Dissecting Time- from Tumor-Related Gene Expression Variability in Bilateral Breast Cancer

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
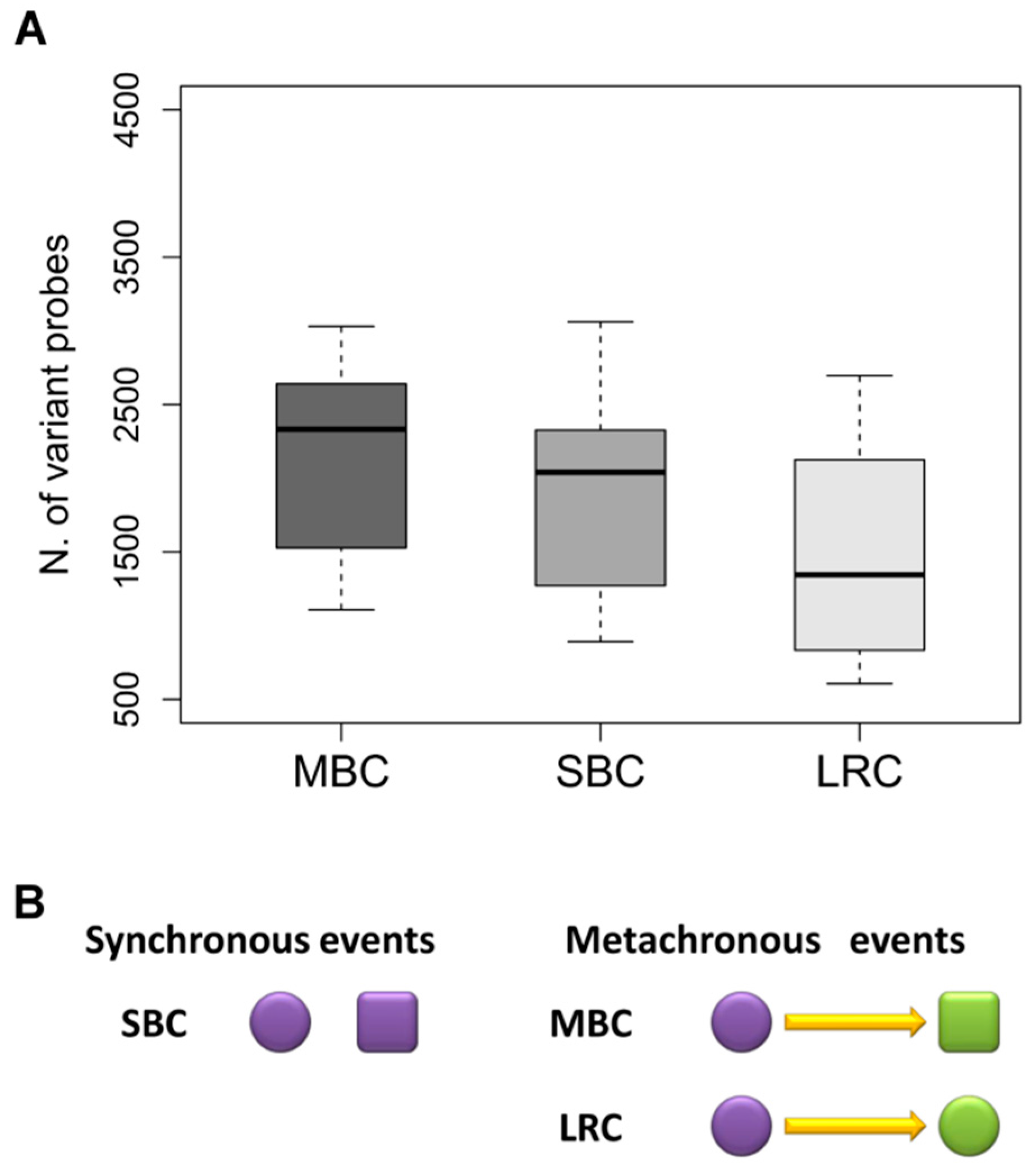
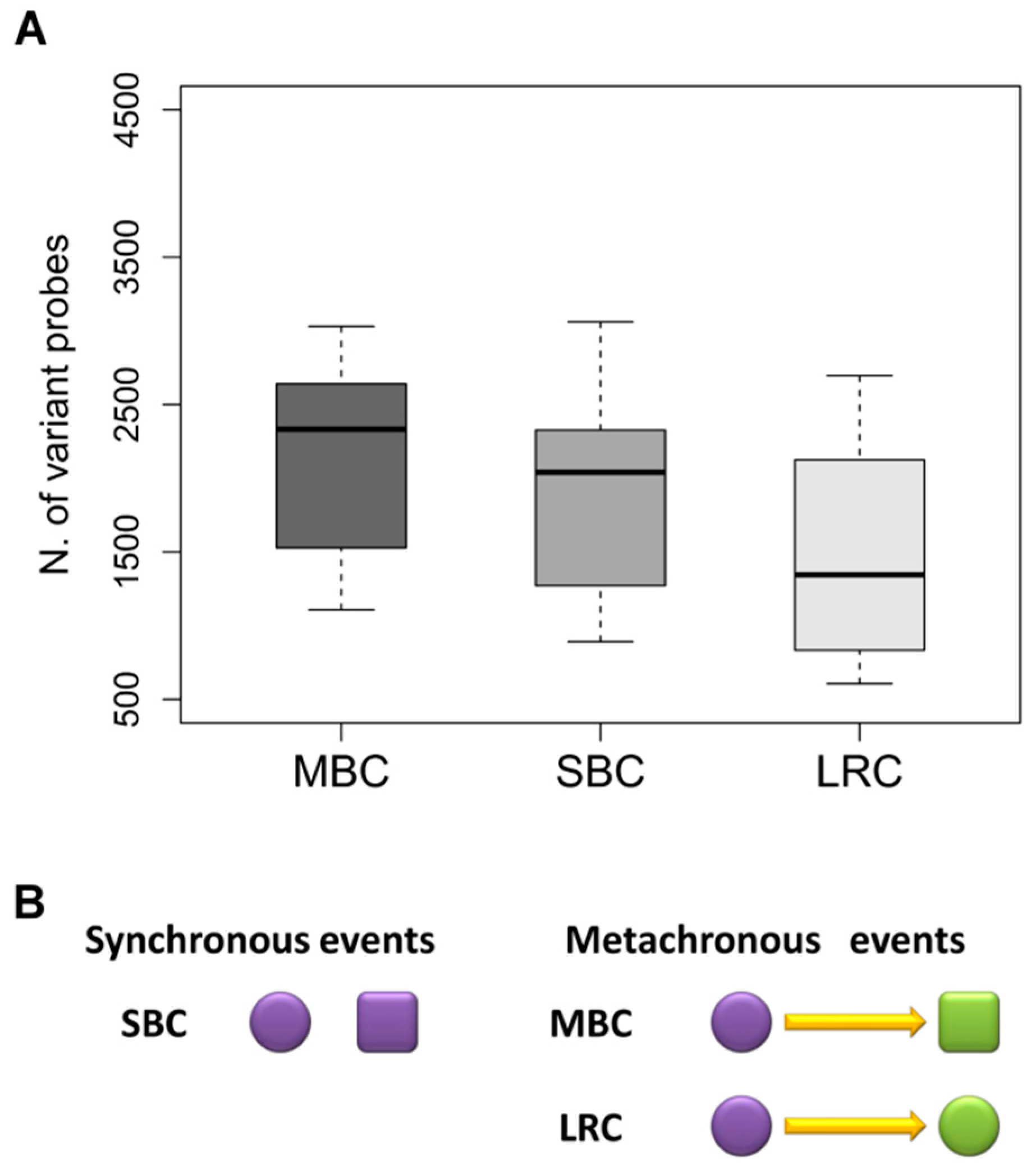
2.1. Patient and Tumor Characteristics
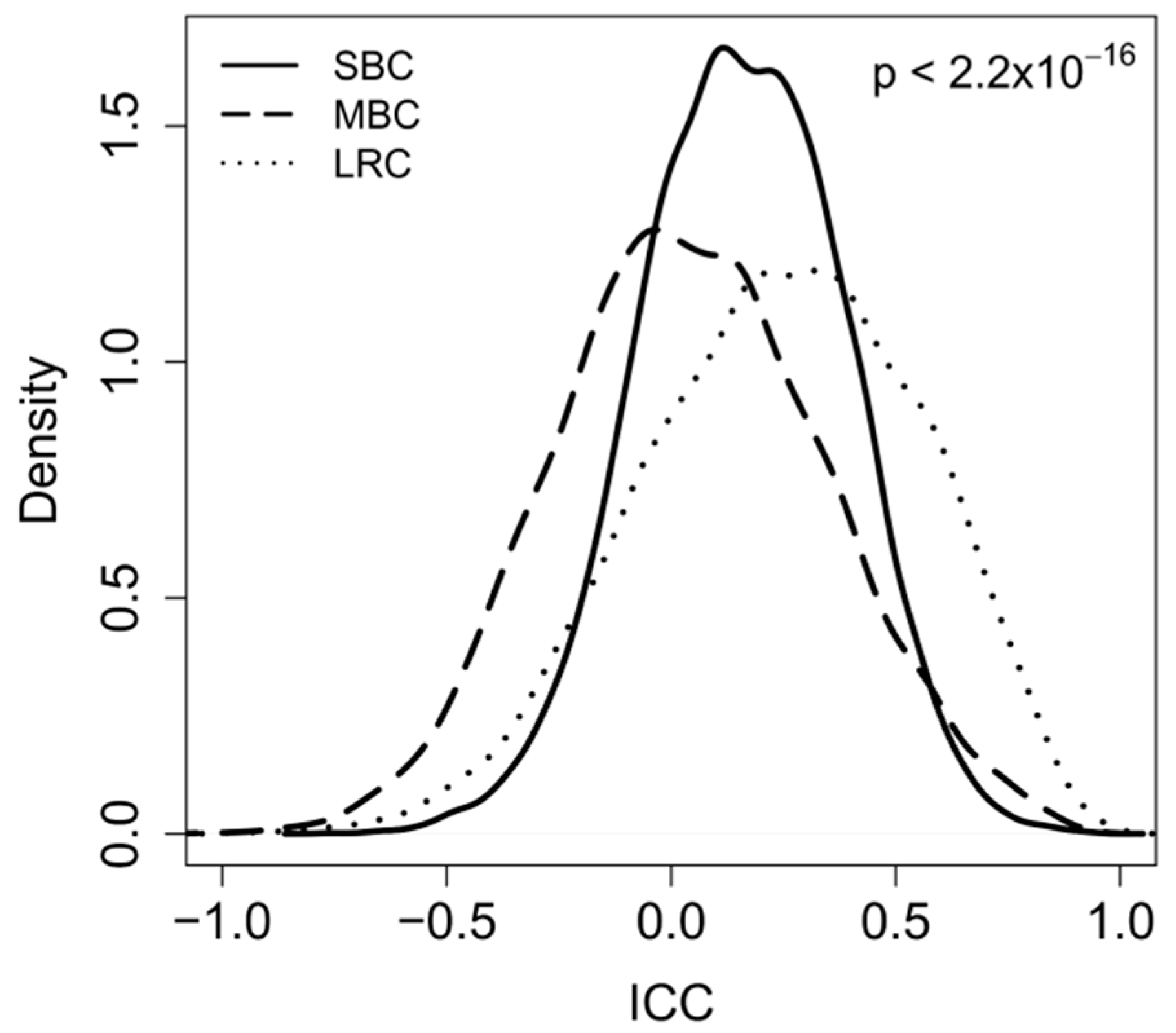
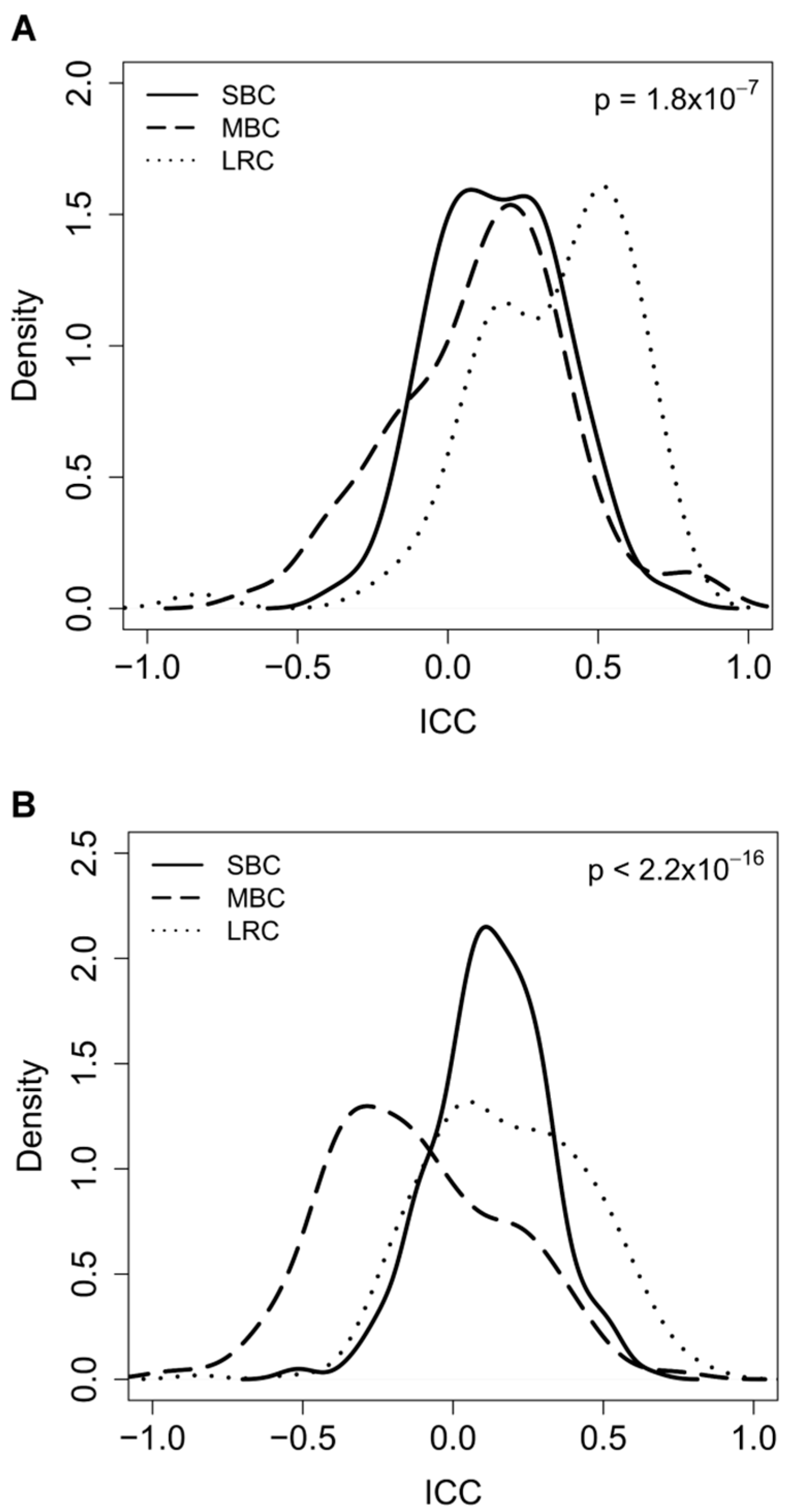
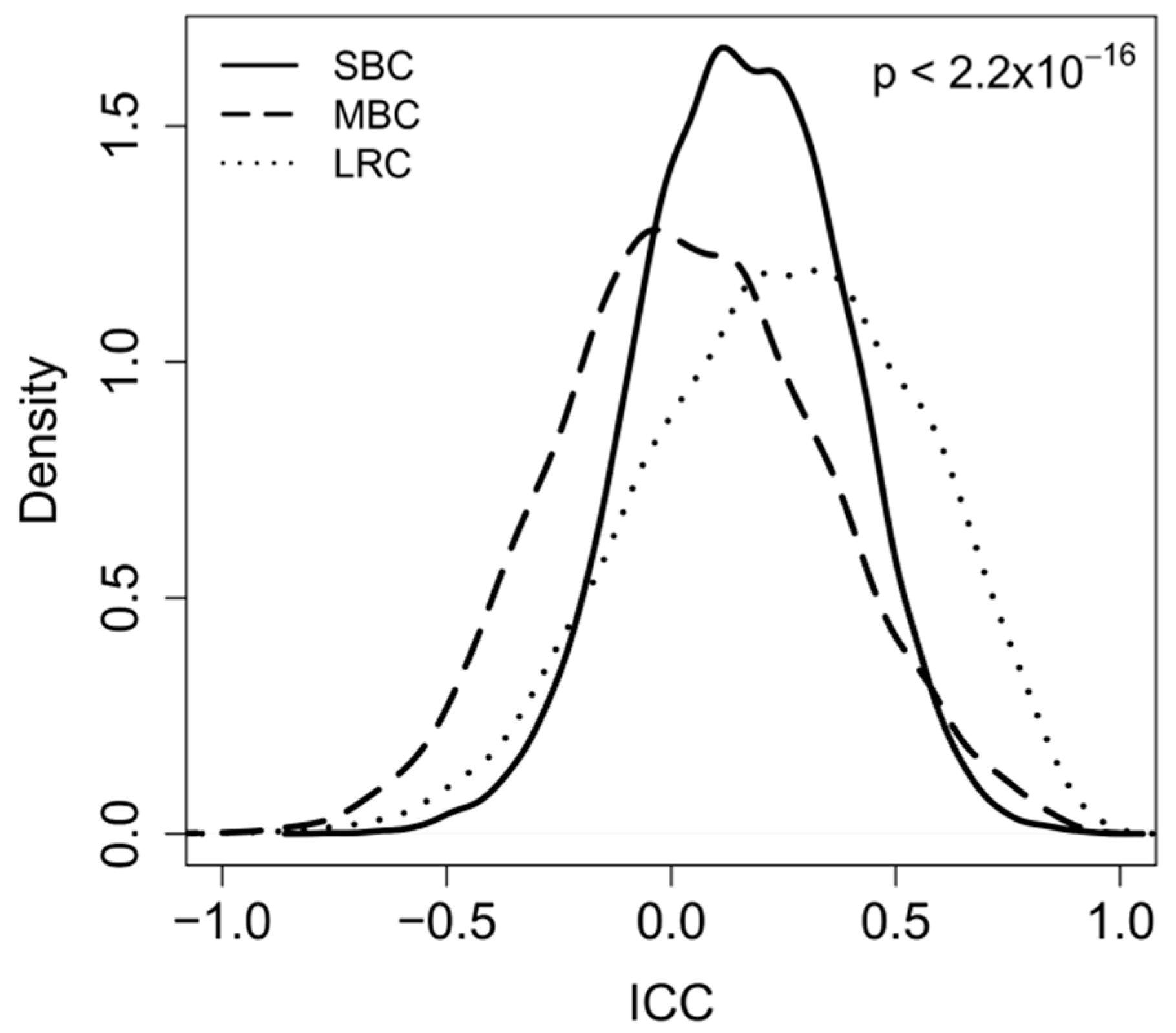
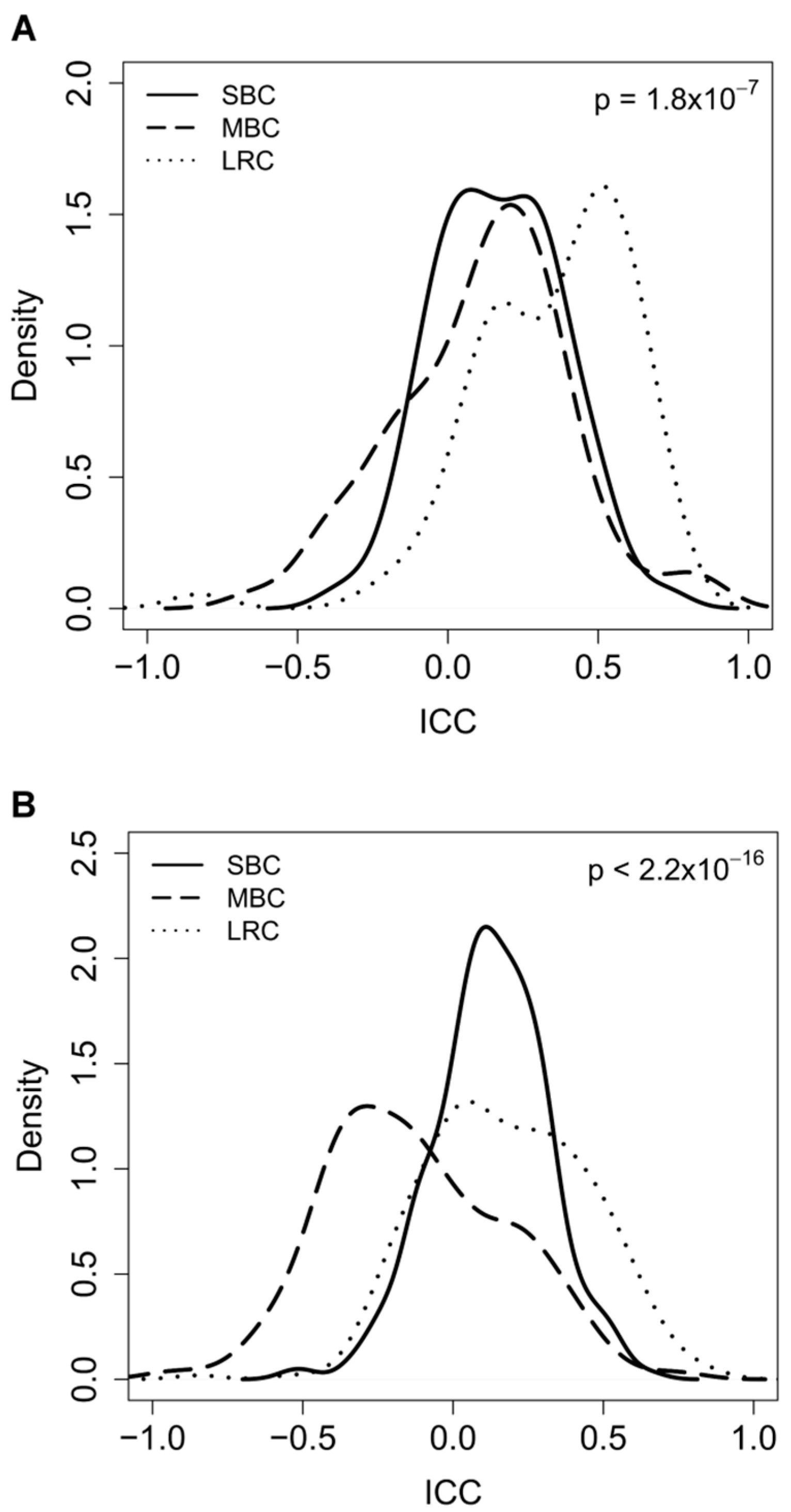
2.2. Correlation between Paired-Samples by Gene Expression Data
2.3. Dissecting Intra-Pair Variability with the Aid Microenvironment-Related and Tumor-Intrinsic Gene Signatures
3. Material and Methods
3.1. Case Series
3.2. RNA Extraction and Gene Expression Profiling
3.3. Data Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harris, L.N.; Ismaila, N.; McShane, L.M.; Andre, F.; Collyar, D.E.; Gonzalez-Angulo, A.M.; Hammond, E.H.; Kuderer, N.M.; Liu, M.C.; Mennel, R.G.; et al. Use of Biomarkers to Guide Decisions on Adjuvant Systemic Therapy for Women With Early-Stage Invasive Breast Cancer: American Society of Clinical Oncology Clinical Practice Guideline. J. Clin. Oncol. 2016, 34, 1134–1150. [Google Scholar] [CrossRef] [PubMed]
