
The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases

 and
and
Abstract
:
1. Introduction
2. PON1 and Nutrigenetics
3. Plant Preparations
4. Pomegranate Juice
5. Phenolic Compounds
6. Vitamins
7. Protein and Amino Acids
8. Dietary Lipids
8.1. Cholesterol Supplementation
8.2. Polyunsaturated Fatty Acids
Fish Oil
8.3. Olive Oil and Coenzyme Q10
8.4. Hydrocarbons
9. Oligoelement Supplementation
10. Others
11. Conditions Negatively Affecting PON1 Activity
12. PON2
13. PON3
14. Conclusions and Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Kingwell, B.A.; Chapman, M.J.; Kontush, A.; Miller, N.E. HDL-targeted therapies: Progress, failures and future. Nat. Rev. Drug Discov. 2014, 13, 445–464. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Rosenblat, M.; Bisgaier, C.L.; Newton, R.S.; Primo-Parmo, S.L.; La Du, B.N. Paraoxonase inhibits high-density lipoprotein oxidation and preserves its functions. A possible peroxidative role for paraoxonase. J. Clin. Investig. 1998, 101, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.I.; Durrington, P.N.; Mackness, B. How high-density lipoprotein protects against the effects of lipid peroxidation. Curr. Opin. Lipidol. 2000, 11, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Kriska, T.; Marathe, G.K.; Schmidt, J.C.; McIntyre, T.M.; Girotti, A.W. Phospholipase action of platelet-activating factor acetylhydrolase, but not paraoxonase-1, on long fatty acyl chain phospholipid hydroperoxides. J. Biol. Chem. 2007, 282, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Biol. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Gaidukov, L.; Khersonsky, O.; Vaya, J.; Oren, R.; Tawfik, D.S.; Aviram, M. The catalytic histidine dyad of high density lipoprotein-associated serum paraoxonase-1 (PON1) is essential for PON1-mediated inhibition of low density lipoprotein oxidation and stimulation of macrophage cholesterol efflux. J. Biol. Chem. 2006, 281, 7657–7665. [Google Scholar] [CrossRef] [PubMed]
- Ben-David, M.; Elias, M.; Filippi, J.-J.; Duñach, E.; Silman, I.; Sussman, J.L.; Tawfik, D.S. Catalytic versatility and backups in enzyme active sites: The case of serum paraoxonase 1. J. Mol. Biol. 2012, 418, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Poeckel, D.; Funk, C.D. The 5-lipoxygenase/leukotriene pathway in preclinical models of cardiovascular disease. Cardiovasc. Res 2010, 86, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Fuhrman, B.; Volkova, N.; Aviram, M. Postprandial serum triacylglycerols and oxidative stress in mice after consumption of fish oil, soy oil or olive oil: Possible role for paraoxonase-1 triacylglycerol lipase-like activity. Nutrition 2006, 22, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.; Durrington, P.; McElduff, P.; Yarnell, J.; Azam, N.; Watt, M.; Mackness, M. Low paraoxonase activity predicts coronary events in the caerphilly prospective study. Circulation 2003, 107, 2775–2779. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.; Mackness, B. Human paraoxonase-1 (PON1): Gene structure and expression, promiscuous activities and multiple physiological roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.M.; Gu, L.; Xia, Y.-R.; Navab, M.; Li, W.-F.; Hama, S.; Castellani, L.W.; Furlong, C.E.; Costa, L.G.; Fogelman, A.M.; et al. Mice lacking serum paraoxonase are susceptible to organophosphate toxicity and atherosclerosis. Nature 1998, 394, 284–287. [Google Scholar] [PubMed]
