Control of Endogenous Auxin Levels in Plant Root Development


1
Department of Plant Production, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000 Gent, Belgium
2
Institute of Science and Technology Austria, Am Campus 1, 3400 Klosterneuburg, Austria
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(12), 2587; https://doi.org/10.3390/ijms18122587
Submission received: 26 October 2017
/
Revised: 26 November 2017
/
Accepted: 28 November 2017
/
Published: 1 December 2017
(This article belongs to the Special Issue Auxin)
Abstract
:In this review, we summarize the different biosynthesis-related pathways that contribute to the regulation of endogenous auxin in plants. We demonstrate that all known genes involved in auxin biosynthesis also have a role in root formation, from the initiation of a root meristem during embryogenesis to the generation of a functional root system with a primary root, secondary lateral root branches and adventitious roots. Furthermore, the versatile adaptation of root development in response to environmental challenges is mediated by both local and distant control of auxin biosynthesis. In conclusion, auxin homeostasis mediated by spatial and temporal regulation of auxin biosynthesis plays a central role in determining root architecture.
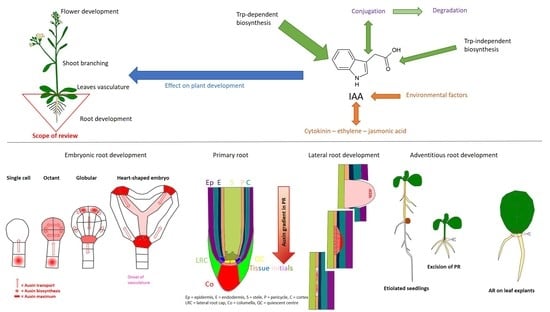
1. Introduction
Flowering plants have evolved a high level of developmental and morphological plasticity to accommodate adaptive growth in response to diverse environmental stimuli [1]. Unlike organ growth in animals, most plant organs are formed post-embryonically and emerge from inherent structures known as meristems [2]. Throughout the plant’s life span, meristems remain present and provide the basis for the plant’s developmental plasticity [3,4]. Initially at the embryonic state, two types of meristems occur: the shoot apical meristem (SAM), which gives rise to above ground tissues and organs and the root apical meristem (RAM), which establishes the root architecture. The root organ appears to have evolved secondary to the shoot allowing plants to invade land and facilitates the uptake of water for surviving the dry conditions [5]. The diversification of plants with the generation of varied plant forms, from tiny mosses to giant sequoia trees and from annual weeds to long-living perennials, was only possible after the emergence of a root system. Following the initiation of roots, features such as secondary growth, gravitropic responses and the development of lateral root branches are important additions that improved the function of the root system and required more complex cellular communication [5,6]. In particular, the roots of flowering plants have evolved highly specialized functions while maintaining developmental and morphological plasticity in order to cope with diverse environmental stimuli [1].
Plant roots explore the soil environment in search for nutrients and water required for growth, development and reproduction while containing a vascular system to provide organs at a distance with these necessary nutrients, water and hormones [7,8]. Generally, the root system consists of two principal root-types: the primary root (PR), which is formed embryonically [9] and secondary roots, which form post-embryonically. These secondary roots encompass both lateral roots (LR), which develop as branches of the primary root and adventitious roots (AR), which develop on non-root tissue such as the hypocotyl, stems and leaves [10,11]. The root architecture of monocots and dicots is highly distinct. Dicots have a tap-root system with a central primary root and lateral roots branching of, while monocots have a fibrous root system consisting of mainly crown or adventitious roots [12].
The plant growth regulator auxin is involved in basically all cellular processes including cell-division and -expansion with implications for all plant developmental processes [13]. Already at an early stage in embryogenesis, auxin determines cell division, tissue patterning and organ development. Likewise, at later developmental stages, auxin is involved in physiological growth responses such as gravitropism and shoot branching [6,14,15,16]. Since cell-division and -expansion requires to be spatiotemporally regulated, an appropriate auxin distribution across the tissue is highly important to coordinate growth and tissue development. Auxin homeostasis is assured not only by appropriate transport but also involves de novo biosynthesis, conjugation, storage, oxidation and catabolism, all of which need to take place in a coordinate fashion [17,18,19,20,21,22].
As an omnipotent regulator of root development, auxin controls the development and architecture of the root system [6,14]. In response to environmental cues, root growth is adapted through the modulation of endogenous auxin levels [23,24]. The establishment of auxin gradients in the root requires the interplay of local auxin biosynthesis [25,26,27,28], transport [29,30,31,32,33], perception and signaling [34,35,36]. In this review, we focus on the plant’s regulation of the endogenous auxin concentrations by biosynthesis, meta- and catabolism and conjugation. We provide information on how these processes are important for root development, encompassing the embryonic root, the PR and the secondary LR and AR. In the next paragraph, we briefly describe the molecular aspects of auxin perception (more can be found in e.g. References [37,38]). For reviews regarding the transport of auxin controlling root development see [16,39].
Auxin signals are perceived intracellularly by a family of AUXIN SIGNALING F-BOX (AFB) receptors of which (TIR1) TRANSPORT INHIBITOR RESPONSE1 is the founder. These F-box proteins are the substrate-binding subunit of SCF-type ubiquitin protein ligase complexes, named after their subunits Skp1, Cullin and an F-box [38]. In the presence of auxin, the SCFTIR1/AFB-complex is brought in close proximity to Aux/IAA (AUXIN/INDOLE-3-ACETIC ACID) proteins, which allows their ubiquitinylation and consequent targeting for proteasome degradation [40]. Normally, the Aux/IAA proteins act as transcriptional repressors by binding to Auxin Response Factor (ARF) transcription factors [41]. The removal of Aux/IAAs upon auxin perception hence results in the activation of ARFs, which on their turn activate the auxin responses. The model species Arabidopsis thaliana contains 6 AFBs [42,43], 29 AUX/IAAs [44] and 23 ARFs [45]. Different TIR1/AFB-Aux/IAA combinations may contribute to different transcriptional responses, depending on their presence in certain tissues or the physiological status of the plants. However, this field and the resulting variety of transcriptional responses is still rather ambiguous and predicting the function of a certain TIR1/AFB-Aux/IAA pair will require much more research [46]. Besides multiple combinations between receptors and Aux/IAAs, also the plenitude of ARFs might contribute to the specificity of the auxin response [38]. Some of the cellular responses to auxin take place at the cell surface. A candidate receptor of auxin at the plasma membrane is AUXIN BINDING PROTEIN 1 (ABP1) [34,36,47]. ABP1 is found at the plasma membrane and in the endoplasmatic reticulum and is shown to act via non-transcriptional processes [38,48]. Recently however, the contribution of ABP1 to auxin signaling and plant development has been questioned and we are awaiting its re-examination as a regulator of auxin perception [49,50].
The differential affinity of ABP1 and TIR1 for synthetic analogues of auxin suggests that different endogenous auxin analogues may contribute to a higher complexity of auxin signaling, possibly by separate ABP1 and TIR1/AFB modules [51]. So far four endogenous molecules—indole-3-acetic acid (IAA), indole-3-butyric acid (IBA), 4-chloroindole-3-acetic acid (4-Cl-IAA) and phenylacetic acid (PAA)—exhibit auxinic activity in plants [48,52,53,54]. IAA is most-studied and omnipresent in plants [17]. In planta, IAA is likely converted to IBA in a reversible reaction catalyzed by IBA synthetase but IBA could also serve as an IAA-precursor or -conjugate. Reversible β-oxidation of IBA to generate active IAA is a slow process and this slow release of active auxin could be the reason for the long-lasting function of IBA in rooting powders. Candidates for IBA to IAA conversion are four peroxisomal enzymes, three INDOLE- 3-BUTYRIC ACID RESPONSE (IBR) isozymes and one ENOYL–COA HYDRATASE2 (ECH2) [55,56,57]. IBA has been identified in a number of plant species such as maize (Zea mays), pea (Pisum sativum) and Arabidopsis thaliana but its inherent auxin-activity is still uncertain. Several studies provide evidence for a role of endogenous IBA in root development but a direct role in auxin signaling remains elusive [19,55,58,59]. The endogenous auxin-analogue PAA can bind to ABP1 but inhibits carrier-mediated auxin transport [48,60]. 4-Cl-IAA on the other hand has strong auxinic effects but also modulates processes that IAA does not affect and can be found exclusively in leguminous species [48,61]. Active auxins together with conjugated auxins constitute the auxin pool in a plant. In particular exogenous application of auxin leads to an increase of auxin-conjugating enzymes whereas conjugate hydrolases are activated to release stored inactive auxin in conjunction with initiation of growth such as during germination [62,63]. Besides the naturally occurring auxins, there are multiple synthetic auxin-derivatives, many of which were developed because of herbicide activity such as for example 2,4-dichlorophenoxyacetic acid (2,4-d), 1-naphthaleneacetic acid (NAA), dicamba and picloram [64].
2. Auxin Biosynthesis
Dissecting the auxin biosynthetic pathway has been proven difficult, largely because several auxin biosynthesis routes exist and the enzymes involved belong to gene families that show redundant functions [65,66,67]. Moreover, de novo auxin biosynthesis is not strictly confined to a specific tissue, although auxin is primarily synthesized in young, not fully expanded leaves [68,69]. Even though biosynthesis is the highest in the younger leaves, the largest auxin pools are measured in the root and in the expanding tissues where auxin is essential for the organ to grow. This underscores the importance of efficient transport between biosynthesis site and action site [68,70,71]. Other parts of the plant maintain the capacity to synthesize auxin as evidenced by the local production upon wounding or other local stimuli [25,69]. Below, we summarize recent findings on de novo auxin biosynthesis, which help to regulate the outcome of auxin homeostasis on root development.
The first enzymes identified to be engaged in the biosynthesis of auxin were from bacterial origin. The genes iaaM and iaaH were discovered in Pseudomonas and Agrobacterium and these enzymes respectively encode tryptophan-2-monooxygenase, which catalyzes the conversion of tryptophan to indole-3-acetamide (IAM) and a hydrolase that releases IAA [66,72]. Klee et al. [73] engineered petunia plants to express iaaM and this resulted in approximately a 10-fold increase in endogenous IAA-levels. Plants overproducing IAA showed typical auxin phenotypes such as excessive xylem and phloem development and bigger epidermal and palisade cells, which together with an extended intercellular space give rise to thicker leaves. Other bacterial IAA-biosynthetic pathways were soon thereafter discovered and for example, IPyA decarboxylase (IPDC) was identified to synthesize indole-3-pyruvic acid (IPyA) as an intermediate of auxin biosynthesis [74,75,76]. Orthologous of iaaM and iaaH have not been found in plants whereas the IPyA-mediated IAA-biosynthetic pathway was shown to occur in plants [25,67].
The predominant auxin biosynthesis route uses tryptophan as the main precursor. This was concluded in feeding experiments with labelled tryptophan (Trp), resulting in the production of labelled IAA, which indicated that plants use Trp as a precursor and contain the enzymes to convert Trp into IAA [77,78]. Tryptophan is synthesized from chorismate—the final product of the shikimate pathway—via intermediate indole-3-glycerol phosphate (IGP) in the chloroplasts [79]. In Arabidopsis thaliana, ASA1 and ASA2 encode the subunits of anthranilate synthase (AS), the enzyme necessary for the first step of tryptophan synthesis. Indole-3-glycerol phosphate synthase (IGS) is then responsible for the generation of IGP, a key intermediate in auxin biosynthesis because it can either form tryptophan or be used as a precursor for the tryptophan-independent auxin biosynthetic pathway [80]. Further conversion to tryptophan occurs through a heterotetrameric a2b2 complex consisting of Trp synthase a (TSA1) and Trp synthase b (TSB, in Arabidopsis there are 2 closely related versions of this gene: TSB1 and TSB2) [81]. A tryptophan-deficient Arabidopsis mutant, defective in anthranilate phosphoribosyl-transferase, accumulates anthranilic acid derivatives and these interfere with IAA-biosynthesis [82]. The observed morphological defects, including slow growth, small crinkled leaves and reduced apical dominance, also occur in mutants deficient in auxin signaling such as the auxin-resistant axr1, a gene encoding a subunit of the RUB1-activating enzymes regulating SCF-complex-mediated protein degradation [83]. The four most well-characterized auxin pathways use Trp as a precursor and are discussed below. Even though one of these pathways—the IPyA pathway—is considered the main route for auxin biosynthesis, the other routes act in their own way, in parallel or in a developmentally/environmentally-regulated manner.