- Narod, S.A. Bilateral breast cancers. Nat. Rev. Clin. Oncol. 2014, 11, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Van der Leij, F.; Elkhuizen, P.H.; Bartelink, H.; van de Vijver, M.J. Predictive factors for local recurrence in breast cancer. Semin. Radiat. Oncol. 2012, 22, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, R.A.; Murad, T.; Hines, J.R. Bilareal breast cancer. Am. Surg. 1989, 55, 441–444. [Google Scholar] [PubMed]
- Kurtz, M.; Amalric, R.; Brandone, H.; Ayme, Y.; Spitalier, J.M. Contralateral breast cancer and other second malignancies in patients treated by breast-conserving therapy with radiation. Int. J. Radiat. Oncol. Biol. Phys. 1988, 15, 277–284. [Google Scholar] [CrossRef]
- Harvey, E.B.; Brinton, L.A. Second cancer following cancer of the breast in Connecticut, 1935–82. Nat. Cancer Inst. Monogr. 1985, 68, 99–112. [Google Scholar] [PubMed]
- Rasmussen, C.B.; Kjær, S.K.; Ejlertsen, B.; Andersson, M.; Jensen, M.B.; Christensen, J.; Langballe, R.; Mellemkjær, L. Incidence of metachronous contralateral breast cancer in Denmark 1978–2009. Int. J. Epidemiol. 2014, 43, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Banelli, B.; Casciano, I.; Di Vinci, A.; Gatteschi, B.; Levaggi, A.; Carli, F.; Bighin, C.; Salvi, S.; Allemanni, G.; Ghiorzo, P.; et al. Pathological and molecular characteristics distinguishing contralateral metastatic from new primary breast cancer. Ann. Oncol. 