] [CrossRef] [PubMed]
- Liebeke, M.; Meyer, H.; Donat, S.; Ohlsen, K.; Lalk, M. A metabolomic view of Staphylococcus aureus and its Ser/Thr kinase and phosphatase deletion mutants: Involvement in cell wall biosynthesis. Chem. Biol. 2010, 17, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Treuner-Lange, A.; Ward, M.J.; Zusman, D.R. Pph1 from Myxococcus xanthus is a protein phosphatase involved in vegetative growth and development. Mol. Microbiol. 2001, 40, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Halbedel, S.; Busse, J.; Schmidl, S.R.; Stülke, J. Regulatory protein phosphorylation in Mycoplasma pneumoniae. A PP2C-type phosphatase serves to dephosphorylate HPr (Ser-P.). J. Biol. Chem. 2006, 281, 26253–26259. [Google Scholar] [CrossRef] [PubMed]
- Menegatti, A.C.; Vernal, J.; Terenzi, H. The unique serine/threonine phosphatase from the minimal bacterium Mycoplasma synoviae: Biochemical characterization and metal dependence. J. Biol. Inorg. Chem. 2015, 20, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.A.; Das, K.; Saikolappan, S.; Materon, L.A.; Dhandayuthapani, S. A serine/threonine phosphatase encoded by MG_207 of Mycoplasma genitalium is critical for its virulence. BMC Microbiol. 2013, 13, 44. [Google Scholar] [CrossRef] [PubMed]
- Archambaud, C.; Gouin, E.; Pizarro-Cerda, J.; Cossart, P.; Dussurget, O. Translation elongation factor EF-Tu is a target for Stp, a serine-threonine phosphatase involved in virulence of Listeria monocytogenes. Mol. Microbiol. 2005, 56, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Archambaud, C.; Nahori, M.A.; Pizarro-Cerda, J.; Cossart, P.; Dussurget, O. Control of Listeria superoxide dismutase by phosphorylation. J. Biol. Chem. 2006, 281, 31812–31822. [Google Scholar] [CrossRef] [PubMed]
- Ueda, A.; Wood, T.K. Connecting quorum sensing, c-di-GMP, pel polysaccharide, and biofilm formation in Pseudomonas aeruginosa through tyrosine phosphatase TpbA (PA3885). PLoS Pathog. 2009, 5, e1000483. [Google Scholar] [CrossRef] [PubMed]
- Missiakas, D.; Raina, S. Signal transduction pathways in response to protein misfolding in the extracytoplasmic compartments of E. coli: Role of two new phosphoprotein phosphatases PrpA and PrpB. EMBO J. 1997, 16, 1670–1685. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.M.; Le Moual, H. PrpZ, a Salmonella enterica serovar Typhi serine/threonine protein phosphatase 2C with dual substrate specificity. Microbiology 2005, 151, 1159–1167. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Bischoff, K.M.; Kennelly, P.J. The icfG gene cluster of Synechocystis sp. strain PCC 6803 encodes an Rsb/Spo-like protein kinase, protein phosphatase, and two phosphoproteins. J. Bacteriol. 1999, 181, 4761–4767. [Google Scholar] [PubMed]
- Kloft, N.; Rasch, G.; Forchhammer, K. Protein phosphatase PphA from Synechocystis sp. PCC 6803: The physiological framework of PII-P dephosphorylation. Microbiology 2005, 151, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Tom, S.K.; Callahan, S.M. The putative phosphatase All1758 is necessary for normal growth, cell size and synthesis of the minor heterocyst-specific glycolipid in the cyanobacterium Anabaena sp. strain PCC 7120. Microbiology 2012, 158, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Lipa, P.; Vinardell, J.M.; Kopcińska, J.; Zdybicka-Barabas, A.; Janczarek, M. Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover. Genes (Basel) 2018, 9, E369. [Google Scholar] [CrossRef] [PubMed]
- Burnside, K.; Rajagopal, L. Regulation of prokaryotic gene expression by eukaryotic-like enzymes. Curr. Opin. Microbiol. 2012, 15, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Carmichael, W.W. pp1-cyano2, a protein serine/threonine phosphatase 1 gene from the cyanobacterium Microcystis aeruginosa UTEX 2063. Arch. Microbiol. 1997, 168, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Carmichael, W.W.; Kennelly, P.J. Cyanobacterial PPP family protein phosphatases possess multifunctional capabilities and are resistant to microcystin-LR. J. Biol. Chem. 1999, 274, 10039–10046. [Google Scholar] [CrossRef] [PubMed]
