Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development



Abstract
:
1. Introduction
2. Developmental Defects Caused by Mutations in Genes that Encode mitoRPs
2.1. Embryo-Lethal Mutations in mitoRPs
2.2. Effects of the Mutations in mitoRPs on Reproductive Tissues
2.3. Mutations in mitoRP Genes Affect Vegetative Development
2.3.1. Alterations in Leaf Morphology
2.3.2. Mutations in mitoRPs and the OGE Retrograde Signalling Pathway
3. Defective Mitoribosomal Function by Mutations in Mitochondrial Proteins Other than mitoRPs
4. Conclusions and Future Perspectives
Acknowledgments
Conflicts of Interest
References
- Selwood, S.P.; Chrzanowska-Lightowlers, Z.M.; Lightowlers, R.N. Does the mitochondrial transcription-termination complex play an essential role in controlling differential transcription of the mitochondrial DNA? Biochem. Soc. Trans. 2000, 28, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Bonawitz, N.D.; Clayton, D.A.; Shadel, G.S. Initiation and beyond: Multiple functions of the human mitochondrial transcription machinery. Mol. Cell 2006, 24, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.; Ramakrishnan, V. The structure of the human mitochondrial ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Brown, A.; Amunts, A.; Ramakrishnan, V. The structure of the yeast mitochondrial ribosome. Science 2017, 355, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Janska, H.; Kwasniak, M. Mitoribosomal regulation of OXPHOS biogenesis in plants. Front. Plant Sci. 2014, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Breiman, A.; Fieulaine, S.; Meinnel, T.; Giglione, C. The intriguing realm of protein biogenesis: Facing the green co-translational protein maturation networks. Biochim. Biophys. Acta 2016, 1864, 531–550. [Google Scholar] [CrossRef] [PubMed]
- Petersen, G.; Cuenca, A.; Moller, I.M.; Seberg, O. Massive gene loss in mistletoe (Viscum, Viscaceae) mitochondria. Sci. Rep. 2015, 5, 17588. [Google Scholar] [CrossRef] [PubMed]
- Bonen, L.; Calixte, S. Comparative analysis of bacterial-origin genes for plant mitochondrial ribosomal proteins. Mol. Biol. Evol. 2006, 23, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Sormani, R.; Masclaux-Daubresse, C.; Daniel-Vedele, F.; Chardon, F. Transcriptional regulation of ribosome components are determined by stress according to cellular compartments in Arabidopsis thaliana. PLoS ONE 2011, 6, e28070. [Google Scholar] [CrossRef] [PubMed]