2010, 21, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Chaudary, M.A.; Millis, R.R.; Hoskins, E.O.; Halder, M.; Bulbrook, R.D.; Cuzick, J.; Hayward, J.L. Bilateral primary breast cancer: A prospective study of disease incidence. Br. J. Surg. 1984, 71, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Rubino, C.; Arriagada, R.; Delaloge, S.; Lê, M.G. Relation of risk of contralateral breast cancer to the interval since the first primary tumour. Br. J. Cancer 2010, 102, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Alkner, S.; Tang, M.H.; Brueffer, C.; Dahlgren, M.; Chen, Y.; Olsson, E.; Winter, C.; Baker, S.; Ehinger, A.; Rydén, L.; et al. Contralateral breast cancer can represent a metastatic spread of the first primary tumor: Determination of clonal relationship between contralateral breast cancers using next-generation whole genome sequencing. Breast Cancer Res. 2015, 17, 102. [Google Scholar] [CrossRef] [PubMed]
- Klevebring, D.; Lindberg, J.; Rockberg, J.; Hilliges, C.; Hall, P.; Sandberg, M.; Czene, K. Exome sequencing of contralateral breast cancer identifies metastatic disease. Breast Cancer Res. Treat. 2015, 151, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Messer, K.; Schwab, R.; Harismendy, O.; Pu, M.; Crain, B.; Yost, S.; Frazer, K.A.; Rana, B.; Hasteh, F.; et al. Mutational Profiling Can Establish Clonal or Independent Origin in Synchronous Bilateral Breast and Other Tumors. PLoS ONE 2015, 10, e0142487. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, X.; Song, F.; Zhao, Y.; Li, H.; Zheng, H.; Gao, Z.; Wang, J.; Zhang, W.; Chen, K. Comparative genomic analysis reveals bilateral breast cancers are genetically independent. Oncotarget 2015, 6, 31820–31829. [Google Scholar] [CrossRef] [PubMed]
- Begg, C.B.; Ostrovnaya, I.; Geyer, F.C.; Papanastasiou, A.D.; Ng, C.K.Y.; Sakr, R.A.; Bernstein, J.L.; Burke, K.A.; King, T.A.; Piscuoglio, S.; et al. Contralateral breast cancers: Independent cancers or metastases? Int. J. Cancer 2017, 142, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.; Anderson, S.; Bryant, J.; Margolese, R.G.; Deutsch, M.; Fisher, E.R.; Jeong, J.H.; Wolmark, N. Twenty-year follow-up of a randomized trial comparing total mastectomy, lumpectomy, and lumpectomy plus irradiation for the treatment of invasive breast cancer. N. Engl. J. Med. 2002, 347, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.A.; Voogd, A.C.; Fentiman, I.S.; Legrand, C.; Sylvester, R.J.; Tong, D.; van der Schueren, E.; Helle, P.A.; van Zijl, K.; Bartelink, H. Long-term results of a randomized trial comparing breast-conserving therapy with mastectomy: European Organization for Research