- Bork, P.; Brown, N.P.; Hegyi, H.; Schultz, J. The protein phosphatase 2C (PP2C) superfamily: Detection of bacterial homologues. Protein Sci. 1996, 5, 1421–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Shi, L.; Li, L.; Guo, S.; Zhang, X.; Yamasaki, S.; Miyoshi, S.; Shinoda, S. Identification and characterization of class 1 integron resistance gene cassettes among Salmonella strains isolated from healthy humans in China. Microbiol. Immunol. 2004, 48, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Shi, L. Manganese-dependent protein O-phosphatases in prokaryotes and their biological functions. Front. Biosci. 2004, 9, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Das, A.K.; Helps, N.R.; Cohen, P.T.; Barford, D. Crystal structure of the protein serine/threonine phosphatase 2C at 2.0 A. resolution. EMBO J. 1996, 15, 6798–6809. [Google Scholar] [CrossRef] [PubMed]
- Alzari, P.M. First structural glimpse at a bacterial Ser/Thr protein phosphatase. Structure 2004, 12, 1923–1924. [Google Scholar] [CrossRef] [PubMed]
- Pullen, K.E.; Ng, H.L.; Sung, P.Y.; Good, M.C.; Smith, S.M.; Alber, T. An alternate conformation and a third metal in PstP/Ppp, the M. tuberculosis PP2C-Family Ser/Thr protein phosphatase. Structure 2004, 12, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Potts, M.; Kennelly, P.J. The serine, threonine, and/or tyrosine-specific protein kinases and protein phosphatases of prokaryotic organisms: A family portrait. FEMS Microbiol. Rev. 1998, 22, 229–253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shi, L. Evolution of the PPM-family protein phosphatases in Streptomyces: Duplication of catalytic domain and lateral recruitment of additional sensory domains. Microbiology 2004, 150, 4189–4197. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Wang, Y. Functional diversity of mammalian type 2C protein phosphatase isoforms: New tales from an old family. Clin. Exp. Pharmacol. Physiol. 2008, 35, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Wehenkel, A.; Bellinzoni, M.; Schaeffer, F.; Villarino, A.; Alzari, P.M. Structural and binding studies of the three-metal center in two mycobacterial PPM Ser/Thr protein phosphatases. J. Mol. Biol. 2007, 374, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Rantanen, M.K.; Lehtiö, L.; Rajagopal, L.; Rubens, C.E.; Goldman, A. Structure of Streptococcus agalactiae serine/threonine phosphatase. The subdomain conformation is coupled to the binding of a third metal ion. FEBS J. 2007, 274, 3128–3137. [Google Scholar] [CrossRef] [PubMed]
- Schlicker, C.; Fokina, O.; Kloft, N.; Grüne, T.; Becker, S.; Sheldrick, G.M.; Forchhammer, K. Structural analysis of the PP2C phosphatase tPphA from Thermosynechococcus elongatus: A flexible flap subdomain controls access to the catalytic site. J. Mol. Biol. 2008, 376, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Madec, E.; Laszkiewicz, A.; Iwanicki, A.; Obuchowski, M.; Séror, S. Characterization of a membrane-linked Ser/Thr protein kinase in Bacillus subtilis, implicated in developmental processes. Mol. Microbiol. 2002, 46, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.M.; Cieslewicz, M.J.; Kasper, D.L.; Wessels, M.R. Regulation of virulence by a two-component system in group B Streptococcus. J. Bacteriol. 2005, 187, 1105–1513. [Google Scholar] [CrossRef] [PubMed]