- Pinel, C.; Douce, R.; Mache, R. A study of mitochondrial ribosomes from the higher plant Solanum tuberosum L. Mol. Biol. Rep. 1986, 11, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Maffey, L.; Degand, H.; Boutry, M. Partial purification of mitochondrial ribosomes from broad bean and identification of proteins encoded by the mitochondrial genome. Mol. Gen. Genet. 1997, 254, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, H.G. Components of bacterial ribosomes. Annu. Rev. Biochem. 1982, 51, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Szick-Miranda, K.; Chang, I.F.; Guyot, R.; Blanc, G.; Cooke, R.; Delseny, M.; Bailey-Serres, J. The organization of cytoplasmic ribosomal protein genes in the Arabidopsis genome. Plant Physiol. 2001, 127, 398–415. [Google Scholar] [CrossRef] [PubMed]
- Van Lijsebettens, M.; Vanderhaeghen, R.; De Block, M.; Bauw, G.; Villarroel, R.; Van Montagu, M. An S18 ribosomal protein gene copy at the Arabidopsis PFL locus affects plant development by its specific expression in meristems. EMBO J. 1994, 13, 3378–3388. [Google Scholar] [PubMed]
- Williams, M.E.; Sussex, I.M. Developmental regulation of ribosomal protein L16 genes in Arabidopsis thaliana. Plant J. 1995, 8, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Hulm, J.L.; McIntosh, K.B.; Bonham-Smith, P.C. Variation in transcript abundance among the four members of the Arabidopsis thaliana RIBOSOMAL PROTEIN S15a gene family. Plant Sci. 2005, 169, 267–278. [Google Scholar] [CrossRef]
- Makrides, S.C.; Goldthwaite, J. Biochemical changes during bean leaf growth, maturity and senescence. Content of DNA, polyribosomes, ribosomal RNA, protein and chlorophyll. J. Exp. Bot. 1981, 32, 725–735. [Google Scholar] [CrossRef]
- Whittle, C.A.; Krochko, J.E. Transcript profiling provides evidence of functional divergence and expression networks among ribosomal protein gene paralogs in Brassica napus. Plant Cell 2009, 21, 2203–2219. [Google Scholar] [CrossRef] [PubMed]
- Delage, L.; Giegé, P.; Sakamoto, M.; Maréchal-Drouard, L. Four paralogues of RPL12 are differentially associated to ribosome in plant mitochondria. Biochimie 2007, 89, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, M.; Day, R.C.; Talbot, M.J.; Ivanova, A.; Ashton, A.R.; Chaudhury, A.M.; Macknight, R.C.; Hrmova, M.; Koltunow, A.M. Developmentally regulated HEART STOPPER, a mitochondrially targeted L18 ribosomal protein gene, is required for cell division, differentiation, and seed development in Arabidopsis. J. Exp. Bot. 2015, 66, 5867–5880. [Google Scholar] [CrossRef] [PubMed]