2.1. The Indole-3-Pyruvic Acid (IPyA) Pathway
The existence of an IPyA auxin biosynthesis pathway was predicted based on the detection of certain intermediates and radiolabeling of tryptophan-derived metabolites [84]. The instability of IPyA at room temperature and its occurrence in two tautomer forms hampered the identification of enzymes involved [85]. The IPyA auxin biosynthesis enzymes were discovered only recently via genetic screens directed to identify regulators of organ development, ethylene response and shade avoidance. Defects in the corresponding genes lead to poor adaptation to low light conditions indicating the importance of this route of auxin biosynthesis in developmental plasticity. The IPyA pathway is a two-step reaction involving tryptophan aminotransferases (TAA1 and TARs) and Flavin-containing monooxygenases of the YUCCA family (YUC1-11) [86,87,88,89] (Blue Pathway in Figure 1). The Trp aminotransferase required for the first step were discovered by screening for ethylene insensitivity in roots, identifying wei8 (weak ethylene insensitive 8) [25] and by screening for lack of a shade avoidance response, identifying sav3 (shade avoidance 3) [67]. Wei8 and sav3 encode an aminotransferase that was renamed TAA1 (TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1). TAA1 catalyzes the transfer of the amino group from Trp to pyruvate or to α-ketoglutarate generating IPyA and the amino acids alanine or glutamine. Both the sav3 and wei8 mutants contain approximately 60% of the wild-type IAA-levels and show a strong impairment in dark induced hypocotyl elongation [67]. Another short hypocotyl mutant TRANSPORT INHIBITOR RESPONSE2 (TIR2) was shown to be allelic to TAA1 [26]. Overexpression of TAA1 causes modest auxin phenotypes indicating that this enzyme does not catalyze a rate-limiting biosynthesis step. The Arabidopsis TAA1 belongs to a small gene family that includes the homologues TAR1 to 4 (TRYPTOPHAN AMINOTRANSFERASE RELATED 1-4), which have overlapping functions [25]. Pea contains three TAA1 homologs: PsTAR1-3, which were shown to synthesize 4-Cl-IAA in addition to the canonical IAA [90].
Tryptophan feeding experiments identified a group of flavin monooxygenase-like enzymes—named YUCCA (YUC)—that catalyze the hydroxylation of the amino group of tryptamine, a rate-limiting step in IAOx tryptophan-dependent auxin biosynthesis pathway [91]. YUCs use NADPH and molecular oxygen to catalyze the oxidative decarboxylation of IPyA to IAA, using flavin as a cofactor [92,93]. Single yucca gene knockout mutants in Arabidopsis thaliana show wild-type growth phenotypes, likely because of redundancy among the 11 homologs [28]. Maize however possesses only 2 yucca copies and single mutants produce strong auxin-deficient phenotypes [94,95]. Overexpression of YUCs results in very limited effects on the IAA-levels, due to the strong homeostatic control of the active IAA-concentration but differences in IAA-metabolites can be clearly measured [87,93]. The YUC genes are mainly expressed in meristems, young primordia and reproductive organs, corresponding to sites of high auxin biosynthesis [69]. YUCs can also catalyze the decarboxylation of phenyl-pyruvate (PPA) to phenyl-acetic acid (PAA) [54]. The TAA- and YUC-family genes are regulated spatiotemporally and contribute to the uneven distribution of IAA in plants [25,26,66,67,96]. Homologs of the Arabidopsis YUC gene family have been found from several other plant species, suggesting a widespread occurrence of this branch throughout the plant kingdom.
Besides the linear pathway from IPyA to IAA, using the YUCs, indole-3-acetaldehyde (IAD) might be formed as an intermediate, like it happens in plant growth promoting rhizobacteria [97]. Whereas, the enzyme that could cause the conversion of IPyA to IAD has not been discovered in plants yet, the downstream aldehyde oxidases (AO) necessary to generate IAA out of IAD are present [98] (Blue Pathway in Figure 1).
2.2. The Indole-3-Acetamide (IAM) Pathway
IAM is the key intermediate in the bacterial auxin biosynthesis pathway of Agrobacterium tumefaciens. IAM is also found in many plant species including Arabidopsis, Citrus and Prunus, rice and maize [99,100,101]. By analogy with the bacterial synthesis route, the putative plant IAM-pathway would also encompass a two-step reaction with in the first step the conversion of Trp to IAM, which may be executed by a trypthophan-2-monooxygenase. Next, IAM is converted to IAA by the action of IAM HYDROLASE. Searching for orthologs of the bacterial iaaH in Arabidopsis thaliana has resulted in the identification of AtAMI1 (AMIDASE 1) as a candidate plant IAM hydrolase [102] (Yellow Pathway in Figure 1). Also, Nicotiana tabacum harbors an iaaH orthologue, NtAMI1, which is required for growth of BY2 cultures on medium, in which NAA is depleted but indole-3-acetamide is supplemented as alternative auxin-source [103]. Homologs of AMI have been identified in other plant species including the monocot rice and algae, suggesting that this auxin biosynthesis route is evolutionary conserved [104,105,106]. AMI1 homologs can convert phenyl-2-acetamide (PAM) into PAA, giving an explanation for the occurrence of auxinic molecules other than IAA in plants [107].
IAM may also be produced from indole-3-acetaldoxime (IAOx) because the IAM-levels are significantly decreased in mutants defective in the cytochrome P450 enzymes CYP79B2 and CYP79B3 (cyp79b2cyp79b3), involved in the IAOx pathway [98,108,109] (Yellow Pathway in Figure 1). However, in rice, maize or tobacco, species that lack the CYP79B2/3 homologs, IAM is still detected, which indicates that in these species the role of this pathway’s contribution to auxin biosynthesis is not entirely clear yet.
2.3. The Tryptamine (TAM) Pathway
Early reports of auxin activity in Avena curvature tests with tryptamine (TAM) suggest it acts as a precursor of auxin biosynthesis [105]. TAM is the first secondary metabolite derived from tryptophan in the synthesis of terpenoid indole alkaloids. In animals TAM is synthesized by tryptophan decarboxylase (TDC) in the cytosol [97] but a corresponding plant TDC-gene has hitherto not been found. TAM is converted to N-hydroxytryptamine in a rate-limiting step catalyzed by members of the YUCCA family [91] (Green Pathway in Figure 1). However, with the now completely described IPyA pathway, this double role of YUCs is questionable because N-hydroxytryptamine has not been detected in plants so far [89,93,110].
2.4. The Indole-3-Acetaldoxime (IAOx) Pathway
The IAOx route is unique to the Brassicaceae family [101]. Two homologous cytochrome P450 enzymes, CYP79B2 and CYP79B3 catalyze the synthesis of IAOx from Trp. This step takes place in the chloroplasts [108,109] (Pink Pathway in Figure 1). In the cyp79b2cyp79b3 double mutant, the IAOx levels were far below the detection limit, confirming the role of these enzymes [101]. In addition to its role as auxin precursor, IAOx is also shunted to several secondary metabolites such as indole glucosinolates (IG) and camalexin [111] (Pink Pathway in Figure 1). Mutations in glucosinolates biosynthesis genes SUR1 (superroot1), SUR2 or UGT74B1 (UDP-glucosyltransferase UGT74B1) resulted in elevated auxin accumulation [112,113,114]. The blocked IG biosynthesis may cause an increased flux of IAOx to IAA [28]. Because the levels of indole-3-acetonitrile (IAN) were also decreased in the cyp79b2cyp79b3 mutant, IAN is assumed to be a downstream intermediate of the IAOx pathway (Pink Pathway in Figure 1). Indeed, IAN can be formed by dehydration of IAOx and this reaction is catalyzed by CYP71A13 [115]. This IAN can be an intermediate for IAA but it is not found in all plant species [94]. Additionally, IAN is produced by hydrolysis of IG by myrosinases [116]. Auxin biosynthesis from IAN might involve nitrilases (NIT 1-4) but that process is not well understood yet [117,118] (Pink Pathway in Figure 1). The reduction of free IAA in the cyp79b2cyp79b3 is more significant at high temperature. This could indicate that the IAOx-pathway in Arabidopsis is involved in temperature-induced or -regulated IAA-formation [109].
2.5. Tryptophan-Independent Auxin Biosynthesis Pathways
Because tryptophan synthase-deficient mutants of Arabidopsis thaliana showed higher levels of IAA-conjugates, it was suggested that Arabidopsis harbors a tryptophan-independent auxin biosynthesis route [78]. Indeed, the intermediates IGP and indole may condense to form IAA, without Trp as an intermediate [82,119] (Gray Pathway in Figure 1). Decorticating the Trp-independent auxin biosynthesis route awaits further research. For reviews on this pathway see [17,80,85].
3. Auxin Biosynthesis Regulates Root Embryogenesis
The initiation of the apical-basal body plan of a plant embryo occurs during the initial stages of embryogenesis and depends on auxin gradients, transport and local biosynthesis [120,121,122,123]. Manipulation of auxin homeostasis by conditional expression of iaaM (indoleacetic acid-tryptophan mono-oxygenase) or the expression of a Pseudomonas syringae indoleacetic acid-lysine synthetase (iaaL) did not interfere with embryo-patterning [124]. Additionally, the sur mutants produced normal embryos but displayed auxin-overproduction phenotypes after germination, which indicates that the IAOx-biosynthetic route is not active during embryogenesis [125]. These results suggest that during embryogenesis there is a compensatory mechanism for biosynthesis-mediated changes in auxin homeostasis. On the other hand, inhibition of transport by 1-N-Naphthylphthalamic acid (NPA), led to fused cotyledons [126], which indicates that proper auxin distribution is critical during embryogenesis. Mutant studies attributed a major role to the PIN1 and PIN4 auxin transporters to establish and maintain the auxin gradient in the embryo [124]. But because of the feedback regulatory role of auxin itself on PIN-trafficking and -localization also auxin biosynthesis must be tightly regulated during embryogenesis [127,128,129].
Initially, auxin is supplied by the maternal tissue but the mathematical models do not exclude that local auxin production takes place in the suspensor [129,130]. This theory is substantiated by specific expression of YUC3, YUC4 and YUC9 in the suspensor and the localization of PIN7 to direct the synthesized IAA towards the developing pro-embryo [122,131]. Afterwards, auxin production in the pro-embryo starts and TAA1 expression is observed in the top apical cells of the 16-cell stage embryos. At a later stage, YUC1 and YUC4 are expressed in this region, while YUC8 is expressed closer to the root pole [96,131,132]. While at the apical part of the developing embryo cotyledons are initiated, the hypophysis, the uppermost suspensor cell, develops into the root meristem with the stem cells and quiescent center (QC) and onset of radial tissue patterning necessary for post-embryonic root growth [3,14,133]. Mutants missing YUC1, YUC4, YUC8, YUC10 or YUC11 failed to develop a hypocotyl and a root meristem, which confirmed the role of auxin generated by the YUCCA flavin monooxygenases during embryogenesis [96]. The expression of the auxin biosynthesis genes (TAA1/TAR and/or different YUC) in different regions of the developing embryo is furthermore essential for vasculature development and the initiation of cotyledons [25,67]. A paralog of Tryptophan synthase α (TSA), indole synthase (INS) is thought to act in a pathway parallel to TAA1. Cellular divisions in the ins-1 mutant embryo are disturbed and this indicates a role for INS during embryogenesis. Moreover, INS is expressed even earlier than TSA in the embryo and suspensor. Lastly, Trp-independent auxin biosynthesis is considered an important source of the plant growth regulator during embryo development [134].