- Lamy, M.C.; Zouine, M.; Fert, J.; Vergassola, M.; Couve, E.; Pellegrini, E.; Glaser, P.; Kunst, F.; Msadek, T.; Trieu-Cuot, P.; et al. CovS/CovR of group B Streptococcus: A two-component global regulatory system involved in virulence. Mol. Microbiol. 2004, 54, 1250–1268. [Google Scholar] [CrossRef] [PubMed]
- Gryllos, I.; Grifantini, R.; Colaprico, A.; Jiang, S.; Deforce, E.; Hakansson, A.; Telford, J.L.; Grandi, G.; Wessels, M.R. Mg(2+) signaling defines the group A streptococcal CsrRS (CovRS) regulon. Mol. Microbiol. 2007, 65, 671–683. [Google Scholar] [CrossRef] [PubMed]
- Bugrysheva, J.; Froehlich, B.J.; Freiberg, J.A.; Scott, J.R. Serine/threonine protein kinase Stk is required for virulence, stress response, and penicillin tolerance in Streptococcus pyogenes. Infect. Immun. 2011, 79, 4201–4209. [Google Scholar] [CrossRef] [PubMed]
- Ulijasz, A.T.; Andes, D.R.; Glasner, J.D.; Weisblum, B. Regulation of iron transport in Streptococcus pneumoniae by RitR, an orphan response regulator. J. Bacteriol. 2004, 186, 8123–8136. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.; Manna, A.C.; Projan, S.J.; Cheung, A.L. SarA, a global regulator of virulence determinants in Staphylococcus aureus, binds to a conserved motif essential for Sar-dependent gene regulation. J. Biol. Chem. 1999, 274, 37169–37176. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.L.; Nishina, K.; Manna, A.C. SarA of Staphylococcus aureus binds to the sarA promoter to regulate gene expression. J. Bacteriol. 2008, 190, 2239–2243. [Google Scholar] [CrossRef] [PubMed]
- Donat, S.; Streker, K.; Schirmeister, T.; Rakette, S.; Stehle, T.; Liebeke, M.; Lalk, M.; Ohlsen, K. Transcriptome and functional analysis of the eukaryotic-type serine/threonine kinase PknB in Staphylococcus aureus. J. Bacteriol. 2009, 191, 4056–4069. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Hooper, D.C. A new two-component regulatory system involved in adhesion, autolysis, and extracellular proteolytic activity of Staphylococcus aureus. J. Bacteriol. 2000, 182, 3955–3964. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.; Truong-Bolduc, Q.C.; Zhang, X.; Hooper, D.C. A mutation in the 5′ untranslated region increases stability of norA mRNA, encoding a multidrug resistance transporter of Staphylococcus aureus. J. Bacteriol. 2001, 183, 2367–2371. [Google Scholar] [CrossRef] [PubMed]
- Kaatz, G.W.; McAleese, F.; Seo, S.M. Multidrug resistance in Staphylococcus aureus due to overexpression of a novel multidrug and toxin extrusion (MATE) transport protein. Antimicrob. Agents Chemother. 2005, 49, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, M. Recent advances in the social and developmental biology of the myxobacteria. Microbiol. Rev. 1996, 60, 70–102. [Google Scholar] [PubMed]
- Nariya, H.; Inouye, S. A protein Ser/Thr kinase cascade negatively regulates the DNA-binding activity of MrpC, a smaller form of which may be necessary for the Myxococcus xanthus development. Mol. Microbiol. 2006, 60, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.L.; Jeanjean, R.; Liu, Y.D.; Zhang, C.C. pkn22 (alr2502) encoding a putative Ser/Thr kinase in the cyanobacterium Anabaena sp. PCC 7120 is induced by both iron starvation and oxidative stress and regulates the expression of isiA. FEBS Lett. 2003, 553, 179–182. [Google Scholar] [CrossRef]
- Janczarek, M. Environmental signals and regulatory pathways that influence exopolysaccharide production in rhizobia. Int. J. Mol. Sci. 2011, 12, 7898–7933. [Google Scholar] [CrossRef] [PubMed]
- Kusebauch, U.; Ortega, C.; Ollodart, A.; Rogers, R.S.; Sherman, D.R.; Moritz, R.L.; Grundner, C. Mycobacterium tuberculosis supports protein tyrosine phosphorylation. Proc. Natl. Acad. Sci. USA 2014, 111, 9265–9270. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.J.; Deighan, P. Regulation of gene expression by histone-like proteins in bacteria. Curr. Opin. Genet. Dev. 2003, 13, 179–184. [Google Scholar] [CrossRef]