- Tward, A.; Xia, Y.-R.; Wang, X.-P.; Shi, Y.-S.; Park, C.; Castellani, L.W.; Lusis, A.J.; Shih, D.M. Decreased atherosclerotic lesion formation in human serum paraoxonase transgenic mice. Circulation 2002, 106, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Giordano, G.; Furlong, C.E. Pharmacological and dietary modulators of paraoxonase 1 (PON1) activity and expression: The hunt goes on. Biochem. Pharmacol. 2011, 81, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Lou-Bonafonte, J.; Gabás-Rivera, C.; Navarro, M.; Osada, J. PON1 and mediterranean diet. Nutrients 2015, 7, 4068–4092. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, S.; Vellingiri, K. Effect of red yeast rice and coconut, rice bran or sunflower oil combination in rats on hypercholesterolemic diet. J. Clin. Diagn. Res. 2016, 10, BF05–BF07. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. The PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The prisma statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Boesch-Saadatmandi, C.; Niering, J.; Minihane, A.M.; Wiswedel, I.; Gardeman, A.; Wolffram, S.; Rimbach, G. Impact of apolipoprotein e genotype and dietary quercetin on paraoxonase 1 status in ApoE3 and ApoE4 transgenic mice. Atherosclerosis 2010, 211, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Dalgard, C.; Christiansen, L.; Jonung, T.; Mackness, M.I.; de Maat, M.P.; Horder, M. No influence of increased intake of orange and blackcurrant juices and dietary amounts of vitamin E on paraoxonase-1 activity in patients with peripheral arterial disease. Eur. J. Nutr. 2007, 46, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Choi, M.S.; Jung, U.J.; Kim, M.J.; Kim, D.J.; Park, H.M.; Park, Y.B.; Lee, M.K. Eucommia ulmoides oliver leaf extract increases endogenous antioxidant activity in type 2 diabetic mice. J. Med. Food 2006, 9, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Mazumder, S. An aqueous extract of Murraya koenigii leaves induces paraoxonase 1 activity in streptozotocin induced diabetic mice. Food Funct. 2013, 4, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Kiyici, A.; Okudan, N.; Gokbel, H.; Belviranli, M. The effect of grape seed extracts on serum paraoxonase activities in streptozotocin-induced diabetic rats. J. Med. Food 2010, 13, 725–728. [Google Scholar] [CrossRef] [PubMed]
- El-Beshbishy, H.A.; Singab, A.N.; Sinkkonen, J.; Pihlaja, K. Hypolipidemic and antioxidant effects of Morus alba L. (egyptian mulberry) root bark fractions supplementation in cholesterol-fed rats. Life Sci. 2006, 78, 2724–2733. [Google Scholar] [CrossRef] [PubMed]
- Noll, C.; Hamelet, J.; Matulewicz, E.; Paul, J.L.; Delabar, J.M.; Janel, N. Effects of red wine polyphenolic compounds on paraoxonase-1 and lectin-like oxidized low-density lipoprotein receptor-1 in hyperhomocysteinemic mice. J. Nutr. Biochem. 2009, 20, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Farrell, N.; Norris, G.; Lee, S.G.; Chun, O.K.; Blesso, C.N. Anthocyanin-rich black elderberry extract improves markers of HDL function and reduces aortic cholesterol in hyperlipidemic mice. Food Funct. 2015, 6, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Ku, C.S.; Pham, T.X.; Park, Y.; Martin, D.A.; Xie, L.; Taheri, R.; Lee, J.; Bolling, B.W. Aronia melanocarpa (chokeberry) polyphenol-rich extract improves antioxidant function and reduces total plasma cholesterol in apolipoprotein e knockout mice. Nutr. Res. 2013, 33, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Rizvi, S.I. Onion extract (Allium cepa L.), quercetin and catechin up-regulate paraoxonase 1 activity with concomitant protection against low-density lipoprotein oxidation in male Wistar rats subjected to oxidative stress. J. Sci. Food Agric. 2014, 94, 2752–2757. [Google Scholar] [CrossRef] [PubMed]
- Francik, R.; Krosniak, M.; Sanocka, I.; Barton, H.; Hebda, T.; Francik, S. Aronia melanocarpa treatment and antioxidant status in selected tissues in Wistar rats. Biomed. Res. Int. 2014, 2014, 457085. [Google Scholar] [CrossRef] [PubMed]
- Francik, R.; Kryczyk, J.; Krosniak, M.; Berkoz, M.; Sanocka, I.; Francik, S. The neuroprotective effect of cornus mas on brain tissue of Wistar rats. Sci. World J. 2014, 2014, 847368. [Google Scholar] [CrossRef] [PubMed]