4. Local Auxin Accumulation Regulates Root Development and Branching
A typical root system of dicotyledonous plants is composed of a primary root (PR) and lateral roots (LRs), which are secondary root branches, formed post- embryonically from PR to enable the plant to exploit the soil environment [135,136]. Common to all roots is the presence of a small group of cells at the center of the tip of the root organ. These cells are the stem cell niche, which coordinates the establishment of various tissues present in the root. Without these stem cells, root initials and the QC, the root can no longer grown and add cells to the developing tissue files. Critical to the activity of the QC and the root initials is the presence of an auxin maximum centered at the stem cell niche [62,137]. The formation of the auxin maximum at the root tip has been attributed to directional transport of shoot-derived auxin moving down into the root tip where complex patterns of auxin efflux carriers result in auxin accumulation at the stem cell niche [6,138,139,140]. Maintenance of the auxin maximum is however not solely the result of transport but also involves local auxin biosynthesis [141], which is possible in all parts of a root system [62,69]. Genes involved in Trp production (ASA1) and auxin biosynthesis genes encoding proteins of the IPyA pathway (TAA1, TAR2 and YUCCA) are expressed in the root tip [25,67,142]. The expression of YUC genes at the root tip depends on symplastic communication between the cells of the stem cell niche, suggesting that stem cell maintenance is intrinsically linked with the maintenance of a local auxin maximum [141]. Auxin derived from the shoot does not fully rescue the root growth and gravitropic responses characteristic for auxin-deficiency at the root tip, emphasizing the importance of local synthesis for root development [142]. In addition, locally synthesized auxin has been shown to rescue a local phenotype, yet does not show effects at a distance [66,96,143]. Also, root growth and patterning is strongly affected by deficiencies in local IPyA-mediated auxin biosynthesis. In rice, disruption of the FISH BONE gene, an orthologue of TAA1, affects many auxin-mediated processes including root development [144]. Antisense expression of YUCCA1 (OsYUC1) in rice results in a defective root, which phenocopies the root of auxin-insensitive mutants [145]. Similarly, loss of function mutants of the rice OsTDD1 gene, encoding anthranilate synthase β-subunit, have severe root defects but their phenotype can be rescued by overexpression of OsYUC1 [146]. In woodland strawberry (Fragaria vesca L.), silencing of YUCCA6 affected various post-embryonic organ developmental steps including root formation [147]. Many steps of Trp-synthesis involve transamination reactions, which require vitamin B6 as a cofactor. The pdx1 mutant, a vitamin B6 biosynthesis mutant with a short root, is defective in the generation of IAA from tryptophan. The root defects in this mutant are another indication that PR growth depends on Trp-dependent local auxin biosynthesis [148]. Other auxin biosynthetic pathways, such as the IAOx pathway are involved in root growth as well. For example, AtTGG4 and AtTGG5 act during the conversion of indole glucosinolates (IG) to the auxin precursor, indole-3-acetonitrile (IAN) and show defects in the establishment of an auxin gradient in the root tips resulting in a disturbed root growth [116]. Downstream nitrilase enzymes, such as NIT1, are involved in auxin biosynthesis in the root [149]. Furthermore, simultaneous inactivation of cyp79b2 and cyp79b3 results in a significant reduction of the auxin levels at the root tips [69,109]. However, due to redundancy in the different auxin biosynthetic pathways, most of the single biosynthetic mutations did not show dramatic root defects.
LRs develop from pericycle cells, adjacent to the xylem poles [150]. Within these so-called xylem pole pericycle founder cells, cell cycle reactivation and subsequent divisions result in the formation of an LR primordium. Priming of the founder cells occurs in the basal root meristem where it requires oscillating auxin fluxes [151,152,153,154]. The generation of IAA out of IBA seems to play a critical role in these oscillation patterns [58,59]. Programmed cell death at the lateral root cap is a second cause for the oscillatory auxin responses because it releases pulses of auxin into the surrounding root tissues [155]. Upon priming, the pericycle cells proceed through two cycles of asymmetric divisions, which gives rise to the formation of a single layer lateral root primordium (LRP) [156]. Further periclinal divisions yield a dome-shaped multilayer LRP, ready to emerge from the main PR [157,158,159]. Without the endogenous priming in the root meristem, a random stimulation of auxin biosynthesis in a pericycle cell results in an increase of auxin, sufficient to start the formation of an LR [160]. The Arabidopsis superroot1 (sur1) and sur2 have excessive amounts of LRs, indicating that local accumulation of auxin is sufficient to induce LR and AR formation [98,112,161,162]. These findings indicate that auxin accumulation is a trigger to modify the developmental program of the pericycle cell [163]. In young seedlings, early after germination, LR emergence depends on shoot-derived auxin but the capacity to synthesize auxin increases with the age of seedlings, which in turn reduces the dependence on shoot-derived auxin for LR development and local auxin biosynthesis takes over to control LR emergence [70]. The local synthesis of auxin appears to depend on auxin transport because disruption of the GNOM-dependent auxin transport resulted in reduced expression of YUCCA genes (YUC2, YUC3, YUC5 and YUC6) and TAA1 and TAR2. In gnom this reduced auxin accumulation at the sites of LRP formation resulted in fewer LR. A complex regulatory network combining auxin transport and local biosynthesis hence plays an important role in the initiation and further development of LR [164].
5. Importance of Auxin Biosynthesis for Adventitious Root Development
Adventitious root (AR) emerge from aerial parts of the plant in response to wounding, flooding and other stresses, or as part of a root developmental program [165,166,167]. There are two types of ARs: roots originating from latent and ready-made initials and de novo initiated AR [168]. Generally, ARs are initiated from tissues close to the vascular tissues including the pericycle in the hypocotyl, phloem or xylem parenchyma cells, interfascicular cambium cells, procambium and vascular parenchyma cells [10,11,168,169].
Like many other developmental processes, AR formation is regulated by external cues such as light, temperature and nutrient availability, which generate endogenous hormone signaling events [168,170]. However, wounding is probably the most powerful signal that induces de novo rooting process, especially when the shoot organ is severed from the root. A primary response to wounding is the accumulation of auxin by means of directional auxin transport and local auxin biosynthesis. In petunia cuttings, IAA peaked at the base of the stem early post-excision accompanied by a fast induction of YUCCA-family members and two IAA-amino acid hydrolases [171]. Their combined activity leads to a higher IAA-concentration typical for a wound-response [172]. A similar study in Arabidopsis thaliana showed that following root pruning the endogenous auxin levels rise due to polar auxin transport and increased expression of YUCCA9 [173,174]. Also for AR induction on detached leaves, YUCCA-mediated auxin synthesis was associated with root initiation [174]. For wound responses, YUC1 and YUC4 play the most important roles but it is not known yet how wound signals trigger these biosynthetic genes [174,175]. TAA1 family members are not induced by wounding, yet mutants lacking TAA1 and its closest homologue TAR2 are required for formation of roots on detached leaves [176]. YUC6-mediated auxin biosynthesis is needed at the tip of the AR to stimulate post-emergence growth, similar to local auxin production in the PR and LR tips [62,177,178]. This local auxin biosynthesis compensates for the reduction of auxin flow to the tip in the presence of cytokinin [177].
In general, easier-to-root varieties contain higher levels of free auxin whereas rooting-recalcitrant varieties have less auxin [179]. The positive correlation between auxin-levels and AR induction is further substantiated by spontaneous AR formation in auxin-overproduction mutants. Transgenic rice overexpressing YUCCA develop more crown roots [145]. The superroot mutants sur1 and sur2 are blocked in glucosinolated indole synthesis and thereby re-channel IAOx for IAA-synthesis, resulting in excessive AR formation [108,112,113,161,162]. The spontaneous rooting in sur1 and sur2 is lost in mutants with reduced Trp synthesis (e.g. mutants in ASA1, ASB1 and TSB1). However, other suppressors of sur2 are not controlling endogenous IAA-levels but are affected in auxin signaling [168]. A high auxin concentration in the plant is therefore as such not sufficient to induce AR.
6. Regulation of Auxin Homeostasis during Root Development by Other Hormones
The balance between auxin and cytokinin is of critical importance for determining the developmental fate of organs, with auxin promoting the root and cytokinin stimulating shoot development. At intermediate auxin/cytokinin ratios, callus, predominantly existing of actively dividing cells, is induced reflecting the stimulatory activity of both auxin and cytokinin [177,180]. The strong link between auxin and cytokinin (co)-activity is mediated by regulatory circuits that control each other’s production and transport. Constitutive downregulation of cytokinin was reflected in a significant downregulation of auxin biosynthesis [178]. Furthermore, cytokinin strongly inhibits PIN-type auxin efflux carriers [181,182,183]. On the other hand, cytokinin-induced repression of the auxin transport machinery resulted in secondary stimulation of the auxin biosynthetic gene YUCCA6 [177,184]. The connectivity between auxin and cytokinin is not necessarily conserved across plant species but in for example poplar and Arabidopsis, auxin biosynthesis was induced by an increase in cytokinin [185,186]. Microarrays and q-RT-PCRs showed altered expression of metabolic genes from different auxin biosynthetic pathways upon cytokinin treatment, confirming that cytokinin interferes with auxin biosynthesis [187]. One of these genes is the Cytokinin-induced Root Curling 6 (CKRC6), which is allelic to ASA1. The ckrc6 mutant is insensitive to both cytokinin and ethylene while containing less IAA [188]. This results in a reduced gravitropic root growth phenotype, which can be compensated by exogenous application of auxin. The downstream Trp synthesis gene ASB1 is also regulated by cytokinin by means of the response regulator ARABIDOPSIS RESPONSE REGULATOR (ARR1). Cytokinin-treatment of the root system leads to an increase in auxin at the transition zone of the root meristem due to locally increased ASB1 expression [189].
The cross-talk between ethylene and auxin inhibits root elongation and regulates LR initiation and emergence [190,191]. Ethylene regulates the expression of two WEAK ETHYLENE INSENSITIVE (WEI2/ASA1 and WEI7/ASB1) genes, which encode subunits of anthranilate synthase, a rate-limiting enzyme in Trp biosynthesis [25,192]. These genes were identified in a screen for ethylene insensitivity. WEI2 is allelic to ASA1, involved in Trp biosynthesis and WEI8 to TAA1, an enzyme in the IPyA auxin biosynthetic pathway. These mutants underscore the link between ethylene signaling and auxin biosynthesis [25]. In ethylene-treated seedlings, an overall increase of the auxin response at the root tip was observed and this is also reflected in direct auxin measurements [191,193]. A member of ethylene-responsive AP2 transcription factors; ETHYLENE RESPONSE FACTOR1 (ERF1) was recently shown to bind the promoter of ASA1 in order to regulate auxin biosynthesis and ethylene-induced root growth inhibition [194]. Another interaction between ethylene and auxin biosynthesis was discovered in a chemical genetic strategy, using L-kynurenine, a chemical that represses the nuclear accumulation of the ethylene insensitive 3 (EIN3) transcription factor and TAA1/TAR were identified as a target for L-kyn [195]. Lastly, rice transgenic plants overexpressing OsEIL1, the closest homolog of EIN3, exhibited a short, coiled primary root and they had increased YUC8/REIN7 expression so auxin biosynthesis through the IPyA pathway is activated [196].
Root growth is also affected by the interplay between auxin and jasmonic acid (JA). Exogenous application of methyl jasmonate enhances LR formation but inhibits PR elongation [197,198]. Jasmonates reduce LR formation in the Arabidopsis jasmonate-induced defective lateral root1 (jdl1/asa1-1) mutant. Inactivation of JDL1/ASA1 suppressed LR formation and reduced auxin accumulation in the basal meristem, which indicates that jasmonates not only affect auxin biosynthesis but also affect auxin transport [199]. The molecular mechanisms modulating the crosstalk between jasmonate and auxin biosynthesis during LR formation act through the Ethylene responsive transcription factor 109 (ERF109), which binds specifically to the promoter elements of YUC2 and ASA1. The JA-signaling pathway is also linked to auxin homeostasis through the modulation of YUCCA8 and YUCCA9 expression [200]. In conclusion, the auxin gradients needed for root development, secondary root initiation and emergence depend on synchronized interactions between auxin biosynthesis and interaction with other phytohormones.
7. Auxin Biosynthesis as an Integrator of Environmental Factors and Root Development
The primary function of the root system is to search for water and minerals and to provide physical support for the shoot to grow towards a light source. Hence light, water, minerals and gravitropic force influence root development and architecture. Light has been reported to regulate the endogenous level of auxins [201,202]. During AR formation, light influences the expression of the auxin responsive GH3 and ARF genes [203,204,205,206]. In Arabidopsis thaliana, the photomorphogenesis mutant red1 defective in SUR2/ATR4 (ALTERED TRYPTOPHAN REGULATION 4) failed to produce AR on the hypocotyl in red light [207]. Light is also shown to induce YUCCAs in the maize root [208]. De novo root organogenesis in Arabidopsis leaf explants involves different YUCCA genes depending on whether explants are incubated in the light or dark. The relevance of light as a factor regulating auxin homeostasis also follows from the identification of shade avoidance mutants some of which were affected in auxin biosynthesis genes. Tao et al. [67] identified TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS (TAA1) in a shade avoidance mutant screen. TAA1 indeed contributes to local auxin biosynthesis during the elongation of the hypocotyl [209]. In low R:FR conditions, PHYTOCHROME INTERACTING FACTOR 7 (PIF7) is activated via dephosphorylation and binds to the YUCCA genes to regulate their expression [210]. This suggests a direct link between the light-mediated PHYTOCHROME B (PHYB) pathway and auxin accumulation.
In Arabidopsis seedlings, exogenous application of sugar enhanced the PR growth and increased LR density by activating auxin biosynthesis [211]. The enhanced root growth correlated with transcriptional activation of the auxin biosynthesis genes YUC2, YUC8, YUC9, CYP79B2, CYP79B3, transporters PIN1, PIN2 and signaling AUX/IAA genes [211,212]. Similarly, sucrose grown seedlings develop elongated roots and auxin accumulates in the shoots and roots [213].