- Aki, T.; Choy, H.E.; Adhya, S. Histone-like protein HU as a specific transcriptional regulator: Co-factor role in repression of gal transcription by GAL repressor. Genes Cells 1996, 1, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Oberto, J.; Nabti, S.; Jooste, V.; Mignot, H.; Rouviere-Yaniv, J. The HU regulon is composed of genes responding to anaerobiosis, acid stress, high osmolarity and SOS induction. PLoS ONE 2009, 4, e4367. [Google Scholar] [CrossRef] [PubMed]
- Sajid, A.; Arora, G.; Gupta, M.; Singhal, A.; Chakraborty, K.; Nandicoori, V.K.; Singh, Y. Interaction of Mycobacterium tuberculosis elongation factor Tu with GTP is regulated by phosphorylation. J. Bacteriol. 2011, 193, 5347–5358. [Google Scholar] [CrossRef] [PubMed]
- Marrakchi, H.; Lanéelle, M.A.; Daffé, M. Mycolic acids: Structures, biosynthesis, and beyond. Chem. Biol. 2014, 21, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Manuse, S.; Fleurie, A.; Zucchini, L.; Lesterlin, C.; Grangeasse, C. Role of eukaryotic-like serine/threonine kinases in bacterial cell division and morphogenesis. FEMS Microbiol. Rev. 2016, 40, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Squeglia, F.; Marchetti, R.; Ruggiero, A.; Lanzetta, R.; Marasco, D.; Dworkin, J.; Petoukhov, M.; Molinaro, A.; Berisio, R.; Silipo, A. Chemical basis of peptidoglycan discrimination by PrkC, a key kinase involved in bacterial resuscitation from dormancy. J. Am. Chem. Soc. 2011, 133, 20676–20679. [Google Scholar] [CrossRef] [PubMed]
- Foulquier, E.; Pompeo, F.; Freton, C.; Cordier, B.; Grangeasse, C.; Galinier, A. PrkC-mediated phosphorylation of overexpressed YvcK protein regulates PBP1 protein localization in Bacillus subtilis mreB mutant cells. J. Biol. Chem. 2014, 289, 23662–23669. [Google Scholar] [CrossRef] [PubMed]
- Soufi, B.; Kumar, C.; Gnad, F.; Mann, M.; Mijakovic, I.; Macek, B. Stable isotope labeling by amino acids in cell culture (SILAC) applied to quantitative proteomics of Bacillus subtilis. J. Proteome Res. 2010, 9, 3638–3646. [Google Scholar] [CrossRef] [PubMed]
- Kamei, A.; Yuasa, T.; Geng, X.; Ikeuchi, M. Biochemical examination of the potential eukaryotic-type protein kinase genes in the complete genome of the unicellular cyanobacterium Synechocystis sp. PCC 6803. DNA Res. 2002, 9, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, F.; Guan, X.; Yang, Y.; Liang, C.; Qin, S. Genome-wide survey of putative serine/threonine protein kinases in cyanobacteria. BMC Genomics 2007, 30, 395. [Google Scholar] [CrossRef] [PubMed]
- Kamei, A.; Yoshihara, S.; Yuasa, T.; Geng, X.; Ikeuchi, M. Biochemical and functional characterization of a eukaryotic-type protein kinase, SpkB, in the cyanobacterium, Synechocystis sp. PCC 6803. Curr. Microbiol. 2003, 46, 296–301. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | STK | Substrate | Function | Reference |
|---|---|---|---|---|
| PknA | FadD, FabH, KasA, KasB, MabA | Mycolic acid synthesis | [21,22] | |
| Mycobacterium | GlmU | Cell wall synthesis | [23] | |
| tuberculosis | FtsZ, MurD, Wag31 | Cell division | [24,25,26,27] | |
| PknB | STK, cell signaling | [27] | ||
| EmbR | Arabinan synthesis | [28] | ||
| GroEL1 | Heat shock protein | [29] | ||
| PknB | FadD, KasA, KasB, MabA | Mycolic acid synthesis | [21] | |
| EmbR | Arabinan synthesis | [28] | ||
| GroEL1 | Heat shock protein | [29] | ||
| GlmU, PBPA | Cell wall synthesis | [23,30] | ||
| PknA | STK, cell signaling | [27] | ||
| RshA | Anti-sigma factor, oxidative stress response | [31] | ||
| SigH | Alternative sigma factor, oxidative stress response | [31] | ||
| GarA | Glycogen recycling, TCA cycle | [32] | ||
| PknD | FadD, FabH, KasA, KasB, MabA | Mycolic acid synthesis | [21,22,33] | |