- Nounou, H.A.; Deif, M.M.; Shalaby, M.A. Effect of flaxseed supplementation and exercise training on lipid profile, oxidative stress and inflammation in rats with myocardial ischemia. Lipids Health Dis. 2012, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Caliskan, A.; Karahan, O.; Yazici, S.; Demirtas, S.; Guclu, O.; Tezcan, O.; Yavuz, C. Protective effects of ginseng extracts and common anti-aggregant drugs on ischaemia-reperfusion injury. Cardiovasc. J. Afr. 2015, 26, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Mohammadshahi, M.; Haidari, F.; Saei, A.A.; Rashidi, B.; Mahboob, S.; Rashidi, M.R. Soy protein, genistein, and daidzein improve serum paraoxonase activity and lipid profiles in rheumatoid arthritis in rats. J. Med. Food 2013, 16, 147–154. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.O.; Silva, M.; Silva, M.E.; Oliveira Rde, P.; Pedrosa, M.L. Diet supplementation with acai (Euterpe oleracea Mart.) pulp improves biomarkers of oxidative stress and the serum lipid profile in rats. Nutrition 2010, 26, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Perez Mendez, O.; Garcia Hernandez, L. High-density lipoproteins (HDL) size and composition are modified in the rat by a diet supplemented with “Hass” avocado (Persea americana Miller). Arch. Cardiol. Mex. 2007, 77, 17–24. [Google Scholar] [PubMed]
- Janeesh, P.A.; Abraham, A. Amelioration of cholesterol induced atherosclerosis by normalizing gene expression, cholesterol profile and antioxidant enzymes by Vigna unguiculata. Plant Foods Hum. Nutr. 2013, 68, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Menini, T.; Heck, C.; Schulze, J.; de Mejia, E.; Gugliucci, A. Protective action of Ilex paraguariensis extract against free radical inactivation of paraoxonase-1 in high-density lipoprotein. Planta Med. 2007, 73, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Begcevic, I.; Simundic, A.M.; Nikolac, N.; Dobrijevic, S.; Rajkovic, M.G.; Tesija-Kuna, A. Can cranberry extract and vitamin C + Zn supplements affect the in vivo activity of paraoxonase 1, antioxidant potential, and lipid status? Clin. Lab. 2013, 59, 1053–1060. [Google Scholar] [PubMed]
- Shidfar, F.; Rajab, A.; Rahideh, T.; Khandouzi, N.; Hosseini, S.; Shidfar, S. The effect of ginger (Zingiber officinale) on glycemic markers in patients with type 2 diabetes. J. Complement. Integr. Med. 2015, 12, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Qian, Q.; Qian, S.; Fan, P.; Huo, D.; Wang, S. Effect of salvia miltiorrhiza hydrophilic extract on antioxidant enzymes in diabetic patients with chronic heart disease: A randomized controlled trial. Phytother. Res. 2012, 26, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, B.; Ekbul, A.; Topal, N.B.; Sarandol, E.; Sag, S.; Baser, K.H.; Cordan, J.; Gullulu, S.; Tuncel, E.; Baran, I.; et al. Effects of Origanum onites on endothelial function and serum biochemical markers in hyperlipidaemic patients. J. Int. Med. Res. 2008, 36, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.C.; Wu, L.C.; Lai, J.M.; Chen, C.T.; Hsueh, C.M.; Hsu, S.L. Ethanol extract of Graptopetalum paraguayense upregulates paraoxonase 1 gene expression via an AKT/NF-κB-dependent pathway. Am. J. Chin. Med. 2012, 40, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Bastos, D.H. Chlorogenic acid protects paraoxonase 1 activity in high density lipoprotein from inactivation caused by physiological concentrations of hypochlorite. Fitoterapia 2009, 80, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Deakova, Z.; Orszaghova, Z.; Andrezalova, L.; Slezak, P.; Lehotay, J.; Muchova, J.; Burki, C.; Durackova, Z. Influence of oak wood polyphenols on cysteine, homocysteine and glutathione total levels and PON1 activities in human adult volunteers—A pilot study. Gen. Physiol. Biophys. 2015, 34, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Volkova, N.; Borochov-Neori, H.; Judeinstein, S.; Aviram, M. Anti-atherogenic properties of date vs. Pomegranate polyphenols: The benefits of the combination. Food Funct. 2015, 6, 1496–1509. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Rosenblat, M.; Gaitini, D.; Nitecki, S.; Hoffman, A.; Dornfeld, L.; Volkova, N.; Presser, D.; Attias, J.; Liker, H.; et al. Pomegranate juice consumption for 3 years by patients with carotid artery stenosis reduces common carotid intima-media thickness, blood pressure and LDL oxidation. Clin. Nutr. 2004, 23, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, M.; Hayek, T.; Aviram, M. Anti-oxidative effects of pomegranate juice (PJ) consumption by diabetic patients on serum and on macrophages. Atherosclerosis 2006, 187, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Balbir-Gurman, A.; Fuhrman, B.; Braun-Moscovici, Y.; Markovits, D.; Aviram, M. Consumption of pomegranate decreases serum oxidative stress and reduces disease activity in patients with active rheumatoid arthritis: A pilot study. Isr. Med. Assoc. J. 2011, 13, 474–479. [Google Scholar] [PubMed]
- Fuhrman, B.; Volkova, N.; Aviram, M. Pomegranate juice polyphenols increase recombinant paraoxonase-1 binding to high-density lipoprotein: Studies in vitro and in diabetic patients. Nutrition 2010, 26, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Volkova, N.; Coleman, R.; Dreher, M.; Reddy, M.K.; Ferreira, D.; Rosenblat, M. Pomegranate phenolics from the peels, arils, and flowers are antiatherogenic: Studies in vivo in atherosclerotic apolipoprotein E-deficient (E0) mice and in vitro in cultured macrophages and lipoproteins. J. Agric. Food Chem. 2008, 56, 1148–1157. [Google Scholar] [CrossRef] [PubMed]
- Rock, W.; Rosenblat, M.; Miller-Lotan, R.; Levy, A.P.; Elias, M.; Aviram, M. Consumption of wonderful variety pomegranate juice and extract by diabetic patients increases paraoxonase 1 association with high-density lipoprotein and stimulates its catalytic activities. J. Agric. Food Chem. 2008, 56, 8704–8713. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.T.; Fitschen, P.J.; Kistler, B.M.; Jeong, J.H.; Chung, H.R.; Aviram, M.; Phillips, S.A.; Fernhall, B.; Wilund, K.R. Effects of pomegranate extract supplementation on cardiovascular risk factors and physical function in hemodialysis patients. J. Med. Food 2015, 18, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Park, E.M.; Bok, S.H.; Jung, U.J.; Kim, J.Y.; Park, Y.B.; Huh, T.L.; Kwon, O.S.; Choi, M.S. Two cinnamate derivatives produce similar alteration in mRNA expression and activity of antioxidant enzymes in rats. J. Biochem. Mol. Toxicol. 2003, 17, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schiborr, C.; Frank, J.; Rimbach, G. Curcumin induces paraoxonase 1 in cultured hepatocytes in vitro but not in mouse liver in vivo. Br. J. Nutr. 2011, 105, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.E.; Boesch-Saadatmandi, C.; Breckwoldt, D.; Schrader, C.; Schmelzer, C.; Doring, F.; Hashida, K.; Hori, O.; Matsugo, S.; Rimbach, G. Ascorbic acid partly antagonizes resveratrol mediated heme oxygenase-1 but not paraoxonase-1 induction in cultured hepatocytes—Role of the redox-regulated transcription factor Nrf2. BMC Complement. Altern. Med. 2011, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Do, G.M.; Kwon, E.Y.; Kim, H.J.; Jeon, S.M.; Ha, T.Y.; Park, T.; Choi, M.S. Long-term effects of resveratrol supplementation on suppression of atherogenic lesion formation and cholesterol synthesis in ApoE-deficient mice. Biochem. Biophys. Res. Commun. 2008, 374, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Noll, C.; Hamelet, J.; Ducros, V.; Belin, N.; Paul, J.L.; Delabar, J.M.; Janel, N. Resveratrol supplementation worsen the dysregulation of genes involved in hepatic lipid homeostasis observed in hyperhomocysteinemic mice. Food Chem. Toxicol. 2009, 47, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, D.; Winkler, J.; Parthiban, V.; Doss, M.X.; Hescheler, J.; Sachinidis, A. Dietary small molecules and large-scale gene expression studies: An experimental approach for understanding their beneficial effects on the development of malignant and non-malignant proliferative diseases. Curr. Med. Chem. 2006, 13, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Bok, S.H.; Jeong, T.S.; Moon, S.S.; Lee, S.E.; Park, Y.B.; Choi, M.S. Supplementation of naringenin and its synthetic derivative alters antioxidant enzyme activities of erythrocyte and liver in high cholesterol-fed rats. Bioorg. Med. Chem. 2002, 10, 2239–2244. [Google Scholar] [CrossRef]