- Schippers, J.; Mueller-Roeber, B. Ribosomal composition and control of leaf development. Plant Sci. 2010, 179, 307–315. [Google Scholar] [CrossRef]
- Mauro, V.P.; Edelman, G.M. The ribosome filter redux. Cell Cycle 2007, 6, 2246–2251. [Google Scholar] [CrossRef] [PubMed]
- Skinner, D.J.; Baker, S.C.; Meister, R.J.; Broadhvest, J.; Schneitz, K.; Gasser, C.S. The Arabidopsis HUELLENLOS gene, which is essential for normal ovule development, encodes a mitochondrial ribosomal protein. Plant Cell 2001, 13, 2719–2730. [Google Scholar] [CrossRef] [PubMed]
- Portereiko, M.F.; Sandaklie-Nikolova, L.; Lloyd, A.; Dever, C.A.; Otsuga, D.; Drews, G.N. NUCLEAR FUSION DEFECTIVE1 encodes the Arabidopsis RPL21M protein and is required for karyogamy during female gametophyte development and fertilization. Plant Physiol. 2006, 141, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.D.; Newton, K.J. The NCS3 mutation: Genetic evidence for the expression of ribosomal protein genes in Zea mays mitochondria. EMBO J. 1991, 10, 1045–1052. [Google Scholar] [PubMed]
- Sakamoto, W.; Kondo, H.; Murata, M.; Motoyoshi, F. Altered mitochondrial gene expression in a maternal distorted leaf mutant of Arabidopsis induced by Chloroplast mutator. Plant Cell 1996, 8, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Majewski, P.; Wołoszyńska, M.; Jańska, H. Developmentally early and late onset of Rps10 silencing in Arabidopsis thaliana: Genetic and environmental regulation. J. Exp. Bot. 2009, 60, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, P.; Masiero, S.; Eubel, H.; Braun, H.P.; Bhushan, S.; Glaser, E.; Salamini, F.; Leister, D. Nuclear photosynthetic gene expression is synergistically modulated by rates of protein synthesis in chloroplasts and mitochondria. Plant Cell 2006, 18, 970–991. [Google Scholar] [CrossRef] [PubMed]
- Shajani, Z.; Sykes, M.T.; Williamson, J.R. Assembly of bacterial ribosomes. Annu. Rev. Biochem. 2011, 80, 501–526. [Google Scholar] [CrossRef] [PubMed]
- Bryant, N.; Lloyd, J.; Sweeney, C.; Myouga, F.; Meinke, D. Identification of nuclear genes encoding chloroplast-localized proteins required for embryo development in Arabidopsis. Plant Physiol. 2011, 155, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Muralla, R.; Lloyd, J.; Meinke, D. Molecular foundations of reproductive lethality in Arabidopsis thaliana. PLoS ONE 2011, 6, e28398. [Google Scholar] [CrossRef] [PubMed]