| GarA | Glycogen recycling, TCA cycle | [32] | ||
| GroEL1 | Heat shock protein | [29] | ||
| Mmp17 | Membrane transporter | [34] | ||
| Rv0516c | Anti-anti-sigma factor | [35] | ||
| Rv1747 | ABC transporter | [36] | ||
| PknE | FadD, KasA, KasB, FabH, MabA | Mycolic acid synthesis | [21,22,33] | |
| GarA | Glycogen recycling, TCA cycle | [32] | ||
| GroEL1 | Heat shock protein | [29] | ||
| Rv1747 | ABC transporter | [36] | ||
| PknF | FadD, KasA, KasB, FabH | Mycolic acid synthesis | [21,22,37] | |
| GroEL1 | Heat shock protein | [29] | ||
| GarA | Glycogen recycling, TCA cycle | [32] | ||
| EthR | Antibiotic resistance | [38] | ||
| Rv1747 | ABC transporter | [37] | ||
| PknG | GarA | Glycogen recycling, TCA cycle, virulence | [39] | |
| PknH | FadD, FabH, KasA, KasB | Mycolic acid synthesis | [21,22,33] | |
| GroEL1 | Heat shock protein | [29] | ||
| Rv0681 | TetR family transcription factor | [40] | ||
| EmbR | Arabinan synthesis, cell wall biosynthesis, virulence | [41] | ||
| DosR | TCS response regulator, oxidative stress tolerance, spore dormancy | [42] | ||
| DacB1 | Penicillin-binding protein | [40] | ||
| GarA | Glycogen recycling, TCA cycle | [39] | ||
| PknI | FadD | Mycolic acid synthesis | [21] | |
| PknJ | EmbR | Arabinan synthesis, cell wall biosynthesis | [41] | |
| PepE | Peptidase | [43] | ||
| Mma4 | Mycolic acid synthesis | [43] | ||
| PknK | FadD | Mycolic acid synthesis | [21] | |
| VirS | Transcription factor, stress response | [44] | ||
| PknL | FadD, MabA, KasA, KasB | Mycolic acid synthesis | [21,22] | |
| Rv2175c | DNA-binding protein, cell envelope | [45] | ||
| GroEL1 | Heat shock protein | [29] | ||
| Bacillus subtilis | PrkA | ND | Indirect regulation of transcription factor δK and regulator ScoC, sporulation | [46] |
| PrkC | CpgA | GTPase, peptydoglycan decomposition, late state of ribosome assembly | [47] | |
| AlsD | Α-acetolactase, central metabolism | [48] | ||
| Icd | Central metabolism | [48] | ||
| YvcK GpsB | Sporulation process, cell growth Cell division protein, sporulation | [49] [49] | ||
| YwjH | Transladolase, central metabolism | [48] | ||
| GlnA | Glutamine synthetase, central metabolism | [48] | ||
| Hpr | Phosphotransferase system kinase | [48] | ||
| WalR | Response regulator of TCS WalRK, cell wall metabolism in stationary phase | [11] | ||
| AbrA | Transcriptional regulator, exoprotease production, competence development and sporulation | [50] | ||
| AbrB | Global transcriptional regulator, transition from exponential to stationary growth phase | [50] | ||
| YkwC | Oxidoreductase | [51] | ||
| EF-G, EF-Tu | Elongation factors, protein translation, spore germination and cell growth | [47,49,52,53] | ||
| PrkD (YbdM) | AbrA | Transcriptional regulator, exoprotease production, competence development | [50] | |
| AbrB | Global transcriptional regulator, transition from exponential to stationary growth phase | [49] | ||
| DnaC | Helicase, DNA replication, cell growth | [49] | ||
| YabT | SsbA | DNA recombinase, spore development | [54] | |
| RacA | DNA-related protein, DNA anchoring to the cell pole, sporulation | [49] | ||
| RecA | DNA recombinase, DNA damage repair, sporulation | [49] | ||
| AbrB | Global transcriptional regulator, transition from exponential to stationary growth phase | [49] | ||
| AbrA | Transcriptional regulator, exoprotease production, competence development and sporulation | [49] | ||
| EF-G, EF-Tu | Elongation factors, inhibition of protein translation in spores | [55,56] | ||
| YdiB (Ser/Thr/Tyr) | YdiE | Translation, oxidative stress response | [10] | |
| MBP * | Human myelin basic protein (artificial substrate) | [10] | ||
| Bacillus anthracis | PrkC (BA-Stk1) | ND MBP * | Survival within macrophages, virulence Human myelin basic protein (artificial substrate) | [57] [57] |