and Treatment of Cancer 10,801 trial. J. Natl. Cancer Inst. 2000, 92, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, U.; Cascinelli, N.; Mariani, L.; Greco, M.; Saccozzi, R.; Luini, A.; Aguilar, M.; Marubini, E. Twenty-year follow-up of a randomized study comparing breast-conserving surgery with radical mastectomy for early breast cancer. N. Engl. J. Med. 2002, 347, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Oskarsson, T.; Acharyya, S.; Nguyen, D.X.; Zhang, X.H.; Norton, L.; Massagué, J. Tumor self-seeding by circulating cancer cells. Cell 2009, 139, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Comen, E.; Norton, L. Self-seeding in cancer. Recent Results Cancer Res. 2012, 195, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Callari, M.; Musella, V.; Di Buduo, E.; Sensi, M.; Miodini, P.; Dugo, M.; Orlandi, R.; Agresti, R.; Paolini, B.; Carcangiu, M.L.; et al. Subtype-dependent prognostic relevance of an interferon-induced pathway metagene in node-negative breast cancer. Mol. Oncol. 2014, 8, 1278–1289. [Google Scholar] [CrossRef] [PubMed]
- Shrout, P.E.; Fleiss, J.L. Intraclass correlations: Uses in assessing rater reliability. Psychol. Bull. 1979, 86, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Koo, T.K.; Li, M.Y. A Guideline of Selecting and Reporting Intraclass