- Neidhardt, F.C.; Curtiss, R. Escherichia coli and Salmonella: Cellular and Molecular Biology, 2nd ed.; American Society of Microbiology Press: Washington, DC, USA, 1996; ISBN 155-5-81-084-5. [Google Scholar]
- Wiegand, G.; Remington, S.J. Citrate synthase: Structure, control, and mechanism. Annu. Rev. Biophys. Biophys. Chem. 1986, 15, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Landschütze, V.; Willmitzer, L.; Müller-Röber, B. Inhibition of flower formation by antisense repression of mitochondrial citrate synthase in transgenic potato plants leads to a specific disintegration of the ovary tissues of flowers. EMBO J. 1995, 14, 660–666. [Google Scholar] [PubMed]
- Van Went, J.L.; Willemse, M.T.M. Fertilization. In Embryology of Angiosperms; Johri, B., Ed.; Springer: Berlin, Germany, 1984; pp. 273–318. [Google Scholar]
- Alexander, R.W.; Cooperman, B.S. Ribosomal proteins neighboring 23 S rRNA nucleotides 803–811 within the 50 S subunit. Biochemistry 1998, 37, 1714–1721. [Google Scholar] [CrossRef] [PubMed]
- Vladimirov, S.N.; Druzina, Z.; Wang, R.; Cooperman, B.S. Identification of 50S components neighboring 23 SrRNA nucleotides A2448 and U2604 within the peptidyl transferase center of Escherichia coli ribosomes. Biochemistry 2000, 39, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Christensen, C.A.; Gorsich, S.W.; Brown, R.H.; Jones, L.G.; Brown, J.; Shaw, J.M.; Drews, G.N. Mitochondrial GFA2 is required for synergid cell death in Arabidopsis. Plant Cell 2002, 14, 2215–2232. [Google Scholar] [CrossRef] [PubMed]
- Pagnussat, G.C.; Yu, H.J.; Ngo, Q.A.; Rajani, S.; Mayalagu, S.; Johnson, C.S.; Capron, A.; Xie, L.F.; Ye, D.; Sundaresan, V. Genetic and molecular identification of genes required for female gametophyte development and function in Arabidopsis. Development 2005, 132, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Pan, G.; Liu, H.; Wu, J.; Li, Y.; Zhao, Z.; Fu, T.; Zhou, Y. The chloroplast ribosomal protein L21 gene is essential for plastid development and embryogenesis in Arabidopsis. Planta 2012, 235, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Savage, L.J.; Imre, K.M.; Hall, D.A.; Last, R.L. Analysis of essential Arabidopsis nuclear genes encoding plastid-targeted proteins. PLoS ONE 2013, 8, e73291. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed]