| PrkD | BasPyk | Pyruvate kinase phosphorylation, glycolysis, cell growth and development | [58] | |
| MBP * | Human myelin basic protein (artificial substrate) | [58] | ||
| PrkG | MBP * | Human myelin basic protein (artificial substrate) STK PrkG involved in cell growth and development | [58] | |
| Corynebacterium | PknA | MurC | Cell wall biosynthesis | [59] |
| glutamicum | FtsZ | Cell division | [60] | |
| OdhI | Glutamate catabolism | [60,61] | ||
| PknG | Soluble STK | [61] | ||
| PknB | FtsZ | Cell division | [60] | |
| OdhI | Glutamate catabolism | [60,61] | ||
| PknG | OdhI | Glutamate catabolism | [60,61] | |
| PknL | FtsZ | Cell division | [60] | |
| Streptococcus | Stk1 | DivIVA | Cell division | [62] |
| agalactiae | CovR | TCS CovRS response regulator, toxin expression, virulence | [63,64] | |
| (Group B | EF-Tu | Elongation factor | [63,64,65] | |
| Streptococcus) | PpaC | Inorganic pyrophosphatase, virulence | [65] | |
| ND | ATP-dependent DNA i RNA helicases | [66] | ||
| Streptococcus pyogenes | SP-STK | WalR CovR | TCS WalRK response regulator, cell wall TCS CovRS response regulator, virulence | [67] [67] |
| (Group A Streptococcus) | SP-HLP, HU | Histone-like protein | [67] | |
| Streptococcus | StkP | FtsZ | Cell division, cellular morphogenesis | [68] |
| pneumoniae | DivIVA | Cell division, cellular morphogenesis | [69] | |
| PpaC | Inorganic pyrophosphatase, virulence | [68] | ||
| RitR | Transcriptional regulator, iron uptake, oxidative stress response | [70] | ||
| MurC | Cell wall biosynthesis | [71] | ||
| ComD | Competence-specific receptor, TCS ComDE | [72] | ||
| RR06 | Adhesion, virulence | [73] | ||
| RpoA | RNA polymerase α subunit | [74] | ||
| Streptococcus mutans | PknB | ND ND | Cell wall metabolism, bacteriocin production, cell wall metabolism, growth, biofilm formation Regulation of Smu2146c, TCSs VicRK and ComDE, oxidative stress tolerance | [75] |
| Staphylococcus | Stk1 | SA0498 | Ribosomal protein L7/L12 | [76] |
| aureus | (PknB) | SA0545 | Phosphate acetyltransferase | [76] |
| (secreted) | SA0731 | Enolase | [76] | |
| SA1359 | Elongation factor P | [76] | ||
| SA2340 | Glyoxalase | [76] | ||
| SA2399 | Fructose biphosphate aldolase | [76] | ||
| MgrA | Global transcriptional regulator, antibiotic resistance | [77] | ||
| SarA | Global transcriptional regulator, virulence | [78] | ||
| SarZ | Oxidative stress response | [79] | ||
| PurA | Purine biosynthesis | [77,78] | ||
| HU | DNA-binding histone-like protein | [77,78] | ||
| CcpA | Catabolite control protein A, carbon metabolism, virulence | [80] | ||
| VraR | Vancomycin-resistance-associated response regulator | [81] | ||
| GraR | TCS GraSR response regulator, antibiotic resistance | [82] | ||
| SpoVG | Transcriptional factor, virulence, antibiotic resistance | [83] | ||
| Staphylococcus epidermidis | Stk | ND | Polysaccharide intercellular adhesin (PIA) production, biofilm formation, virulence | [84] |
| Mycoplasma pneumoniae | PrkC | ND | Surface protein (adhesin P1), HmW1-3, and MPN474 phosphorylation, adhesion | [85] |
| Mycoplasma genitalium | MG_109 | ND | Virulence | [86] |
| Enterococcus faecium | Stk | P5AP | Cell signaling, antibiotic resistance, peptydoglycan biosynthesis | [87,88] |
| Myxococcus | Pkn2 | HU | Histone-like protein | [89] |
| xanthus | Pkn4 | PFK | Glycolysis | [90] |
| Pkn5 | ND | Soluble STK, cell growth and development, formation of fruiting bodies | [91] | |
| Pkn6 | ND | Transmembrane STK, cell growth and development, formation of fruiting bodies | [91] | |
| Pkn8 | Pkn14 | Soluble STK | [92] | |
| MrpC | Transcription factor, development of fruiting bodies | [92] | ||
| Pkn14 | MrpC | Transcription factor, development of fruiting bodies | [92] | |