- Kravchenko, L.V.; Avren’eva, L.I.; Aksenov, I.V.; Balakina, A.S.; Guseva, G.V.; Trusov, N.V. Effects of rutin on protective capacity in rats. Vopr. Pitan. 2015, 84, 22–30. [Google Scholar] [PubMed]
- Hsu, S.P.; Wu, M.S.; Yang, C.C.; Huang, K.C.; Liou, S.Y.; Hsu, S.M.; Chien, C.T. Chronic green tea extract supplementation reduces hemodialysis-enhanced production of hydrogen peroxide and hypochlorous acid, atherosclerotic factors, and proinflammatory cytokines. Am. J. Clin. Nutr. 2007, 86, 1539–1547. [Google Scholar] [PubMed]
- Schrader, C.; Ernst, I.M.; Sinnecker, H.; Soukup, S.T.; Kulling, S.E.; Rimbach, G. Genistein as a potential inducer of the anti-atherogenic enzyme paraoxonase-1: Studies in cultured hepatocytes in vitro and in rat liver in vivo. J. Cell. Mol. Med. 2012, 16, 2331–2341. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.Y.; Kim, S.S.; Chung, H.Y.; Yoon, S. Isoflavone supplements exert hormonal and antioxidant effects in postmenopausal Korean women with diabetic retinopathy. J. Med. Food 2005, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Clerici, C. Equol: History, chemistry, and formation. J. Nutr. 2010, 140, 1355S–1362S. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, X.; Zhang, Y.; Wang, Y.; Liu, Y.; Sun, R.; Xia, M. Anthocyanin supplementation improves HDL-associated paraoxonase 1 activity and enhances cholesterol efflux capacity in subjects with hypercholesterolemia. J. Clin. Endocrinol. Metab. 2014, 99, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Ferretti, G.; Bacchetti, T.; Masciangelo, S.; Pallotta, G. Lipid peroxidation in hemodialysis patients: Effect of vitamin C supplementation. Clin. Biochem. 2008, 41, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Ferre, N.; Camps, J.; Fernandez-Ballart, J.; Arija, V.; Murphy, M.M.; Marsillach, J.; Joven, J. Longitudinal changes in serum paraoxonase-1 activity throughout normal pregnancy. Clin. Chem. Lab. Med. 2006, 44, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Ardalic, D.; Stefanovic, A.; Kotur-Stevuljevic, J.; Vujovic, A.; Spasic, S.; Spasojevic-Kaliomanvska, V.; Jelic-Ivanovic, Z.; Mandic-Markovic, V.; Mikovic, Z.; Cerovic, N. The influence of maternal smoking habits before pregnancy and antioxidative supplementation during pregnancy on oxidative stress status in a non-complicated pregnancy. Adv. Clin. Exp. Med. 2014, 23, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Manolescu, B.N.; Berteanu, M.; Cinteza, D. Effect of the nutritional supplement ALAnerv® on the serum PON1 activity in post-acute stroke patients. Pharmacol. Rep. 2013, 65, 743–750. [Google Scholar] [CrossRef]
- Sarandol, E.; Tas, S.; Dirican, M.; Serdar, Z. Oxidative stress and serum paraoxonase activity in experimental hypothyroidism: Effect of vitamin E supplementation. Cell Biochem. Funct. 2005, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Motta, S.; Letellier, C.; Ropert, M.; Motta, C.; Thiebault, J.J. Protecting effect of vitamin E supplementation on submaximal exercise-induced oxidative stress in sedentary dogs as assessed by erythrocyte membrane fluidity and paraoxonase-1 activity. Vet. J. 2009, 181, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Tsakiris, S.; Karikas, G.A.; Parthimos, T.; Tsakiris, T.; Bakogiannis, C.; Schulpis, K.H. α-Tocopherol supplementation prevents the exercise-induced reduction of serum paraoxonase 1/arylesterase activities in healthy individuals. Eur. J. Clin. Nutr. 2009, 63, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, T.; Nouri, M.; Saei, A.A.; Rashidi, M.R. Aldehyde and xanthine oxidase activities in tissues of streptozotocin-induced diabetic rats: Effects of vitamin E and selenium supplementation. Biol. Trace Elem. Res. 2012, 147, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.Y.; Cho, Y.Y.; Do, G.M.; Kim, H.J.; Jeon, S.M.; Park, Y.B.; Lee, M.K.; Min, T.S.; Choi, M.S. Actions of ferulic acid and vitamin E on prevention of hypercholesterolemia and atherogenic lesion formation in apolipoprotein E-deficient mice. J. Med. Food 2009, 12, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, J.; Dopsaj, V.; Kotur-Stevuljevic, J.; Dopsaj, M.; Vujovic, A.; Stefanovic, A.; Nesic, G. Oxidative stress biomarker monitoring in elite women volleyball athletes during a 6-week training period. J. Strength Cond. Res. 2011, 25, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, J.; Dopsaj, V.; Kotur-Stevuljevic, J.; Dopsaj, M.; Vujovic, A.; Stefanovic, A.; Nesic, G. Proxidant antioxidant balance in supplemented elite female volleyball athletes during a six week training period. J. Sports Med. Phys. Fit. 2011, 51, 145–152. [Google Scholar]
- Haraguchi, F.K.; Pedrosa, M.L.; Paula, H.; Santos, R.C.; Silva, M.E. Evaluation of biological and biochemical quality of whey protein. J. Med. Food 2010, 13, 1505–1509. [Google Scholar] [CrossRef] [PubMed]
- Siktar, E.; Ekinci, D.; Siktar, E.; Beydemir, S.; Gulcin, I.; Gunay, M. Protective role of L-carnitine supplementation against exhaustive exercise induced oxidative stress in rats. Eur. J. Pharmacol. 2011, 668, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Dirican, M.; Tas, S.; Sarandol, E. High-dose taurine supplementation increases serum paraoxonase and arylesterase activities in experimental hypothyroidism. Clin. Exp. Pharmacol. Physiol. 2007, 34, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Durak, I.; Ozbek, H.; Devrim, E.; Karagenc, N.; Erguder, I.B. Effects of cholesterol supplementation on antioxidant enzyme activities in rat hepatic tissues: Possible implications of hepatic paraoxonase in atherogenesis. Nutr. Metab. Cardiovasc. Dis. 2004, 14, 211–214. [Google Scholar] [CrossRef]
- Acin, S.; Navarro, M.A.; Carnicer, R.; Arbones-Mainar, J.M.; Guzman, M.A.; Arnal, C.; Beltran, G.; Uceda, M.; Maeda, N.; Osada, J. Dietary cholesterol suppresses the ability of olive oil to delay the development of atherosclerotic lesions in apolipoprotein E knockout mice. Atherosclerosis 2005, 182, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Boukhari, N.; Taleb-Senouci, D.; Chabane, F.Z.; Besbes, M.; Lamri-Senhadji, M.Y. Fish by-products oil corrects dyslipidemia, improves reverse cholesterol transport and stimulates paraoxonase-1 activity in obese rat. Ann. Cardiol. Angeiol. 2013, 62, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kravchenko, L.V.; Aksenov, I.V.; Avren’eva, L.I.; Beketova, N.A.; Trusov, N.V.; Guseva, G.V. Effects of ω-3 polyunsaturated fatty acids on antioxidant capacity in rats. Vopr. Pitan. 2013, 82, 4–9. [Google Scholar] [PubMed]
- Popovic, T.; Borozan, S.; Arsic, A.; Martacic, J.D.; Vucic, V.; Trbovic, A.; Mandic, L.; Glibetic, M. Fish oil supplementation improved liver phospholipids fatty acid composition and parameters of oxidative stress in male Wistar rats. J. Anim. Physiol. Anim. Nutr. 2012, 96, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanihaghjo, A.; Kolahi, S.; Seifirad, S.; Rashtchizadeh, N.; Argani, H.; Hajialilo, M.; Khabazi, A.; Alizadeh, S.; Bahreini, E. Effect of fish oil supplements on serum paraoxonase activity in female patients with rheumatoid arthritis: A double-blind randomized controlled trial. Arch. Iran. Med. 2012, 15, 549–552. [Google Scholar] [PubMed]