- Shoji, S.; Dambacher, C.M.; Shajani, Z.; Williamson, J.R.; Schultz, P.G. Systematic chromosomal deletion of bacterial ribosomal protein genes. J. Mol. Biol. 2011, 413, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Breitenreuter, G.; Lotti, M.; Stöffler-Meilicke, M.; Stöffler, G. Comparative electron microscopic study on the location of ribosomal proteins S3 and S7 on the surface of the E. coli 30S subunit using monoclonal and conventional antibody. Mol. Gen. Genet. 1984, 197, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, F.J.; Nierhaus, K.H. Ribosomal proteins L15 and L16 are mere late assembly proteins of the large ribosomal subunit. Analysis of an Escherichia coli mutant lacking L15. J. Biol. Chem. 1990, 265, 16676–16682. [Google Scholar] [PubMed]
- Manavski, N.; Guyon, V.; Meurer, J.; Wienand, U.; Brettschneider, R. An essential pentatricopeptide repeat protein facilitates 5′ maturation and translation initiation of rps3 mRNA in maize mitochondria. Plant Cell 2012, 24, 3087–3105. [Google Scholar] [CrossRef] [PubMed]
- Burmann, B.M.; Schweimer, K.; Luo, X.; Wahl, M.C.; Stitt, B.L.; Gottesman, M.E.; Rösch, P. A NusE:NusG complex links transcription and translation. Science 2010, 328, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Kwasniak, M.; Majewski, P.; Skibior, R.; Adamowicz, A.; Czarna, M.; Sliwinska, E.; Janska, H. Silencing of the nuclear RPS10 gene encoding mitochondrial ribosomal protein alters translation in arabidopsis mitochondria. Plant Cell 2013, 25, 1855–1867. [Google Scholar] [CrossRef] [PubMed]
- Nomura, M. Regulation of ribosome biosynthesis in Escherichia coli and Saccharomyces cerevisiae: Diversity and common principles. J. Bacteriol. 1999, 181, 6857–6864. [Google Scholar] [PubMed]
- Fleischmann, T.T.; Scharff, L.B.; Alkatib, S.; Hasdorf, S.; Schöttler, M.A.; Bock, R. Nonessential plastid-encoded ribosomal proteins in tobacco: A developmental role for plastid translation and implications for reductive genome evolution. Plant Cell 2011, 23, 3137–3155. [Google Scholar] [CrossRef] [PubMed]
- Schuwirth, B.S.; Borovinskaya, M.A.; Hau, C.W.; Zhang, W.; Vila-Sanjurjo, A.; Holton, J.M.; Cate, J.H. Structures of the bacterial ribosome at 3.5 A resolution. Science 2005, 310, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Van Dyke, N.; Xu, W.; Murgola, E.J. Limitation of ribosomal protein L11 availability in vivo affects translation termination. J. Mol. Biol. 2002, 319, 329–339. [Google Scholar] [CrossRef]