| Synechocystis sp. PCC 6803 | SpkA | ND | PilA1, A2, A5, A6, A9, A10 expression, cell motility | [93,94] |
| MBP *, casein *, histone * | Artificial substrates | [93,94] | ||
| SpkB | GlyS | Glycyl-tRNA synthetase β-subunit, oxidative stress adaptation | [95] | |
| SpkC | SpkK | Soluble STK, stress response | [96] | |
| SpkD | ND | Carbon metabolism, TCA cycle regulation, bacterial growth | [97] | |
| SpkE | ND | Cell signaling | [98] | |
| SpkF | SpkC | Membrane-associated STK, stress response | [96] | |
| SpkG | ND | High salt resistance, stress-mediated signaling | [99] | |
| SpkK | GroES | Small co-chaperonin | [96] | |
| Anabaena sp. | PknA | ND | Optimal growth | [100] |
| PknC | ND | Optimal growth | [100] | |
| PknD | ND | Optimal growth, heterocyst functioning, nitrogen fixation | [101,102,103] | |
| PCC 7120 | PknE | ND | Optimal diazotrophic growth, heterocyst differentiation, nitrogen fixation | [101,102,103] |
| PknH | ND | Diazotrophic growth, maintaining connections between heterocysts and vegetative cells | [104] | |
| L. | PrkA | PolC | DNA Polymerase III α subunit | [105] |
| monocytogenes | RpoA | RNA polymerase α subunit | [105] | |
| RpoB | RNA polymerase β subunit | [105] | ||
| RecA | Recombinant protein | [105] | ||
| EF-Tu, EF-G | Translation elongation factors | [105] | ||
| Yvck | Cell wall homeostasis, glycerol metabolism, cytosolic survival, virulence | [106] | ||
| Pseudomonas aeruginosa | PpkA | Fha1 | FHA domain-containing protein, hemolysin-coregulated protein 1 (Hcp1) secretion, biofilm formation, virulence, stress tolerance | [107] |
| H1 * | Eukaryotic histone H1 | [108] | ||
| Yersinia pseudotuberculosis | YpkA (secreted) | ND | Virulence factor activated by host cell actin, cytoskeleton disruption, inhibition of macrophage function | [109] |
| E. coli (EHEC) | Stk | ND | Virulence | [110] |
| Rhizobium leguminosarum | BAE36_06965 BAE36_16215 BAE36_31125 | ND | Optimal cell growth, oxidative stress adaptation | [111] |
| Species | STP | Type | Partner kinase | Substrate | Function | References |
|---|---|---|---|---|---|---|
| Mycobacterium tuberculosis | PstP | PPM | PknB | PknA | STK, cell signaling, cell growth and division, cell survival | [136,137] |
| PknB | STK, cell signaling, cell growth | [137] | ||||
| PknH | STK, cell signaling, cell growth | [136] | ||||
| PknJ | STK, cell signaling, cell growth | [138] | ||||
| PykA | Pyruvate kinase, glycolysis | [138] | ||||
| EmbR | Transcriptional regulator of embCAB operon | [136] | ||||
| ND | S-adenosylhomocysteine hydrolase, homocysteine metabolism | [139] | ||||
| PapA5 | Cell wall metabolism | [140] | ||||
| Rv0019c | FHA-domain-containing protein interacting with FtsZ, GtsQ, and PapA5, cell division | [140] | ||||
| PBPA | PBP, cell wall biosynthesis | [30] | ||||
| Bacillus | PrpC | PP2C | PrkC | EF-G, EF-Tu | Translation factor | [47,52] |
| subtilis | PrkC | STK, cell signaling, biofilm formation, sporulation | [47,52] | |||
| CpG | Ribosome-associated GTPase | [47] | ||||
| YezB | Stress response | [47] | ||||
| HPr kinase | Phosphotransferase system | [141] | ||||
| RsbX | PPM | RsbB | RsbS, TsbR | Sigma B regulation, stress response | [142] | |
| RsbU | PPM | RsbV | RsbV | Sigma B regulation, stress response | [142] | |
| RsbP | PPM | RsbV | Energy stress response | [142] | ||
| SpoIIE | PPM | SpoIIAA | Anti-anti-sigma factor, sporulation | [143] | ||
| Bacillus anthracis | PrpC (BA-Stp1) | PP2C | BA-Stk1 | PrkC (BA-Stk1) | STK, cell signaling, survival within macrophages, virulence | [144] |
| PrkD PrkG | Dual-specificity tyrosine phosphorylation-regulated kinases | [58] | ||||
| Streptococcus | Stp1 | PP2C | Stk1 | Stk1 | STK, cell signaling, cell aggregation | [65] |
| agalactiae | P35 | Mn2+-dependent inorganic pyrophosphatase | [65] | |||