Correlation Coefficients for Reliability Research. J. Chiropr. Med. 2016, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sorlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.S.; Mullins, M.; Cheang, M.C.; Leung, S.; Voduc, D.; Vickery, T.; Davies, S.; Fauron, C.; He, X.; Hu, Z.; et al. Supervised risk predictor of breast cancer based on intrinsic subtypes. J. Clin. Oncol. 2009, 27, 1160–1167. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, G.; Qi, Y.; Alvarez, R.H.; Iwamoto, T.; Coutant, C.; Ibrahim, N.K.; Valero, V.; Cristofanilli, M.; Green, M.C.; Radvanyi, L.; et al. Molecular anatomy of breast cancer stroma and its prognostic value in estrogen receptor-positive and-negative cancers. J. Clin. Oncol. 2010, 28, 4316–4323. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA integrity number for assigning integrity values to RNA measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Zeisberg, M. Fibroblasts in cancer. Nat. Rev. Cancer 2006, 6, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Merlino, G.; Miodini, P.; Paolini, B.; Carcangiu, M.L.; Gennaro, M.; Dugo, M.; Daidone, M.G.; Cappelletti, V. Stromal Activation by Tumor Cells: An in Vitro Study in Breast Cancer. Microarrays (Basel) 2016, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, B.; Fumagalli, D.; Brohee, S.; Hatse, S.; Govaere, O.; Floris, G.; Van