- Pesaresi, P.; Varotto, C.; Meurer, J.; Jahns, P.; Salamini, F.; Leister, D. Knock-out of the plastid ribosomal protein L11 in Arabidopsis: Effects on mRNA translation and photosynthesis. Plant J. 2001, 27, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Linneweber, C.; Small, I. Pentatricopeptide repeat proteins: A socket set for organelle gene expression. Trends Plant Sci. 2008, 13, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Manna, S. An overview of pentatricopeptide repeat proteins and their applications. Biochimie 2015, 113, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Uyttewaal, M.; Mireau, H.; Rurek, M.; Hammani, K.; Arnal, N.; Quadrado, M.; Giegé, P. PPR336 is associated with polysomes in plant mitochondria. J. Mol. Biol. 2008, 375, 626–636. [Google Scholar] [CrossRef] [PubMed]
- Del Valle-Echevarria, A.R.; Kiełkowska, A.; Bartoszewski, G.; Havey, M.J. The Mosaic Mutants of Cucumber: A Method to Produce Knock-Downs of Mitochondrial Transcripts. G3 Genes Genomes Genet. 2015, 5, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Del Valle-Echevarria, A.R.; Sanseverino, W.; Garcia-Mas, J.; Havey, M.J. Pentatricopeptide repeat 336 as the candidate gene for paternal sorting of mitochondria (Psm) in cucumber. Theor. Appl. Genet. 2016, 129, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Selmer, M.; Dunham, C.M.; Murphy, F.V.; Weixlbaumer, A.; Petry, S.; Kelley, A.C.; Weir, J.R.; Ramakrishnan, V. Structure of the 70S ribosome complexed with mRNA and tRNA. Science 2006, 313, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, V.; Nierhaus, K.H. Assembly of the 30S subunit from Escherichia coli ribosomes occurs via two assembly domains which are initiated by S4 and S7. Biochemistry 1988, 27, 7051–7055. [Google Scholar] [CrossRef] [PubMed]
- Hammani, K.; Gobert, A.; Hleibieh, K.; Choulier, L.; Small, I.; Giegé, P. An Arabidopsis dual-localized pentatricopeptide repeat protein interacts with nuclear proteins involved in gene expression regulation. Plant Cell 2011, 23, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Xiu, Z.H.; Meeley, R.; Tan, B.C. Empty Pericarp5 encodes a pentatricopeptide repeat protein that is required for mitochondrial RNA editing and seed development in maize. Plant Cell 2013, 25, 868–883. [Google Scholar] [CrossRef] [PubMed]