- Burillo, E.; Mateo-Gallego, R.; Cenarro, A.; Fiddyment, S.; Bea, A.M.; Jorge, I.; Vazquez, J.; Civeira, F. Beneficial effects of ω-3 fatty acids in the proteome of high-density lipoprotein proteome. Lipids Health Dis. 2012, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Shidfar, F.; Amani, S.; Vafa, M.; Shekarriz, R.; Hosseini, S.; Shidfar, S.; Eshraghian, M.; Mousavi, S.N. Effects of iron supplementation with and without docosahexaenoic acid on the cardiovascular disease risk based on paraoxonase-1, hs-CRP, and ApoB/Apoa-I ratio in women with iron deficiency anemia. Biol. Trace Elem. Res. 2016, 169, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Whelan, J.; Gouffon, J.; Zhao, Y. Effects of dietary stearidonic acid on biomarkers of lipid metabolism. J. Nutr. 2012, 142, 630S–634S. [Google Scholar] [CrossRef] [PubMed]
- Bruge, F.; Bacchetti, T.; Principi, F.; Scarpa, E.S.; Littarru, G.P.; Tiano, L. Olive oil supplemented with coenzyme Q10: Effect on plasma and lipoprotein oxidative status. Biofactors 2012, 38, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Gabas-Rivera, C.; Barranquero, C.; Martinez-Beamonte, R.; Navarro, M.A.; Surra, J.C.; Osada, J. Dietary squalene increases high density lipoprotein-cholesterol and paraoxonase 1 and decreases oxidative stress in mice. PLoS ONE 2014, 9, e104224. [Google Scholar] [CrossRef] [PubMed]
- Baralic, I.; Djordjevic, B.; Dikic, N.; Kotur-Stevuljevic, J.; Spasic, S.; Jelic-Ivanovic, Z.; Radivojevic, N.; Andjelkovic, M.; Pejic, S. Effect of astaxanthin supplementation on paraoxonase 1 activities and oxidative stress status in young soccer players. Phytother. Res. 2013, 27, 1536–1542. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.D.; Bansal, M.P. Studies on HDL associated enzymes under experimental hypercholesterolemia: Possible modulation on selenium supplementation. Lipids Health Dis. 2009, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Hidiroglou, N.; Lok, E.; Taylor, M.; Kapal, K.; Ross, N.; Sarafin, K.; Lau, A.; de Souza, A.; Chan, H.M.; et al. Dietary selenium (Se) and vitamin E (VE) supplementation modulated methylmercury-mediated changes in markers of cardiovascular diseases in rats. Cardiovasc. Toxicol. 2012, 12, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Ferre, N.; Camps, J.; Cabre, M.; Paul, A.; Joven, J. Hepatic paraoxonase activity alterations and free radical production in rats with experimental cirrhosis. Metabolism 2001, 50, 997–1000. [Google Scholar] [PubMed]
- Helmy, M.M.; Mahmoud, S.S.; Fahmy, Z.H. Schistosoma mansoni: Effect of dietary zinc supplement on egg granuloma in Swiss mice treated with praziqantel. Exp. Parasitol. 2009, 122, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Ardabili, B.; Argani, H.; Ghorbanihaghjo, A.; Rashtchizadeh, N.; Naghavi-Behzad, M.; Ghorashi, S.; Nezami, N. Paraoxonase enzyme activity is enhanced by zinc supplementation in hemodialysis patients. Ren. Fail. 2012, 34, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Klevay, L.M. Ischemic heart disease as deficiency disease. Cell. Mol. Biol. 2004, 50, 877–884. [Google Scholar] [PubMed]
- Alp, H.; Varol, S.; Celik, M.M.; Altas, M.; Evliyaoglu, O.; Tokgoz, O.; Tanriverdi, M.H.; Uzar, E. Protective effects of β-glucan and gliclazide on brain tissue and sciatic nerve of diabetic rats induced by streptozosin. Exp. Diabetes Res. 2012, 2012, 230342. [Google Scholar] [CrossRef] [PubMed]
- Anwar, H.; Rahman, Z.U.; Javed, I.; Muhammad, F. Effect of protein, probiotic, and symbiotic supplementation on serum biological health markers of molted layers. Poult. Sci. 2012, 91, 2606–2613. [Google Scholar] [CrossRef] [PubMed]
- Summerbell, J.; Wynne, H.; Hankey, C.R.; Williams, F.M. The effect of age and frailty upon blood esterase activities and their response to dietary supplementation. Br. J. Clin. Pharmacol. 1993, 36, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Rainwater, D.L.; Mahaney, M.C.; VandeBerg, J.L.; Wang, X.L. Vitamin E dietary supplementation significantly affects multiple risk factors for cardiovascular disease in baboons. Am. J. Clin. Nutr. 2007, 86, 597–603. [Google Scholar] [PubMed]