den Eynde, K.; Bareche, Y.; Schöffski, P.; Smeets, A.; et al. The footprint of the ageing stroma in older patients with breast cancer. Breast Cancer Res. 2017, 19, 78. [Google Scholar] [CrossRef] [PubMed]
- Merlino, G.; Miodini, P.; Callari, M.; D’Aiuto, F.; Cappelletti, V.; Daidone, M.G. Prognostic and functional role of subtype-specific tumor–stroma interaction in breast cancer. Mol. Oncol. 2017, 11, 1399–1412. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| SBC (n = 18 Pairs) | MBC (n = 11 Pairs) | LRC (n = 10 Pairs) | ||||
|---|---|---|---|---|---|---|
| Age (years) | ||||||
| Range | 29–82 | 38–68 | 30–70 | |||
| Median | 63.5 | 57 | 54.5 | |||
| ≤50 | 2 (11.1%) | 5 (45.5%) | 5 (50%) | |||
| >50 | 16 (88.9%) | 6 (54.5%) | 5 (50%) | |||
| Size (cm) | Right | Left | Primary | Secondary | Primary | Secondary |
| Range | 1–8.5 | 1.2–3.1 | 1.3–3.7 | 0.4–3 | 0.7–2.8 | 0.5–3.4 |
| Median | 2.25 | 2 | 1.8 | 1.7 | 1.75 | 1.6 |
| ≤2 | 10 (55.6%) | 11 (61.1%) | 8 (72.7%) | 8 (72.7%) | 6 (60%) | 6 (60%) |
| >2 | 8 (44.4%) | 7 (38.9%) | 3 (27.3%) | 3 (27.3%) | 4 (40%) | 4 (40%) |
| ER | Right | Left | Primary | Secondary | Primary | Secondary |
| positive | 12 (66.7%) | 15 (83.3%) | 10 (90.9%) | 10 (90.9%) | 10 (100%) | 8 (80%) |