- Brehme, N.; Bayer-Császár, E.; Glass, F.; Takenaka, M. The DYW Subgroup PPR Protein MEF35 Targets RNA Editing Sites in the Mitochondrial rpl16, nad4 and cob mRNAs in Arabidopsis thaliana. PLoS ONE 2015, 10, e0140680. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Gao, C.; Qiu, J.L. Progress and prospects in plant genome editing. Nat. Plants 2017, 3, 17107. [Google Scholar] [CrossRef] [PubMed]
- Jo, A.; Ham, S.; Lee, G.H.; Lee, Y.I.; Kim, S.; Lee, Y.S.; Shin, J.H.; Lee, Y. Efficient Mitochondrial Genome Editing by CRISPR/Cas9. Biomed. Res. Int. 2015, 2015, 305716. [Google Scholar] [CrossRef] [PubMed]
- Candela, H.; Pérez-Pérez, J.M.; Micol, J.L. Uncovering the post-embryonic functions of gametophytic- and embryonic-lethal genes. Trends Plant Sci. 2011, 16, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Chaiwongsar, S.; Strohm, A.K.; Su, S.H.; Krysan, P.J. Genetic analysis of the Arabidopsis protein kinases MAP3Kε1 and MAP3Kε2 indicates roles in cell expansion and embryo development. Front. Plant Sci. 2012, 3, 228. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Rivera, B.; Dawe, R.K. An Arabidopsis tissue-specific RNAi method for studying genes essential to mitosis. PLoS ONE 2012, 7, e51388. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Kobayashi, K.; Nakamura, Y.; Wada, H. Inducible knockdown of MONOGALACTOSYLDIACYLGLYCEROL SYNTHASE1 reveals roles of galactolipids in organelle differentiation in Arabidopsis cotyledons. Plant Physiol. 2014, 166, 1436–1449. [Google Scholar] [CrossRef] [PubMed]
- Dinman, J.D. Pathways to Specialized Ribosomes: The Brussels Lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gutierrez-Vargas, C.; Wei, J.; Grassucci, R.A.; Sun, M.; Espina, N.; Madison-Antenucci, S.; Tong, L.; Frank, J. Determination of the ribosome structure to a resolution of 2.5 Å by single-particle cryo-EM. Protein Sci. 2017, 26, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Sierra, R.G.; Gati, C.; Laksmono, H.; Dao, E.H.; Gul, S.; Fuller, F.; Kern, J.; Chatterjee, R.; Ibrahim, M.; Brewster, A.S.; et al. Concentric-flow electrokinetic injector enables serial crystallography of ribosome and photosystem II. Nat. Methods 2016, 13, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83. [Google Scholar] [CrossRef] [PubMed]
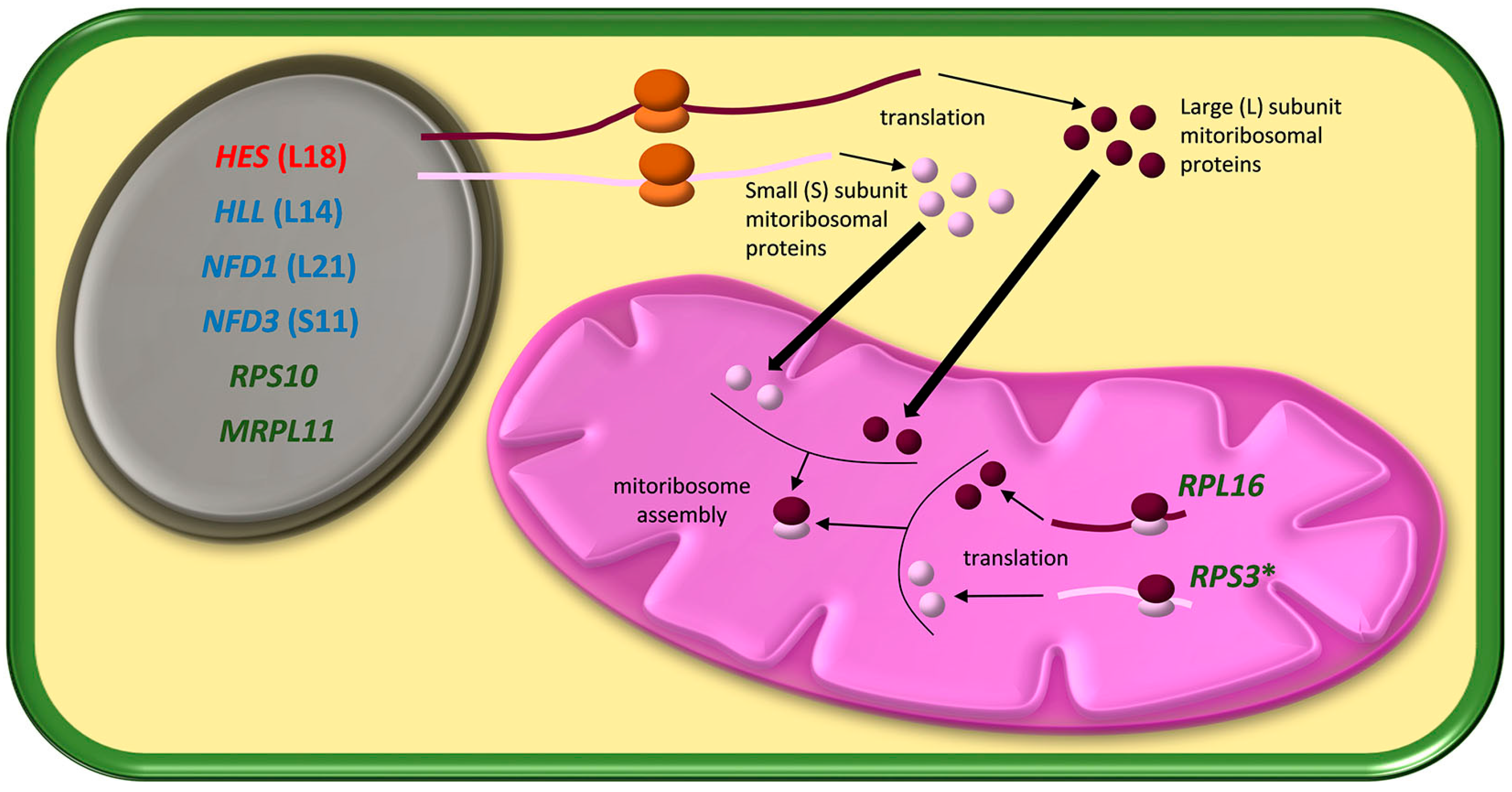

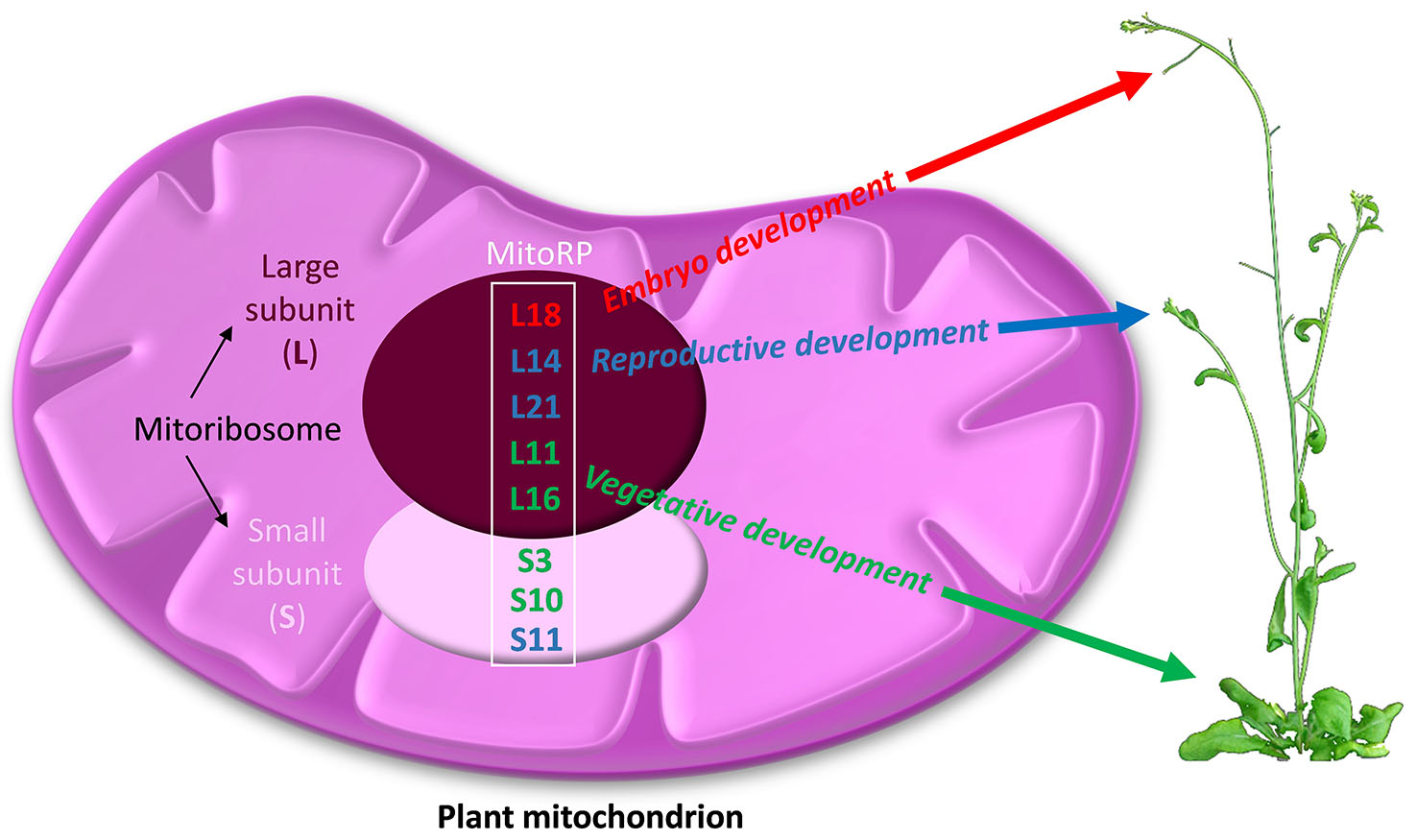{kind=link}
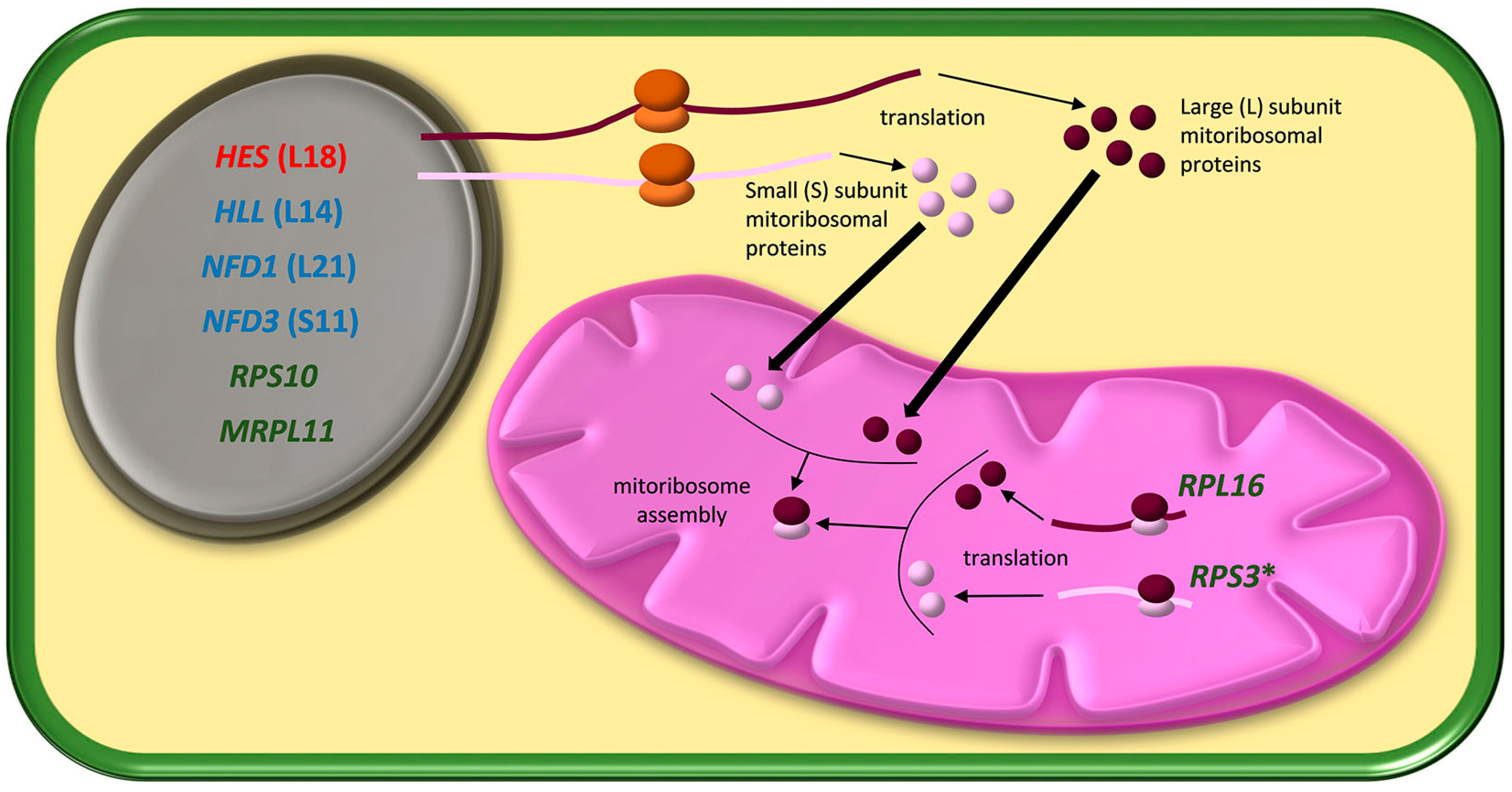{kind=link}
| Defects in | Gene | mitoRP a | Species | Mutant Phenotype |