Root branching is enhanced in nutrient rich-soils [214,215,216,217]. Nitrate is a growth-limiting nutrient and the major source of nitrogen (N) for plants [218]. High and low concentrations of N significantly inhibit respectively enhance LR development with no or limited effect on growth of the PR [219,220]. N-depletion suppresses auxin-levels in the shoot while it stimulates auxin accumulation in the root, in line with a reduction in TAA1 and TAR1 expression in the shoot and upregulation in the root [221,222,223]. Also in wheat, TAR transcription is affected by N-supply [224]. Not much is known about the mechanism by which N-availability regulates auxin biosynthesis genes. The overexpression of AGAMOUS-like 21 (AGL 21) has been shown to stimulate LR elongation as a function of N-availability in agreement with enhanced expression of YUC5, YUC8 and TAR3 [225]. An agl21 mutant had fewer and shorter LRs and exogenous auxin application rescued the root phenotype, pointing to a role in the regulation of auxin biosynthesis. In Arabidopsis, at low concentrations of phosphate, PR growth is impaired, while LR development and root hair formation are promoted [226]. The working hypothesis is that auxin accumulation is required to ameliorate root growth under phosphate-starving conditions and overexpression of YUC1 promotes root architecture changes in P-limiting conditions [227]. NFYA (NUCLEAR FACTOR Y A)-B1 responds to nitrate- and phosphate-starvation by modulating the expression of TAR2 [228]. Finally, sulphate-availability modulates transcriptional activation of nitrilase 3 (NIT3) and this leads to enhanced local auxin biosynthesis [229].
Gravitropism ensures roots to grow downward in their search of sufficient nutrients and water [230]. In the Arabidopsis root, an asymmetric auxin gradient between the lower and upper sides of roots is responsible for root curving in response to gravity [231,232]. Besides a differential auxin transport, differential TAA1 activation in the lower and upper epidermal cells, aids in the formation of an asymmetric auxin gradient and response to the gravity vector [26].
The PR elongates under mild drought conditions, allowing access to deeper water reserves and this growth is supported by modulating auxin activity in the PR meristem. Transgenic plants (Arabidopsis, potato and poplar) overexpressing iaaM, YUC6 or YUC7, produce more IAA and show enhanced drought resistance, while the yuc1/yuc2/yuc6 triple mutant is more sensitive to drought stress [233,234,235,236,237]. However, co-overexpression of iaaL, an IAA-conjugating enzyme and YUC6, reduces the endogenous IAA-levels but drought stress tolerance remained unaffected, which indicated that the tolerance is not solely based on IAA-overproduction or -levels [233]. Also, IAA-Ala Resistant3 (IAR3) plays a role in modulating root architecture during osmotic stress. This hydrolase is capable of generating free IAA from IAA-amino acid conjugates under drought stress. The locally generated auxin then stimulates LR development to promote plant survival in dry conditions [238]. Water availability stimulates auxin biosynthesis and -transport via the regulation of TAA1 and PIN3. Newly initiated LR are preferentially initiated towards the water gradient and this as well might be mediated by local auxin-biosynthesis [239]. This hydro-patterning is reduced in wei8 and sav3 but can be rescued by constitutive expression of TAA1 in the root epidermis [240].
Aluminum (Al), when present as Al3+ ion, constitutes a major treat to the survival of plants in acidic soils (pH < 5) [241,242]. Al-induced inhibition of root growth is mediated by auxin [243,244,245]. In Arabidopsis, Al induced TAA1 directly or through the intervention of ethylene and as a consequence inhibited root growth [245,246]. Similarly, auxin-overproducing yucca mutants had an increased Al-sensitivity and contributed to the root growth inhibition in response to Al-stress [247,248]. Besides an effect on auxin biosynthesis, a reduction of PIN2 expression and localization contributed to the Al-induced root inhibition [249,250]. The metal ion of manganese (Mn) also inhibited PR growth and LR development by reducing the expression of YUC2, YUC3, SUR1, ASA1, PIN4 and PIN7 [251]. Boron-deprivation led to a significant increase of the endogenous auxin content thanks to the induction of TAA1, TAR2, YUC3 and YUC8 together with a decrease of the rootward IAA-transport (mediated by PIN1 and PIN4) [252].
In addition to abiotic environmental factors, root growth is influenced by beneficial growth-promoting rhizobacteria (PGPR) or fungi (PGPF) growing in the vicinity, on or in root tissue and pathogens [253]. The growth hormone auxin plays an important role in the communication between these microbes and the plant. Many plant-pathogenic microorganisms produce indole-3-acetic acid (IAA), which often determines their pathogenicity or influence endogenous auxin biosynthesis [254,255,256]. In general, both abiotic and biotic stresses and environmental conditions affect root development through the modulation of auxin biosynthesis.
8. Meta- and Catabolic Processes Controlling Auxin Levels
For auxin to display its crucial role during plant development, the endogenous concentration has to be spatio-temporally regulated. The role of biosynthesis has been discussed above but also inactivation, metabolism and storage contribute to the establishment of local levels of active auxin. IAA is known to act as a free acid but also conjugated forms, with a wide variety of sugars, peptides and amino acids are found in plants [17,18,65,257].
Conjugation of IAA occurs predominantly at the carboxyl group involving amino acids, sugars and other primary metabolites in order to render IAA inactive [19]. Conjugation could reversibly modulate free IAA levels and hence allows rapid and versatile responses to environmental changes and developmental [258,259,260]. IAA–amino acid conjugates can be classified into two groups based on in vitro activity and in planta feeding assays. Conjugation of IAA to aspartate (Asp), cysteine (Cys), histidine (His), isoleucine (Ile), lysine (Lys), proline (Pro), tryptophan (Trp) and valine (Val), various alcohols and sugars, serves as a way to inactivate IAA and to label it for degradation [255,256,258,259,260], whereas conjugation to alanine (Ala), leucine (Leu), phenylalanine (Phe), asparagine (Asn), glutamine-(Gln), glutamic acid (Glu), glycine (Gly), methionine (Met), serine (Ser), threonine (Thr) and tyrosine (Tyr), myo-inositol, or peptides creates temporary storage forms that can generate free IAA by hydrolysis [21,259,260,261,262,263]. Not only exogenous 2,4-d but also its amide-linked metabolite 2,4-d-Glu displayed an inhibitory effect on plant growth via the TIR1/AFB auxin-mediated signaling pathway and this metabolite can be found endogenously in the 2,4-d-treated plants [264]. At least seven members of the GRETCHEN HAGEN 3 (GH3) protein family have been shown to be involved in IAA-conjugation to amino acids and their expression is regulated by free auxin [20,265] (Purple pathway Figure 1). The development of adenosine-5′ (2-(1H-indol-3-yl) ethyl) phosphate (AIEP), a small molecule inhibitor of IAA-amido synthase, has helped the functional analysis of IAA-amido synthases [266]. The hydrolysis of IAA-conjugates to free IAA is regulated by IAA-LEUCINE RESISTANT 1 (ILR1), IAA-LEUCINE RESISTANT-LIKE 2 (ILL2) and IAA-ALANINE RESISTANT 3 (IAR3) [257,267,268,269] (Purple pathway Figure 1). The inactive IAA-methyl ester, Me-IAA, is generated by IAA CAROXYMETHYLTRANSFERASE1 (IAMT1) and hydrolyzed by METHYL ESTERASE17 (MES17) (Purple pathway Figure 1). Me-IAA has been shown to play a role in gravitropism, it exerts, like IAA, an inhibitory activity on root and hypocotyl growth, leaf development and is involved in plant fertility because RNAi IAMT1 plants form less seeds [270,271]. However, MeIAA does not function by itself but exogenous application results in in planta hydrolysis and release of free IAA. Interestingly, blocking the conversion of auxin storage forms to free IAA results in a compensating activity of the IPyA biosynthetic pathway [272].
IBA can be considered a precursor of IAA. IBA is metabolized slower than other conjugates and less prone to non-enzymatic degradation. Furthermore, IBA does not undergo oxidation [258]. IBA is transported in the root, probably by the carrier proteins PLEIOTROPIC DRUG RESISTANCE 8 (PDR8) and PDR9 [6,58]. IBA-activity depends on peroxisomal import through PEROXISOMAL ABC TRANSPORTER1 (PXA1) and subsequent β-oxidation to IAA. The peroxisomal enzyme 3-ketoacyl-CoA thiolase PED, contributes to both fatty acid and IBA β-oxidation. Despite ample reports on bioactivity of IBA, its biological role is still controversial [273,274] (Purple pathway Figure 1).
The inactivation of auxin by oxidation resulting in the formation of the catabolite 2-oxindole-3-acetic acid (oxIAA) has only recently been studied in more detail but was already longtime considered a main auxin catabolism pathway [275,276]. oxIAA is an irreversible IAA catabolite with little bio-activity and it is not transported via the canonical polar auxin transport system [277]. Recently, three groups independently published back to back on the role of the Arabidopsis gene DIOXYGENASE FOR AUXIN OXIDATION 1 (AtDAO1) to catalyze the formation of oxIAA both in vitro and in vivo [278,279,280] (Purple pathway Figure 1). DAO was previously characterized in rice to have a role in auxin metabolism and reproductive development [281]. Arabidopsis mutants lacking this enzyme contain 50–95% less oxIAA (resp. data from [20,279]) and have phenotypes reminiscent of elevated auxin levels [279,280]. A second isoform, AtDAO2, is only expressed in the root cap and is also involved in the formation of oxIAA. Both genes are closely related to the apple Adventitious Rooting Related Oxygenase 1 (ARRO1) and rice DAO [280,282,283]. Temporal and tissue-specific inactivation of auxin by AtDAO1 is essential to regulate the plant’s endogenous auxin levels. The localization of AtDAO1 in the border cells flanking the lateral root primordia (LRP), suggests that this enzyme is important to create an auxin maximum at the center of the root primordium [279]. In mutants lacking AtDAO1, the expression of GH3s as well as the synthesis of IAA-Asp and IAA-Glu is upregulated. This suggests that there is a compensation feedback between the different redundant IAA-regulating pathways [20,279,280]. In plants, there is a fast auxin conjugation response through the formation of conjugates such as IAA-Asp, which is more active at higher auxin concentrations and a constitutive slow conjugation response through oxidation and oxIAA formation, which acts at lower auxin concentrations [20,279]. The increase in auxin production in the dao1 mutant is mainly mediated via the IPyA pathway [20,280]. AtDAO2 was shown to follow a circadian rhythm-based oscillation in the root cap, which is a trigger for the activation of lateral root initials [58,59,281].
The generation of oxIAA is followed by a subsequent glycosylation to oxIAA-glucoside (oxIAA-Glc), which is very efficiently executed by UGT74D1 [284] (Purple pathway Figure 1). UDP-glycosyltransferases (UGTs) are one of the largest families of glycosyltransferases in plants [281,283] and some of them act in the conversion of IAA to glucose-conjugates. UGT84B1 catalyzes the conversion of IAA to 1-O-(indol-3-ylacetyl)-β-d-glucose (IAA-Glc), while UGT74E2 catalyzes the formation of 1-O-(indol-3-ylbutanoyl)-β-d-glucose (IBA-Glc) from indole-3-butylic acid (IBA) and UGT74D1 converts both IAA and IBA but also NAA, 2,4-D and ICA to their corresponding glucosides [285,286,287]. The iaglu gene encodes IAA glucosyltransferase in Zea mays [288]. In Arabidopsis over 100 uridine diphosphate (UDP)-glycosyltransferases (UGTs) have been identified that are classified into 14 subgroups [289,290]. Members of the L subgroup are potentially involved in the formation of IAA-glucose esters. Overexpression of UGT74D1 results in the loss of root gravitropism presumably due to increased glycosylation of ox-IAA and reduced IAA level at the root tip [284]. At the whole plant level, no decrease in IAA was observed, which suggest a compensating feedback mechanism [284]. A similar phenomenon was observed in plants overexpressing UGT84B1 in which an increase of IAA was measured [291]. The overexpression of UGT74E2 on the other hand led to an increase in IBA [287]. To allow for a temporal- and concentration-dependent regulation of auxin homeostasis, it was shown that GH3s act fast while UGT74D1 was slower upon auxin treatment. These results suggest that the OxIAA and GH3-pathways have distinct roles in IAA-homeostasis. The OxIAA pathway may function constitutively to maintain the basal levels of IAA in plants while the GH3 pathway may play a role in cases where plant cells have to rapidly reduce the relative amount of IAA in response to developmental and environmental changes [284].
Last but not lease, conjugation compartmentalization of IAA in the endoplasmic reticulum has been shown to regulate the availability of IAA to cellular conjugation mechanisms [292].