| negative | 6 (33.3%) | 3 (16.7%) | 1 (9.1%) | 1 (9.1%) | 0 | 2 (20%) |
| ERBB2 | Right | Left | Primary | Secondary | Primary | Secondary |
| positive | 1 (5.6%) | 2 (11.1%) | 1 (9.1%) | 1 (9.1%) | 1 (10%) | 2(20%) |
| negative | 17 (94.4%) | 16 (88.9%) | 10 (90.9%) | 10 (90.9%) | 9 (90%) | 8 (80%) |
| Time to 2nd event (months) | ||||||
| Range | - | 9–149 | 42–151 | |||
| Median | - | 76 | 74 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callari, M.; Dugo, M.; Miodini, P.; Veneroni, S.; Bianchini, G.; Daidone, M.G.; Cappelletti, V. Dissecting Time- from Tumor-Related Gene Expression Variability in Bilateral Breast Cancer. Int. J. Mol. Sci. 2018, 19, 196. https://doi.org/10.3390/ijms19010196
Callari M, Dugo M, Miodini P, Veneroni S, Bianchini G, Daidone MG, Cappelletti V. Dissecting Time- from Tumor-Related Gene Expression Variability in Bilateral Breast Cancer. International Journal of Molecular Sciences. 2018; 19(1):196. https://doi.org/10.3390/ijms19010196
Chicago/Turabian StyleCallari, Maurizio, Matteo Dugo, Patrizia Miodini, Silvia Veneroni, Giampaolo Bianchini, Maria Grazia Daidone, and Vera Cappelletti. 2018. "Dissecting Time- from Tumor-Related Gene Expression Variability in Bilateral Breast Cancer" International Journal of Molecular Sciences 19, no. 1: 196. https://doi.org/10.3390/ijms19010196