|---|---|---|---|---|
| Embryo development | HEART STOPPER (HES) b AT1G08845 d | L18 | Arabidopsis thaliana | Reduced proliferation of endosperm cells and arrested embryo development in the late globular stage [20] |
| Reproductive development | HUELLENLOS (HLL) b AT1G17560 d | L14 | Arabidopsis thaliana | Early cellular degeneration of the eggs, characterised by arrested ovule development before or just after the formation of the integuments (hll-1) or after the integuments have begun to spread around the nucela (hll-2). hll-1 and hll-2 also show alterations in the gynoecium [23] |
| NUCLEAR FUSION DEFECTIVE1 (NFD1) b AT4G30925 d | L21 | Arabidopsis thaliana | Defective in kariogamy during fertilization and development of the female and male gametophytes [24] | |
| NFD3 b AT1G31817 d | S11 | Arabidopsis thaliana | Defective in kariogamy during fertilization and development of the female gametophyte [24] | |
| Vegetative development | rps3 c and rpl16 c | S3 and L16 | Zea mays | Sectors of poorly developed tissue on leaves and ears, which result from the segregation of somatic wild-type and mutant mitochondria [25] |
| rps3 c and rpl16 c AtMg00090 d and AtMg00080 d | S3 and L16 | Arabidopsis thaliana | Distorted leaf phenotype [26] | |
| Rps10 b AT3G22300 d | S10 | Arabidopsis thaliana | Plants homozygous for S10 silencing, show severe morphological alterations; they exhibit small, undulating, and yellowish leaves and died prior bolting [27] | |
| Mrpl11 b AT4G35490 d | L11 | Arabidopsis thaliana | Stunted plant size and a darker leaf coloring than the wild type [28] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robles, P.; Quesada, V. Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development. Int. J. Mol. Sci. 2017, 18, 2595. https://doi.org/10.3390/ijms18122595
Robles P, Quesada V. Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development. International Journal of Molecular Sciences. 2017; 18(12):2595. https://doi.org/10.3390/ijms18122595
Chicago/Turabian StyleRobles, Pedro, and Víctor Quesada. 2017. "Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development" International Journal of Molecular Sciences 18, no. 12: 2595. https://doi.org/10.3390/ijms18122595
APA StyleRobles, P., & Quesada, V. (2017). Emerging Roles of Mitochondrial Ribosomal Proteins in Plant Development. International Journal of Molecular Sciences, 18(12), 2595. https://doi.org/10.3390/ijms18122595