9. Conclusions
What have we learned so far from auxin-related studies is that the combined action of auxin biosynthesis and transport set the stage for adaptive developmental processes. The root system in particular is sensitive to auxin. Whereas the last decade’s emphasis was more on auxin redistribution by means of polar and local transport, current research progress underscores the importance of local auxin biosynthesis. Auxin biosynthesis genes have been found to play a role in every aspect of root development translating environmental cues into adaptive responses (see Table 1). As auxin, derived from distantly located apical origins as well as locally synthesized auxin equally have impact on root architecture and development, the mechanism by which auxin is perceived and possibly how it enters the cells may hold specific information relevant for the adaptive growth responses. Future research should focus on local auxin signaling and spatial and temporal perception mechanisms controlling auxin-mediated establishment of root meristems.
Author Contributions
Damilola Olatunji contributed to the manuscript by writing, figure ideas and editing, Danny Geelen created the review’s outline and edited the manuscript, Inge Verstraeten wrote the manuscript, designed the figures and edited.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 2,4-d | 2,4-dichlorophenoxy acetic acid |
| 4-Cl-IAA | 4-chloroindole-3-acetic acid |
| ABP1 | auxin binding protein 1 |
| AFB | AUXIN SIGNALING F-BOX (AFB) family of auxin receptors |
| AO | aldehyde oxidases |
| AR(s) | adventitious root(s) |
| ARF | auxin response factor |
| Aux/IAA | AUXIN/INDOLE-3-ACETIC ACID proteins |
| IAA | indole-3-acetic acid |
| IAD | indole-3-acetaldehyde |
| IAM | indole-3-acetamide |
| IAN | indole-3-acetonitrile |
| IAOx | indole-3-acetaldoxime |
| IBA | indole-3-butyric acid |
| IG | indole glucosinolates |
| IGS | indole-3-glycerol phosphate synthase |
| IPyA | indole-3-pyruvic acid |
| LR(s) | lateral root(s) |
| NAA | 1-naphthaleneacetic acid |
| PAA | phenyl acetic acid |
| PIN | PIN-formed family of auxin transporters |
| PHYB | PHYTOCHROME B |
| PR | primary root |
| QC | quiescent center |
| RAM | root apical meristem |
| SAM | shoot apical meristem |
| SCF | ubiquitin protein ligase complex, with subunits: Skp1, Cullin and F-box |
| TAA1/TAR1-4 | TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1/TRYPTOPHAN AMINOTRANSFERASE RELATED 1-4 |
| TAM | Tryptamine |
| TIR1/2 | TRANSPORT INHIBITOR RESPONSE ½ |
| Trp | Tryptophan |
| TSA1 | Trp synthase a |
| TSB | Trp synthase b |
| YUC1-11 | yucca genes encoding flavin-containing monooxygenases |
References
- Palmer, C.M.; Bush, S.M.; Maloof, J.N. Phenotypic and Developmental Plasticity in Plants; Encyclopedia of Life Science, John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Gaillochet, C.; Lohmann, J.U. The never-ending story: From pluripotency to plant developmental plasticity. Development 2015, 142, 2237–2249. [Google Scholar] [CrossRef] [PubMed]
- Stahl, Y.; Simon, R. Plant primary meristems: Shared functions and regulatory mechanisms. Curr. Opin. Plant Biol. 2010, 13, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Wendrich, J.R.; Möller, B.K.; Li, S.; Saiga, S.; Sozzani, R.; Benfey, P.N.; De Rybel, B.; Weijers, D. Framework for gradual progression of cell ontogeny in the Arabidopsis root meristem. Proc. Natl. Acad. Sci. USA 2017, 111, E8922–E8929. [Google Scholar] [CrossRef] [PubMed]
- Kenrick, P.; Strullu-Derrien, C. The origin and early evolution of roots. Plant Physiol. 2014, 166, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef] [PubMed]
- Petricka, J.J.; Winter, C.M.; Benfey, P.N. Control of Arabidopsis root development. Annu. Rev. Plant Biol. 2012, 63, 563–590. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; De Smet, I. Root system architecture: Insights from Arabidopsis and cereal crops. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2012, 367, 1441–1452. [Google Scholar] [CrossRef] [PubMed]
- Scheres, B.; Wolkenfelt, H.; Willemsen, V.; Terlouw, M.; Lawson, E.; Dean, C.; Weisbeek, P. Embryonic origin of the Arabidopsis primary root and root-meristem initials. Development 1994, 120, 2475–2487. [Google Scholar]
- Bellini, C.; Pacurar, D.I.; Perrone, I. Adventitious roots and lateral roots: Similarities and differences. Annu. Rev. Plant Biol. 2014, 65, 639–666. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, I.; Schotte, S.; Geelen, D. Hypocotyl adventitious root organogenesis differs from lateral root development. Front. Plant Sci. 2014, 5, 495. [Google Scholar] [PubMed]
- Coudert, P.; van Anh Le, T.; Gantet, P. Rice: A model plant to decipher the hidden origin of adventitious roots. In Plant Roots: The Hidden Half; Beeckman, T., Ed.; CRC Press: Boca Raton, FL, USA, 2013. [Google Scholar]
- Palme, K.; Hesse, T.; Moore, I.; Campos, N.; Feldwisch, J.; Garbers, C.; Hesse, F.; Schell, J. Hormonal modulation of plant growth: The role of auxin perception. Mech. Dev. 1991, 33, 97–106. [Google Scholar] [CrossRef]
- Benkova, E.; Hejatko, J. Hormone interactions at the root apical meristem. Plant Mol. Biol. 2009, 69, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Peer, W.A.; Blakeslee, J.J.; Yang, H.; Murphy, A.S. Seven things we think we know about auxin transport. Mol. Plant 2011, 4, 487–504. [Google Scholar] [CrossRef] [PubMed]
- Zazimalová, E.; Murphy, A.S.; Yang, H.; Hoyerová, K.; Hošek, P. Auxin transporters—Why so many? Cold Spring Harb. Perspect. Biol. 2010, 2, a001552. [Google Scholar] [CrossRef] [PubMed]
- Korasick, D.A.; Enders, T.A.; Strader, L.C. Auxin biosynthesis and storage forms. J. Exp. Bot. 2013, 64, 2541–2555. [Google Scholar] [CrossRef] [PubMed]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Ludwig-Muller, J. Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 2000, 32, 219–230. [Google Scholar] [CrossRef]
- Mellor, N.; Band, L.R.; Pencik, A.; Novak, O.; Rashed, A.; Holman, T.; Wilson, M.H.; Voss, U.; Bishopp, A.; King, J.R.; et al. Dynamic regulation of auxin oxidase and conjugating enzymes AtDAO1 and GH3 modulates auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 11022–11027. [Google Scholar] [CrossRef] [PubMed]
- Rosquete, M.R.; Barbez, E.; Kleine-Vehn, J. Cellular auxin homeostasis: Gatekeeping is housekeeping. Mol. Plant 2012, 5, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Rozov, S.; Zagorskaya, A.; Deineko, E.; Shumny, V. Auxins: Biosynthesis, metabolism, and transport. Biol. Bull. Rev. 2013, 3, 286–295. [Google Scholar] [CrossRef]
- Kramer, E.M.; Ackelsberg, E.M. Auxin metabolism rates and implications for plant development. Front. Plant Sci. 2015, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.D.W.; Cho, H.T. Auxin, the organizer of the hormonal/environmental signals for root hair growth. Front. Plant Sci. 2013, 4, 448. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.N.; Robertson-Hoyt, J.; Yun, J.; Benavente, L.M.; Xie, D.Y.; Dolezal, K.; Schlereth, A.; Jurgens, G.; Alonso, J.M. TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell 2008, 133, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Greenham, K.; Prigge, M.J.; Jensen, P.J.; Estelle, M. The TRANSPORT INHIBITOR RESPONSE2 gene is required for auxin synthesis and diverse aspects of plant development. Plant Physiol. 2009, 151, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. Auxin biosynthesis: The Arabidopsis Book. BioOne 2014, 12, e0173. [Google Scholar] [CrossRef] [PubMed]
- Band, L.R.; Wells, D.M.; Fozard, J.A.; Ghetiu, T.; French, A.P.; Pound, M.P.; Wilson, M.H.; Yu, L.; Li, W.D.; Hijazi, H.I.; et al. Systems analysis of auxin transport in the Arabidopsis root apex. Plant Cell 2014, 26, 862–875. [Google Scholar] [CrossRef] [PubMed]
- Benkova, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertova, D.; Jurgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef]
- Petrasek, J.; Friml, J. Auxin transport routes in plant development. Development 2009, 136, 2675–2688. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Skupa, P.; Viaene, T.; Zwiewka, M.; Tejos, R.; Klima, P.; Carna, M.; Rolcik, J.; De Rycke, R.; Moreno, I.; et al. PIN6 auxin transporter at endoplasmic reticulum and plasma membrane mediates auxin homeostasis and organogenesis in Arabidopsis. New Phytol. 2016, 211, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Swarup, K.; Benkova, E.; Swarup, R.; Casimiro, I.; Peret, B.; Yang, Y.; Parry, G.; Nielsen, E.; De Smet, I.; Vanneste, S.; et al. The auxin influx carrier LAX3 promotes lateral root emergence. Nat. Cell Biol. 2008, 10, 946–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargmann, B.O.; Estelle, M. Auxin perception: In the IAA of the beholder. Physiol. Plant 2014, 151, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Farcot, E.; Lavedrine, C.; Vernoux, T. A modular analysis of the auxin signalling network. PLoS ONE 2015, 10, e0122231. [Google Scholar] [CrossRef] [PubMed]
- Strader, L.C.; Zhao, Y.D. Auxin perception and downstream events. Curr. Opin. Plant Biol. 2016, 33, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Lavy, M.; Estelle, M. Mechanisms of auxin signaling. Development 2016, 143, 3226–3229. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin signaling. Plant Physiol. 2017, 175. [Google Scholar] [CrossRef] [PubMed]
- Adamowski, M.; Friml, J. PIN-dependent auxin transport: Action, regulation, and evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Maraschin, F.; Memelink, J.; Offringa, R. Auxin-induced, SCFTIR1-mediated poly-ubiquitination marks AUX/IAA proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Dharmasiri, N.; Dharmasiri, S.; Weijers, D.; Lechner, E.; Yamada, M.; Hobbie, L.; Ehrismann, J.S.; Jürgens, G.; Estelle, M. Plant development is regulated by a family of auxin receptor F box proteins. Dev. Cell 2005, 9, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Prigge, M.J.; Greenham, K.; Zhang, Y.; Santner, A.; Castillejo, C.; Mutka, A.M.; O’Malley, R.C.; Ecker, J.R.; Kunkel, B.N.; Estelle, M. The Arabidopsis auxin receptor F-Box proteins AFB4 and AFB5 are required for response to the synthetic auxin picloram. G3 2016, 6, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Paponov, I.A.; Paponov, M.; Teale, W.; Menges, M.; Chakrabortee, S.; Murray, J.A.H.; Palme, K. Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 2008, 1, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Vernoux, T.; Brunoud, G.; Farcot, E.; Morin, V.; Van den Daele, H.; Legrand, J.; Oliva, M.; Das, P.; Larrieu, A.; Wells, D.; et al. The auxin signalling network translates dynamic input into robust patterning at the shoot apex. Mol. Syst. Biol. 2011, 7, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderon Villalobos, L.I.A.; Lee, S.; De Oliveira, C.; Ivetac, A.; Brandt, W.; Armitage, L.; Sheard, L.B.; Tan, X.; Parry, G.; Mao, H.; et al. A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 2012, 8, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Jones, A.R. Endoplasmic reticulum: The rising compartment in auxin biology. Plant Physiol. 2010, 154, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Simon, S.; Petrášek, J. Why plants need more than one type of auxin. Plant Sci. 2011, 180, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Kim, J.Y. Revisiting apoplastic auxin signaling mediated by AUXIN BINDING PROTEIN 1. Mol. Cell 2015, 38, 829–835. [Google Scholar]
- Gao, Y.; Zhang, Y.; Zhang, D.; Dai, X.; Estelle, M.; Zhao, Y. Auxin binding protein 1 (ABP1) is not required for either auxin signaling or Arabidopsis development. Proc. Natl. Acad. Sci. USA 2015, 112, 2275–2280. [Google Scholar] [CrossRef] [PubMed]
- Dahlke, R.I.; Fraas, S.; Ullrich, K.K.; Heinemann, K.; Romeiks, M.; Rickmeyer, T.; Klebe, G.; Palme, K.; Luthen, H.; Steffens, B. Protoplast swelling and hypocotyl growth depend on different auxin signaling pathways. Plant Physiol. 2017, 175, 982–994. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.D.; Ross, J.J. The auxins, IAA and PAA, are synthesized by similar steps catalyzed by different enzymes. Plant Signal. Behav. 2016, 11, e1250993. [Google Scholar] [CrossRef] [PubMed]