- Jiang, H.; Stabler, S.P.; Allen, R.H.; Maclean, K.N. Altered expression of apoA-I, apoA-IV and PON-1 activity in CBS deficient homocystinuria in the presence and absence of treatment: Possible implications for cardiovascular outcomes. Mol. Genet. Metab. 2012, 107, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Shiner, M.; Fuhrman, B.; Aviram, M. Macrophage paraoxonase 2 (PON2) expression is up-regulated by pomegranate juice phenolic anti-oxidants via PPARγ and AP-1 pathway activation. Atherosclerosis 2007, 195, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Machado Mde, O.; Becker, A.M.; de Andrade, F.; Maraschin, M.; da Silva, E.L. Yerba mate (Ilex paraguariensis) enhances the gene modulation and activity of paraoxonase-2: In vitro and in vivo studies. Nutrition 2012, 28, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
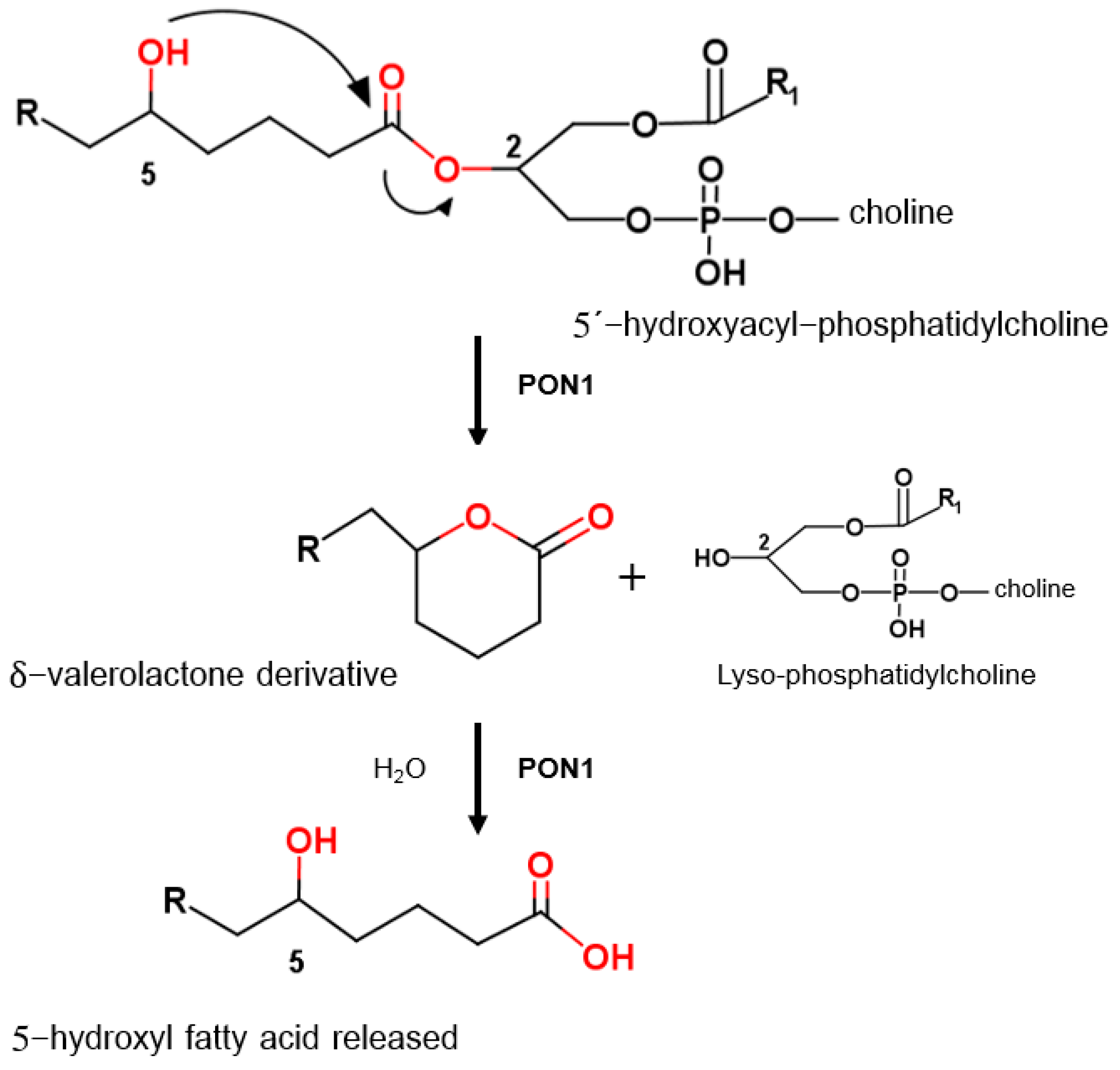

{kind=link}
{kind=link}
{kind=link}
| Extract | Experimental Model | Dose | Effect | References |
|---|---|---|---|---|
| Eucommia ulmoides Oliver leaf | Diabetic C57BL/KsJ-db/db mice | 400 mg/kg bw | ↑ 22% | [21] |
| Murraya koenigii | Streptozotocin-induced diabetic mice | 150 mg/kg | ↑ 105% | [22] |
| Grape seed extracts | Streptozotocin-induced diabetic rats | 100 mg/kg | ↑ 86% | [23] |
| Red wine polyphenol extract | Heterozygous Cbs-deficient mice | 100 mg/kg | ↑ 20% | [25] |
| Sambucus nigra | Apoe-deficient mice | 200 mg/kg | ↑ 20% | [26] |
| Aronia melanocarpa | Apoe-deficient mice | 6 mg/kg | ↑ 39% | [27] |
| Onion extract | Mercuric chloride-induced oxidative insult in male Wistar rats | 20 mg/kg | ↑ 30% | [28] |
| Aronia melanocarpa | Rats on a high-fructose and high-fat diet | Not reported | ↑ 65% | [29] |
| Cornelian cherry | Rats on a high-fructose and high-fat diets | Not reported | ↑ 45% | [30] |
| Genistein | Arthritic rats | 20 mg/kg | ↑ 230% | [33] |
| Euterpe oleracea Mart | Female Fischer rats on high-cholesterol, high-fat diets | 2 mg/kg | ↑ 60% | [34] |
| Avocado | Male Wistar rats | 28 g/kg | ↑ 33% | [35] |
| Ilex paraguariensis | Healthy volunteers | 0.5 L of extract | ↑ 10% | [37] |
| Cranberry extract with vitamin C and zinc | Healthy volunteers | 2 g/day (300 mg/day) | ↑ 67% | [38] |
| Zingiber officinale | Type 2 diabetic patients | 3 g | ↑ 28% | [39] |
| Salvia miltiorrhiza | Type 2 diabetic patients | Not reported | Increased PON1 activity | [40] |
| Origanum onites | Hyperlipidemic patients | Not reported | ↑ 14% | [41] |
| Compound | Experimental Model | Dose | Effect | References |
|---|---|---|---|---|
| 3-(4-Hydroxyphenyl) propionic acid | Rats fed a high-cholesterol diet | 1.35 mmol/kg | ↑ | [53] |
| Resveratrol | Apoe-deficient mice | 12 mg/kg | ↑75% | [56] |
| Flavonoids | ||||
| Naringenin | High-cholesterol-fed Sprague–Dawley rats | 4 mg/kg | ↑ 37% | [59] |
| Rutin | Wistar rats | 400 mg/kg | ↑ 17% | [60] |
| Quercetin | HgCl2 treated Wistar rats | 20 mg/kg | ↑ 20% | [28] |
| Genistein | Arthritic rats | 20 mg/kg | ↑ 230% | [33] |
| Tea catechins | Hemodialyzed patients | 6 mg/kg | ↑ 150% | [61] |
| Anthocyanins | Hypercholesterolemic subjects | 4 mg/kg | ↑ 22% | [65] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou-Bonafonte, J.M.; Gabás-Rivera, C.; Navarro, M.A.; Osada, J. The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases. Int. J. Mol. Sci. 2017, 18, 416. https://doi.org/10.3390/ijms18020416
Lou-Bonafonte JM, Gabás-Rivera C, Navarro MA, Osada J. The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases. International Journal of Molecular Sciences. 2017; 18(2):416. https://doi.org/10.3390/ijms18020416
Chicago/Turabian StyleLou-Bonafonte, José M., Clara Gabás-Rivera, María A. Navarro, and Jesús Osada. 2017. "The Search for Dietary Supplements to Elevate or Activate Circulating Paraoxonases" International Journal of Molecular Sciences 18, no. 2: 416. https://doi.org/10.3390/ijms18020416