| (group B | PpaC | Inorganic pyrophosphatase | [65] | |||
| Streptococcus) | PurA ND | Purine synthesis Hemolysin activity, autolysis, virulence | [65] [65] | |||
| Streptococcus | PhpP | PP2C | StkP | StkP | STK, cell signaling | [74] |
| pneumoniae | RitR | Transcriptional regulator | [70] | |||
| ComD | Competence-specific receptor, TCS ComDE system | [72] | ||||
| MurC | Peptydoglycan synthesis | [71] | ||||
| MapZ | Cell division | [145] | ||||
| RR06 | TCS response regulator | [67,146] | ||||
| DivIVA | Cell division | [147] | ||||
| Streptococcus | SP-STP | PP2C | SP-STK | SP-STK | STK, cell signaling | [67,148] |
| pyogenes | (secreted) | SP-HLP ND | Histone-like protein Induction of host cell apoptosis | [148] | ||
| Streptococcus mutans | PppL | PP2C | PknB | STK, cell signaling | [149] | |
| Staphylococcus aureus | Stp1 | PP2C | Stk1 | Stk1 | STK, cell signaling, membrane integrity, cell division, cell wall biosynthesis | [150,151,152] |
| GraR | TCS response regulator | [150,151,152] | ||||
| MgrA | Global transcriptional regulator | [79] | ||||
| Myxococcus xanthus | Pph1 | PP2C | Pkn5 | Pkn5 | Negative effector of development, vegetative growth and formation of fruiting bodies | [153] |
| Enterococcus faecium | StpA | PP2C | Stk | Stk P5AP | STK, cell signaling, antibiotic resistance, peptydoglycan biosynthesis | [85] |
| MBP * | Myelin basic protein (artificial substrate) | [85] | ||||
| Mycoplasma pneumoniae | PrpC | PP2C | HPr | Phosphocarrier protein, phosphotransferase system | [154] | |
| Mycoplasma synoviae | PrpC | PP2C | ND Phosphopeptides * | Cell signaling Artificial substrates | [155] | |
| Mycoplasma genitalium | MG_207 | PPM | MG_109 | ND | Cell signaling, virulence | [156] |
| Listeria | Stp | PP2C | EF-Tu | Translation factor, protein synthesis regulation | [157] | |
| monocytogenes | SOD | Superoxide dismutase, respiratory metabolism | [158] | |||
| Pseudomonas aeruginosa | PppA | PP2C | PpkA | FHA-1 | FHA-domain-containing protein, hemolysin-coregulated protein 1 (Hcp1) secretion | [105] |
| TpbA | PPP | TpbB | Dual-specificity Ser/Thr/Tyr kinase, cell motility, biofilm formation | [159] | ||
| Stp1 | PP2C | ND | Protein synthesis | [105,159] | ||
| Escherichia coli | PrpA | PPP | ND Casein, MBP * | Signaling protein misfolding via TCS CpxRA, heat shock response Dual-specificity Ser/Thr/Tyr phosphatase | [160] | |
| Salmonella enterica ser. Typhi | PrpZ (Ser/Thr/Tyr) | PP2C | MBP * | Myelin basic protein | [161] | |
| Synechocystis sp. PCC 6803 | IcfG | PP2C | Slr1856 | Carbon metabolism | [162] | |
| PphA | PPM | PII | Nitrogen assimilation | [163] | ||
| Anabaena sp. PCC7120 | All1758 | PP2C | ND | Diazotrophic growth, cell morphology, glycolipid synthesis | [164] | |
| Rhizobium leguminosarum | PssZ | PP2C | ND | Cell envelope biogenesis, stress response, motility | [165] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janczarek, M.; Vinardell, J.-M.; Lipa, P.; Karaś, M. Hanks-Type Serine/Threonine Protein Kinases and Phosphatases in Bacteria: Roles in Signaling and Adaptation to Various Environments. Int. J. Mol. Sci. 2018, 19, 2872. https://doi.org/10.3390/ijms19102872
Janczarek M, Vinardell J-M, Lipa P, Karaś M. Hanks-Type Serine/Threonine Protein Kinases and Phosphatases in Bacteria: Roles in Signaling and Adaptation to Various Environments. International Journal of Molecular Sciences. 2018; 19(10):2872. https://doi.org/10.3390/ijms19102872
Chicago/Turabian StyleJanczarek, Monika, José-María Vinardell, Paulina Lipa, and Magdalena Karaś. 2018. "Hanks-Type Serine/Threonine Protein Kinases and Phosphatases in Bacteria: Roles in Signaling and Adaptation to Various Environments" International Journal of Molecular Sciences 19, no. 10: 2872. https://doi.org/10.3390/ijms19102872