- Enders, T.A.; Strader, L.C. Auxin activity: Past, present, and future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, S.; Mashiguchi, K.; Tanaka, K.; Hishiyama, S.; Sakai, T.; Hanada, K.; Kinoshita-Tsujimura, K.; Yu, H.; Dai, X.H.; Takebayashi, Y.; et al. Distinct characteristics of indole-3-acetic acid and phenylacetic acid, two common auxins in plants. Plant Cell Physiol. 2015, 56, 1641–1654. [Google Scholar] [CrossRef] [PubMed]
- Strader, L.C.; Wheeler, D.L.; Christensen, S.E.; Berens, J.C.; Cohen, J.D.; Rampey, R.A.; Bartel, B. Multiple facets of Arabidopsis seedling development require indole-3-butyric acid-derived auxin. Plant Cell 2011, 23, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Zolman, B.K.; Nyberg, M.; Bartel, B. IBR3, a novel peroxisomal acyl-CoA dehydrogenase-like protein required for indole-3-butyric acid response. Plant Mol. Biol. 2007, 64, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Zolman, B.R.; Martinez, N.; Millius, A.; Adham, A.R.; Bartel, B. Identification and characterization of Arabidopsis indole-3-butyric acid response mutants defective in novel peroxisomal enzymes. Genetics 2008, 180, 237–251. [Google Scholar] [CrossRef] [PubMed]
- De Rybel, B.; Audenaert, D.; Xuan, W.; Overvoorde, P.; Strader, L.C.; Kepinski, S.; Hoye, R.; Brisbois, R.; Parizot, B.; Vanneste, S.; et al. A role for the root cap in root branching revealed by the non-auxin probe naxillin. Nat. Chem. Biol. 2012, 8, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Audenaert, D.; Parizot, B.; Möller, B.K.; Njo, M.F.; De Rybel, B.; De Rop, G.; Van Isterdael, G.; Mähönen, A.P.; Vanneste, S.; et al. Root cap-derived auxin pre-patterns the longitudinal axis of the Arabidopsis root. Curr. Biol. 2015, 25, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Strader, L.C.; Nemhauser, J.L. Auxin 2012: A rich mea ho’oulu. Development 2013, 140, 1153–1157. [Google Scholar] [CrossRef] [PubMed]
- Bennett, T.; Leyser, O. The auxin question: A philosophical overview. In Auxin and Its Role in Plant Development; Zazimalova, E., Petrasek, J., Benkova, E., Eds.; Springer: Wien, Austria, 2014; pp. 3–19. [Google Scholar]
- Petersson, S.V.; Johansson, A.I.; Kowalczyk, M.; Makoveychuk, A.; Wang, J.Y.; Moritz, T.; Grebe, M.; Benfey, P.N.; Sandberg, G.; Ljung, K. An auxin gradient and maximum in the Arabidopsis root apex shown by high-resolution cell-specific analysis of IAA distribution and synthesis. Plant Cell 2009, 21, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Niu, T.; Yu, Q.; Quan, T.; Ding, Z. Auxin gradient is crucial for the maintenance of root distal stem cell identity in Arabidopsis. Plant Signal. Behav. 2013, 8, e26429. [Google Scholar] [CrossRef] [PubMed]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Ferrer, J.-L.; Ljung, K.; Pojer, F.; Hong, F.; Long, J.A.; Li, L.; Moreno, J.E.; Bowman, M.E.; Ivans, L.J. Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants. Cell 2008, 133, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Ljung, K.; Bhalerao, R.P.; Sandberg, G. Sites and homeostatic control of auxin biosynthesis in Arabidopsis during vegetative growth. Plant J. 2001, 28, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Ljung, K.; Hull, A.K.; Celenza, J.; Yamada, M.; Estelle, M.; Normanly, J.; Sandberg, G. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell 2005, 17, 1090–1104. [Google Scholar] [CrossRef] [PubMed]
- Bhalerao, R.P.; Eklof, J.; Ljung, K.; Marchant, A.; Bennett, M.; Sandberg, G. Shoot-derived auxin is essential for early lateral root emergence in Arabidopsis seedlings. Plant J. 2002, 29, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Renton, M.; Hanan, J.; Ferguson, B.J.; Beveridge, C.A. Models of long-distance transport: How is carrier-dependent auxin transport regulated in the stem? New Phytol. 2012, 194, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, T.; Heskett, M.; Wilson, E. Microbial synthesis and degradation of indole-3-acetic acid I. The conversion of L-tryptophan to indole-3-acetamide by an enzyme system from Pseudomonas savastanoi. J. Biol. Chem. 1966, 241, 3738–3744. [Google Scholar] [PubMed]
- Klee, H.J.; Horsch, R.B.; Hinchee, M.A.; Hein, M.B.; Hoffmann, N.L. The effects of overproduction of two Agrobacterium tumefaciens T-DNA auxin biosynthetic gene products in transgenic petunia plants. Genes Dev. 1987, 1, 86–96. [Google Scholar] [CrossRef]
- Badenoch-Jones, J.; Summons, R.E.; Djordjevic, M.A.; Shine, J.; Letham, D.S.; Rolfe, B.G. Mass spectrometric quantification of indole-3-acetic acid in Rhizobium culture supernatants: Relation to root hair curling and nodule initiation. Appl. Environ. Microbiol. 1982, 44, 275–280. [Google Scholar] [PubMed]
- Badenoch-Jones, J.; Summons, R.E.; Entsch, B.; Rolfe, B.G.; Parker, C.W.; Letham, D.S. Mass spectrometric identification of indole compounds produced by Rhizobium strains. Biol. Mass Spectrom. 1982, 9, 429–437. [Google Scholar] [CrossRef]
- Koga, J.; Adachi, T.; Hidaka, H. Purification and characterization of indolepyruvate decarboxylase. A novel enzyme for indole-3-acetic acid biosynthesis in Enterobacter cloacae. J. Biol. Chem. 1992, 267, 15823–15828. [Google Scholar] [PubMed]
- Wright, A.D.; Sampson, M.B.; Neuffer, M.G.; Michalczuk, L.; Slovin, J.P.; Cohen, J.D. Indole-3-acetic acid biosynthesis in the mutant maize orange pericarp, a tryptophan auxotroph. Science 1991, 254, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J.; Cohen, J.D.; Fink, G.R. Arabidopsis thaliana auxotrophs reveal a tryptophan-independent biosynthetic pathway for indole-3-acetic acid. Proc. Natl. Acad. Sci. USA 1993, 90, 10355–10359. [Google Scholar] [CrossRef] [PubMed]
- Radwanski, E.R.; Last, R.L. Tryptophan biosynthesis and metabolism: Biochemical and molecular genetics. Plant Cell 1995, 7, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Shao, X.; Li, J. Indole-3-glycerol phosphate, a branchpoint of indole-3-acetic acid biosynthesis from the tryptophan biosynthetic pathway in Arabidopsis thaliana. Plant J. 2000, 24, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef] [PubMed]
- Last, R.L.; Fink, G.R. Tryptophan-requiring mutants of the plant Arabidopsis thaliana. Science 1988, 240, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Estelle, M.A.; Somerville, C. Auxin-resistant mutants of Arabidopsis thaliana with an altered morphology. Mol. Genet. Genom. 1987, 206, 200–206. [Google Scholar] [CrossRef]
- Cooney, T.P.; Nonhebel, H.M. The measurement and mass-spectral identification of indole-3-pyruvate from tomato shoots. Biochem. Biophys. Res. Commun. 1989, 162, 761–766. [Google Scholar] [CrossRef]
- Tivendale, N.D.; Ross, J.J.; Cohen, J.D. The shifting paradigms of auxin biosynthesis. Trends Plant Sci. 2014, 19, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhao, Y. Auxin biosynthesis and catabolism. In Auxin and Its Role in Plant Development; Zazimalova, E., Petrasek, J., Benkova, E., Eds.; Springer: Wien, Austria, 2014; pp. 21–38. [Google Scholar]
- Stepanova, A.N.; Yun, J.; Robles, L.M.; Novak, O.; He, W.R.; Guo, H.W.; Ljung, K.; Alonso, J.M. The Arabidopsis YUCCA1 flavin monooxygenase functions in the indole-3-pyruvic acid branch of auxin biosynthesis. Plant Cell 2011, 23, 3961–3973. [Google Scholar] [CrossRef] [PubMed]
- Won, C.; Shen, X.L.; Mashiguchi, K.; Zheng, Z.Y.; Dai, X.H.; Cheng, Y.F.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y.D. Conversion of tryptophan to indole-3-acetic acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18518–18523. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. Auxin biosynthesis: A simple two-step pathway converts tryptophan to indole-3-acetic acid in plants. Mol. Plant 2012, 5, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Tivendale, N.D.; Davidson, S.E.; Davies, N.W.; Smith, J.A.; Dalmais, M.; Bendahmane, A.I.; Quittenden, L.J.; Sutton, L.; Bala, R.K.; Le Signor, C.; et al. Biosynthesis of the halogenated auxin, 4-chloroindole-3-acetic acid. Plant Physiol. 2012, 159, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.H.; Mashiguchi, K.; Chen, Q.G.; Kasahara, H.; Kamiya, Y.; Ojha, S.; DuBois, J.; Ballou, D.; Zhao, Y.D. The biochemical mechanism of auxin biosynthesis by an Arabidopsis YUCCA flavin-containing monooxygenase. J. Biol. Chem. 2013, 288, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed]
- Gallavotti, A.; Barazesh, S.; Malcomber, S.; Hall, D.; Jackson, D.; Schmidt, R.J.; McSteen, P. Sparse inflorescence 1 encodes a monocot-specific YUCCA-like gene required for vegetative and reproductive development in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 15196–15201. [Google Scholar] [CrossRef] [PubMed]
- Phillips, K.A.; Skirpan, A.L.; Liu, X.; Christensen, A.; Slewinski, T.L.; Hudson, C.; Barazesh, S.; Cohen, J.D.; Malcomber, S.; McSteen, P. Vanishing tassel 2 encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize. Plant Cell 2011, 23, 550–566. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.F.; Dai, X.H.; Zhao, Y.D. Auxin synthesized by the YUCCA flavin monooxygenases is essential for embryogenesis and leaf formation in Arabidopsis. Plant Cell 2007, 19, 2430–2439. [Google Scholar] [CrossRef] [PubMed]
- Mano, Y.; Nemoto, K. The pathway of auxin biosynthesis in plants. J. Exp. Bot. 2012, 63, 2853–2872. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Akaba, S.; Oritani, T.; Delarue, M.; Bellini, C.; Caboche, M.; Koshiba, T. Higher activity of an aldehyde oxidase in the auxin-overproducing superroot1 mutant of Arabidopsis thaliana. Plant Physiol. 1998, 116, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Novak, O.; Henykova, E.; Sairanen, I.; Kowalczyk, M.; Pospisil, T.; Ljung, K. Tissue-specific profiling of the Arabidopsis thaliana auxin metabolome. Plant J. 2012, 72, 523–536. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, S.; Müller, A.; Piotrowski, M.; Weiler, E.W. Occurrence and formation of indole-3-acetamide in Arabidopsis thaliana. Planta 2002, 216, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, S.; Hishiyama, S.; Jikumaru, Y.; Hanada, A.; Nishimura, T.; Koshiba, T.; Zhao, Y.; Kamiya, Y.; Kasahara, H. Biochemical analyses of indole-3-acetaldoxime-dependent auxin biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5430–5435. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, S.; Neu, D.; Weiler, E.W. Molecular cloning and characterization of an amidase from Arabidopsis thaliana capable of converting indole-3-acetamide into the plant growth hormone, indole-3-acetic acid. Phytochemistry 2003, 62, 293–300. [Google Scholar] [CrossRef]
- Nemoto, K.; Hara, M.; Suzuki, M.; Seki, H.; Muranaka, T.; Mano, Y. The NtAMI1 gene functions in cell division of tobacco BY-2 cells in the presence of indole-3-acetamide. FEBS Lett. 2009, 583, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Parra, B.; Frerigmann, H.; Alonso, M.-M.P.; Loba, V.C.; Jost, R.; Hentrich, M.; Pollmann, S. Characterization of four bifunctional plant IAM/PAM-amidohydrolases capable of contributing to auxin biosynthesis. Plants 2014, 3, 324–347. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, T.; Hoffmann, M.; Hentrich, M.; Pollmann, S. Indole-3-acetamide-dependent auxin biosynthesis: A widely distributed way of indole-3-acetic acid production? Eur. J. Cell Biol. 2010, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Novák, O.; Hradecká, V.; Pĕnčík, A.; Rolčík, J.; Strnad, M.; Van Staden, J. Endogenous cytokinins, auxins and abscisic acid in Ulva fasciata (Chlorophyta) and Dictyota humifusa (Phaeophyta): Towards understanding their biosynthesis and homoeostasis. Eur. J. Phycol. 2009, 44, 231–240. [Google Scholar] [CrossRef]
- Abu-Zaitoon, Y.; Aladaileh, S.; Tawaha, A.R.A. Contribution of the IAM pathway to IAA pool in developing rice grains. Braz. Arch. Biol. Technol. 2016, 59, e16150677. [Google Scholar] [CrossRef]
- Mikkelsen, M.D.; Hansen, C.H.; Wittstock, U.; Halkier, B.A. Cytochrome P450 CYP79B2 from Arabidopsis catalyzes the conversion of tryptophan to indole-3-acetaldoxime, a precursor of indole glucosinolates and indole-3-acetic acid. J. Biol. Chem. 2000, 275, 33712–33717. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D.; Hull, A.K.; Gupta, N.R.; Goss, K.A.; Alonso, J.; Ecker, J.R.; Normanly, J.; Chory, J.; Celenza, J.L. Trp-dependent auxin biosynthesis in Arabidopsis: Involvement of cytochrome P450s CYP79B2 and CYP79B3. Genes Dev. 2002, 16, 3100–3112. [Google Scholar] [CrossRef] [PubMed]
- Tivendale, N.D.; Davies, N.W.; Molesworth, P.P.; Davidson, S.E.; Smith, J.A.; Lowe, E.K.; Reid, J.B.; Ross, J.J. Reassessing the role of N-hydroxytryptamine in auxin biosynthesis. Plant Physiol. 2010, 154, 1957–1965. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.; Celenza, J.L. Indolic glucosinolates at the crossroads of tryptophan metabolism. Phytochem. Rev. 2009, 8, 25–37. [Google Scholar] [CrossRef]
- Bak, S.; Tax, F.E.; Feldmann, K.A.; Galbraith, D.W.; Feyereisen, R. CYP83B1, a cytochrome P450 at the metabolic branch point in auxin and indole glucosinolate biosynthesis in Arabidopsis. Plant Cell 2001, 13, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Douglas Grubb, C.; Zipp, B.J.; Ludwig-Müller, J.; Masuno, M.N.; Molinski, T.F.; Abel, S. Arabidopsis glucosyltransferase UGT74B1 functions in glucosinolate biosynthesis and auxin homeostasis. Plant J. 2004, 40, 893–908. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, M.D.; Naur, P.; Halkier, B.A. Arabidopsis mutants in the C-S lyase of glucosinolate biosynthesis establish a critical role for indole-3-acetaldoxime in auxin homeostasis. Plant J. 2004, 37, 770–777. [Google Scholar] [CrossRef] [PubMed]
- Nafisi, M.; Goregaoker, S.; Botanga, C.J.; Glawischnig, E.; Olsen, C.E.; Halkier, B.A.; Glazebrook, J. Arabidopsis cytochrome P450 monooxygenase 71A13 catalyzes the conversion of indole-3-acetaldoxime in camalexin synthesis. Plant Cell 2007, 19, 2039–2052. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.L.; Wang, M.; Han, B.Y.; Tan, D.G.; Sun, X.P.; Zhang, J.M. Arabidopsis myrosinase genes AtTGG4 and AtTGG5 are root-tip specific and contribute to auxin biosynthesis and root growth regulation. Int. J. Mol. Sci. 2016, 17, 892. [Google Scholar] [CrossRef] [PubMed]
- Bartel, B.; Fink, G.R. Differential regulation of an auxin-producing Nitrilase gene family in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1994, 91, 6649–6653. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J.; Grisafi, P.; Fink, G.R.; Bartel, B. Arabidopsis mutants resistant to the auxin effects of indole-3-acetonitrile are defective in the nitrilase encoded by the NIT1 gene. Plant Cell 1997, 9, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wang, B.; Ouyang, J.; Li, J.; Wang, Y. Arabidopsis indole synthase, a homolog of tryptophan synthase alpha, is an enzyme involved in the trp-independent indole-containing metabolite biosynthesis. J. Integr. Plant Biol. 2008, 50, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Eilbert, E.; Bolbol, A.; Lukowitz, W. Going mainstream: How is the body axis of plants first initiated in the embryo? Dev. Biol. 2016, 419, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Mironova, V.; Teale, W.; Shahriari, M.; Dawson, J.; Palme, K. The systems biology of auxin in developing embryos. Trends Plant Sci. 2017, 22, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Robert, H.S.; Grones, P.; Stepanova, A.N.; Robles, L.M.; Lokerse, A.S.; Alonso, J.M.; Weijers, D.; Friml, J. Local auxin sources orient the apical-basal axis in Arabidopsis embryos. Curr. Biol. 2013, 23, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Saiga, S.; Weijers, D. Auxin regulation of embryonic root formation. Plant Cell Physiol. 2013, 54, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Sauer, M.; Meurette, O.; Friml, J.; Ljung, K.; Sandberg, G.; Hooykaas, P.; Offringa, R. Maintenance of embryonic auxin distribution for apical-basal patterning by PIN-FORMED-dependent auxin transport in Arabidopsis. Plant Cell 2005, 17, 2517–2526. [Google Scholar] [CrossRef] [PubMed]
- Geldner, N.; Hamann, T.; Jürgens, G. Is there a role for auxin in early embryogenesis? Plant Growth Regul. 2000, 32, 187–191. [Google Scholar] [CrossRef]
- Friml, J.; Vieten, A.; Sauer, M.; Weijers, D.; Schwarz, H.; Hamann, T.; Offringa, R.; Jurgens, G. Efflux-dependent auxin gradients establish the apical-basal axis of Arabidopsis. Nature 2003, 426, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Benjamins, R.; Scheres, B. Auxin: The looping star in plant development. Annu. Rev. Plant Biol. 2008, 59, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Chen, J.; Yang, Z. Auxin regulation of cell polarity in plants. Curr. Opin. Plant Biol. 2015, 28, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Wabnik, K.; Robert, H.S.; Smith, R.S.; Friml, J. Modeling framework for the establishment of the apical-basal embryonic axis in plants. Curr. Biol. 2013, 23, 2513–2518. [Google Scholar] [CrossRef] [PubMed]
- Picciarelli, P.; Ceccarelli, N.; Paolicchi, F.; Calistri, G. Endogenous auxins and embryogenesis in Phaseolus coccineus. Aust. J. Plant Physiol. 2001, 28, 73–78. [Google Scholar] [CrossRef]
- Smit, M.E.; Weijers, D. The role of auxin signaling in early embryo pattern formation. Curr. Opin. Plant Biol. 2015, 28, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Möller, B.; Weijers, D. Auxin control of embryo patterning. Cold Spring Harb. Perspect. Biol. 2009, 1, a001545. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Umeda, M. Hormonal control of cell division and elongation along differentiation trajectories in roots. J. Exp. Bot. 2014, 65, 2633–2643. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chu, J.; Yu, T.; Xu, Q.; Sun, X.; Yuan, J.; Xiong, G.; Wang, G.; Wang, Y.; Li, J. Tryptophan-independent auxin biosynthesis contributes to early embryogenesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, 4821–4826. [Google Scholar] [CrossRef] [PubMed]
- Nibau, C.; Gibbs, D.J.; Coates, J.C. Branching out in new directions: The control of root architecture by lateral root formation. New Phytol. 2008, 179, 595–614. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Gutjahr, C.; Li, C.J.; Hochholdinger, F. Genetic control of lateral root formation in cereals. Trends Plant Sci. 2016, 21, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, S.; Beis, D.; Wolkenfelt, H.; Murfett, J.; Guilfoyle, T.; Malamy, J.; Benfey, P.; Leyser, O.; Bechtold, N.; Weisbeek, P. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root. Cell 1999, 99, 463–472. [Google Scholar] [CrossRef]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Benkova, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jurgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef]
- Grieneisen, V.A.; Xu, J.; Maree, A.F.M.; Hogeweg, P.; Scheres, B. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, M.; Liang, N.; Zheng, Y.; Yu, Q.; Wu, S. Symplastic communication spatially directs local auxin biosynthesis to maintain root stem cell niche in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 4005–4010. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.G.; Dai, X.H.; De-Paoli, H.; Cheng, Y.F.; Takebayashi, Y.; Kasahara, H.; Kamiya, Y.; Zhao, Y.D. Auxin overproduction in shoots cannot rescue auxin deficiencies in Arabidopsis roots. Plant Cell Physiol. 2014, 55, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Pinon, V.; Prasad, K.; Grigg, S.P.; Sanchez-Perez, G.F.; Scheres, B. Local auxin biosynthesis regulation by PLETHORA transcription factors controls phyllotaxis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Ito, M.; Sumikura, T.; Nakayama, A.; Nishimura, T.; Kitano, H.; Yamaguchi, I.; Koshiba, T.; Hibara, K.I.; Nagato, Y.; et al. The rice FISH BONE gene encodes a tryptophan aminotransferase, which affects pleiotropic auxin-related processes. Plant J. 2014, 78, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin biosynthesis by the YUCCA genes in rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Sazuka, T.; Kamiya, N.; Nishimura, T.; Ohmae, K.; Sato, Y.; Imamura, K.; Nagato, Y.; Koshiba, T.; Nagamura, Y.; Ashikari, M.; et al. A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 2009, 60, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xie, W.F.; Zhang, L.; Valpuesta, V.; Ye, Z.W.; Gao, Q.H.; Duan, K. Auxin biosynthesis by the YUCCA6 flavin monooxygenase gene in woodland strawberry. J. Integr. Plant Biol. 2014, 56, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xiong, L. Localized auxin biosynthesis and postembryonic root development in Arabidopsis. Plant Signal. Behav. 2009, 4, 752–754. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, T.; Janowitz, T.; Sanchez-Parra, B.; Alonso, M.M.P.; Trompetter, I.; Piotrowski, M.; Pollmann, S. Arabidopsis NITRILASE 1 contributes to the regulation of root growth and development through modulation of auxin biosynthesis in seedlings. Front. Plant Sci. 2017, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Beeckman, T.; Burssens, S.; Inze, D. The peri-cell-cycle in Arabidopsis. J. Exp. Bot. 2001, 52, 403–411. [Google Scholar] [PubMed]
- De Smet, I.; Tetsumura, T.; De Rybel, B.; Frei dit Frey, N.; Laplaze, L.; Casimiro, I.; Swarup, R.; Naudts, M.; Vanneste, S.; Audenaert, D.; et al. Auxin-dependent regulation of lateral root positioning in the basal meristem of Arabidopsis. Development 2007, 134, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M. Lateral root initiation is a probabilistic event whose frequency is set by fluctuating levels of auxin response. J. Exp. Bot. 2013, 64, 2609–2617. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Risueno, M.A.; Van Norman, J.M.; Moreno, A.; Zhang, J.Y.; Ahnert, S.E.; Benfey, P.N. Oscillating gene expression determines competence for periodic Arabidopsis root branching. Science 2010, 329, 1306–1311. [Google Scholar] [CrossRef] [PubMed]
- Van Norman, J.M.; Xuan, W.; Beeckman, T.; Benfey, P.N. To branch or not to branch: The role of pre-patterning in lateral root formation. Development 2013, 140, 4301–4310. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Band, L.R.; Kumpf, R.P.; Van Damme, D.; Parizot, B.; De Rop, G.; Opdenacker, D.; Möller, B.K.; Skorzinski, N.; Njo, M.F.; et al. Cyclic programmed cell death stimulates hormone signaling and root development in Arabidopsis. Science 2016, 351, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Malamy, J.E. Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 2005, 28, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Benkova, E.; Bielach, A. Lateral root organogenesis—From cell to organ. Curr. Opin. Plant Biol. 2010, 13, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Dastidar, M.G.; Jouannet, V.; Maizel, A. Root branching: Mechanisms, robustness, and plasticity. WIREs Dev. Biol. 2012, 1, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Teeples, M.; Lanctot, A.; Nemhauser, J.L. As above, so below: Auxin’s role in lateral organ development. Dev. Biol. 2016, 419, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, J.G.; Sauer, M.; Napsucialy-Mendivil, S.; Ivanchenko, M.G.; Friml, J.; Shishkova, S.; Celenza, J.; Benková, E. Auxin acts as a local morphogenetic trigger to specify lateral root founder cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8790–8794. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Cervera, M.-T.; Delarue, M.; Beeckman, T.; Dewitte, W.; Bellini, C.; Caboche, M.; Van Onckelen, H.; Van Montagu, M.; Inzé, D. Superroot, a recessive mutation in Arabidopsis, confers auxin overproduction. Plant Cell 1995, 7, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Delarue, M.; Prinsen, E.; Va, H.; Caboche, M.; Bellini, C. Sur2 mutations of Arabidopsis thaliana define a new locus involved in the control of auxin homeostasis. Plant J. 1998, 14, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.Z.; Wei, J.; Xu, J.; Sun, M.X. Inducible knock-down of GNOM during root formation reveals tissue-specific response to auxin transport and its modulation of local auxin biosynthesis. J. Exp. Bot. 2014, 65, 1165–1179. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Sun, B.; Xu, L.; Liu, W. Wound signaling: The missing link in plant regeneration. Plant Signal. Behav. 2016, 11, e1238548. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, N. Getting to the root of regeneration: Adventitious rooting and callus formation. Plant Cell 2014, 26, 845. [Google Scholar] [CrossRef] [PubMed]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Pacurar, D.I.; Perrone, I.; Bellini, C. Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiol. Plant. 2014, 151, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha Correa, L.; Troleis, J.; Mastroberti, A.; Mariath, J.; Fett-Neto, A. Distinct modes of adventitious rooting in Arabidopsis thaliana. Plant Biol. 2012, 14, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Pop, T.I.; Pamfil, D.; Bellini, C. Auxin control in the formation of adventitious roots. Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 307–316. [Google Scholar]
- Ahkami, A.H.; Lischewski, S.; Haensch, K.T.; Porfirova, S.; Hofmann, J.; Rolletschek, H.; Melzer, M.; Franken, P.; Hause, B.; Druege, U.; et al. Molecular physiology of adventitious root formation in Petunia hybrida cuttings: Involvement of wound response and primary metabolism. New Phytol. 2009, 181, 613–625. [Google Scholar] [CrossRef] [PubMed]
- Druege, U.; Franken, P.; Lischewski, S.; Ahkami, A.H.; Zerche, S.; Hause, B.; Hajirezaei, M.R. Transcriptomic analysis reveals ethylene as stimulator and auxin as regulator of adventitious root formation in petunia cuttings. Front. Plant Sci. 2014, 5, 494. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Miao, J.; Yumoto, E.; Yokota, T.; Asahina, M.; Watahiki, M. YUCCA9-mediated auxin biosynthesis and polar auxin transport synergistically regulate regeneration of root systems following root cutting. Plant Cell Physiol. 2017, 58, 1710–1723. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Tong, J.H.; Xiao, L.T.; Ruan, Y.; Liu, J.C.; Zeng, M.H.; Huang, H.; Wang, J.W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.L.; Cheng, Z.J.; Zhang, X.S. Endogenous auxin biosynthesis and de novo root organogenesis. J. Exp. Bot. 2016, 67, 4011–4013. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.B.; Chen, L.Q.; Liu, J.C.; Zhang, X.N.; Yang, Z.N.; Liu, W.; Xu, L. TAA family contributes to auxin production during de novo regeneration of adventitious roots from Arabidopsis leaf explants. Sci. Bull. 2016, 61, 1728–1731. [Google Scholar] [CrossRef]
- Della Rovere, F.; Fattorini, L.; D’Angeli, S.; Veloccia, A.; Falasca, G.; Altamura, M.M. Auxin and cytokinin control formation of the quiescent centre in the adventitious root apex of Arabidopsis. Ann. Bot. 2013, 112, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Della Rovere, F.; Fattorini, L.; Ronzan, M.; Falasca, G.; Altamura, M.M. The quiescent center and the stem cell niche in the adventitious roots of Arabidopsis thaliana. Plant Signal. Behav. 2016, 11, e1176660. [Google Scholar] [CrossRef] [PubMed]
- Blakesley, D. Auxin metabolism and adventitious root initiation in Biology of adventitious root formation. Eds. Davis, T.D.; Haissig, B.E. Springer, Boston MA. Basic Life Sci. 1994, 62, 143. [Google Scholar]
- Zhang, W.; Swarup, R.; Bennett, M.; Schaller, G.E.; Kieber, J.J. Cytokinin induces cell division in the quiescent center of the Arabidopsis root apical meristem. Curr. Biol. 2013, 23, 1979–1989. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.J.; Estelle, M. Cytokinin and auxin intersection in root meristems. Genome Biol. 2009, 10, 210. [Google Scholar] [CrossRef]
- Marhavy, P.; Duclercq, J.; Weller, B.; Feraru, E.; Bielach, A.; Offringa, R.; Friml, J.; Schwechheimer, C.; Murphy, A.; Benkova, E. Cytokinin controls polarity of PIN1-dependent auxin transport during lateral root organogenesis. Curr. Biol. 2014, 24, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Simaskova, M.; O’Brien, J.A.; Khan, M.; Van Noorden, G.; Otvos, K.; Vieten, A.; De Clercq, I.; Van Haperen, J.M.; Cuesta, C.; Hoyerova, K.; et al. Cytokinin response factors regulate PIN-FORMED auxin transporters. Nat. Commun. 2015, 6, 8717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.Y.; Zhang, C.G.; Wu, L.; Zhang, C.G.; Chai, J.; Wang, M.; Jha, A.; Jia, P.F.; Cui, S.J.; Yang, M. Functional characterization of the CKRC1/TAA1 gene and dissection of hormonal actions in the Arabidopsis root. Plant J. 2011, 66, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Gunnerås, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin regulation of auxin synthesis in Arabidopsis involves a homeostatic feedback loop regulated via auxin and cytokinin signal transduction. Plant Cell 2010, 22, 2956–2969. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.I.; Noh, E.W.; Kim, H.J.; Park, W.J. Differential regulation of cytokinin oxidase genes and cytokinin-induced auxin biosynthesis by cellular cytokinin level in transgenic poplars. Plant Cell Rep. 2014, 33, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.; Ljung, K. Auxin and cytokinin regulate each other’s levels via a metabolic feedback loop. Plant Signal. Behav. 2011, 6, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Di, D.W.; Wu, L.; Luo, P.; Zhang, L.; Zhang, T.Z.; Sun, X.; Wei, S.D.; An, C.W.; Guo, G.Q. Analysis the role of arabidopsis CKRC6/ASA1 in auxin and cytokinin biosynthesis. J. Plant Biol. 2016, 59, 162–171. [Google Scholar] [CrossRef]
- Moubayidin, L.; Di Mambro, R.; Sozzani, R.; Pacifici, E.; Salvi, E.; Terpstra, I.; Bao, D.; Van Dijken, A.; Ioio, R.D.; Perilli, S. Spatial coordination between stem cell activity and cell differentiation in the root meristem. Dev. Cell 2013, 26, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Ivanchenko, M.G.; Muday, G.K.; Dubrovsky, J.G. Ethylene-auxin interactions regulate lateral root initiation and emergence in Arabidopsis thaliana. Plant J. 2008, 55, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, K.; Ljung, K.; Vanneste, S.; Podhorska, R.; Beeckman, T.; Friml, J.; Benkova, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanova, A.N.; Hoyt, J.M.; Hamilton, A.A.; Alonso, J.M. A link between ethylene and auxin uncovered by the characterization of two root-specific ethylene-insensitive mutants in Arabidopsis. Plant Cell 2005, 17, 2230–2242. [Google Scholar] [CrossRef] [PubMed]
- Swarup, R.; Perry, P.; Hagenbeek, D.; Van Der Straeten, D.; Beemster, G.T.S.; Sandberg, G.; Bhalerao, R.; Ljung, K.; Bennett, M.J. Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation. Plant Cell 2007, 19, 2186–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.L.; Miao, Z.Q.; Wang, Z.; Yu, L.H.; Cai, X.T.; Xiang, C.B. Arabidopsis ERF1 mediates crosstalk between ethylene and auxin biosynthesis during primary root elongation by regulating ASA1 expression. PLoS Genet. 2016, 12, e1005760. [Google Scholar]
- He, W.R.; Brumos, J.; Li, H.J.; Ji, Y.S.; Ke, M.; Gong, X.Q.; Zeng, Q.L.; Li, W.Y.; Zhang, X.Y.; An, F.Y.; et al. A small-molecule screen identifies L-Kynurenine as a competitive inhibitor of TAA1/TAR activity in ethylene-directed auxin biosynthesis and root growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Zhang, Z.; Wang, J.; Chen, X.; Wei, P.; Huang, R. The activation of OsEIL1 on YUC8 transcription and auxin biosynthesis is required for ethylene-inhibited root elongation in rice early seedling development. PLoS Genet. 2017, 13, e1006955. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.Y.; Chao, Y.Y.; Kao, C.H. Methyl jasmonate-induced lateral root formation in rice: The role of heme oxygenase and calcium. J. Plant Physiol. 2013, 170, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Raya-Gonzalez, J.; Pelagio-Flores, R.; Lopez-Bucio, J. The jasmonate receptor COI1 plays a role in jasmonate-induced lateral root formation and lateral root positioning in Arabidopsis thaliana. J. Plant Physiol. 2012, 169, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Q.; Xu, Y.X.; Ye, S.Q.; Jiang, H.L.; Chen, Q.; Liu, F.; Zhou, W.K.; Chen, R.; Li, X.G.; Tietz, O.; et al. Arabidopsis ASA1 is important for jasmonate-mediated regulation of auxin biosynthesis and transport during lateral root formation. Plant Cell 2009, 21, 1495–1511. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.T.; Xu, P.; Zhao, P.X.; Liu, R.; Yu, L.H.; Xiang, C.B. Arabidopsis ERF109 mediates cross-talk between jasmonic acid and auxin biosynthesis during lateral root formation. Nat. Commun. 2014, 5, 5833. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Galván-Ampudia, C.S.; Demarsy, E.; Langowski, L.; Kleine-Vehn, J.; Fan, Y.; Morita, M.T.; Tasaka, M.; Fankhauser, C.; Offringa, R. Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis. Nat. Cell Biol. 2011, 13, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Sassi, M.; Lu, Y.; Zhang, Y.; Wang, J.; Dhonukshe, P.; Blilou, I.; Dai, M.; Li, J.; Gong, X.; Jaillais, Y. COP1 mediates the coordination of root and shoot growth by light through modulation of PIN1-and PIN2-dependent auxin transport in Arabidopsis. Development 2012, 139, 3402–3412. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.-L.; Okamoto, H.; Wang, M.; Ang, L.-H.; Matsui, M.; Goodman, H.; Deng, X.W. FIN219, an auxin-regulated gene, defines a link between phytochrome A and the downstream regulator COP1 in light control of Arabidopsis development. Genes Dev. 2000, 14, 1958–1970. [Google Scholar] [PubMed]
- Nakazawa, M.; Yabe, N.; Ichikawa, T.; Yamamoto, Y.Y.; Yoshizumi, T.; Hasunuma, K.; Matsui, M. DFL1, an auxin-responsive GH3 gene homologue, negatively regulates shoot cell elongation and lateral root formation, and positively regulates the light response of hypocotyl length. Plant J. 2001, 25, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Sorin, C.; Bussell, J.D.; Camus, I.; Ljung, K.; Kowalczyk, M.; Geiss, G.; McKhann, H.; Garcion, C.; Vaucheret, H.; Sandberg, G.; et al. Auxin and light control of adventitious rooting in Arabidopsis require ARGONAUTE1. Plant Cell 2005, 17, 1343–1359. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.-I.; Mochizuki, N.; Nagatani, A. Expression of the AtGH3a gene, an Arabidopsis homologue of the soybean GH3 gene, is regulated by phytochrome B. Plant Cell Physiol. 2002, 43, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Hoecker, U.; Toledo-Ortiz, G.; Bender, J.; Quail, P.H. The photomorphogenesis-related mutant red1 is defective in CYP83B1, a red light-induced gene encoding a cytochrome P450 required for normal auxin homeostasis. Planta 2004, 219, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Yokawa, K.; Nakano, S.; Yoshida, Y.; Fabrissin, I.; Okamoto, T.; Baluska, F.; Koshiba, T. Root cap-dependent gravitropic U-turn of maize root requires light-induced auxin biosynthesis via the YUC pathway in the root apex. J. Exp. Bot. 2016, 67, 4581–4591. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Guo, Y.; Novák, O.; Chen, W.; Ljung, K.; Noel, J.P.; Chory, J. Local auxin metabolism regulates environment-induced hypocotyl elongation. Nat. Plants 2016, 2, 16025. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ljung, K.; Breton, G.; Schmitz, R.J.; Pruneda-Paz, J.; Cowing-Zitron, C.; Cole, B.J.; Ivans, L.J.; Pedmale, U.V.; Jung, H.S.; et al. Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 2012, 26, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Sairanen, I.; Novák, O.; Pěnčík, A.; Ikeda, Y.; Jones, B.; Sandberg, G.; Ljung, K. Soluble carbohydrates regulate auxin biosynthesis via PIF proteins in Arabidopsis. Plant Cell 2012, 24, 4907–4916. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.S.; Singh, M.; Aggrawal, P.; Laxmi, A. Glucose and auxin signaling interaction in controlling Arabidopsis thaliana seedlings root growth and development. PLoS ONE 2009, 4, e4502. [Google Scholar] [CrossRef] [PubMed]
- Lilley, J.L.S.; Gee, C.W.; Sairanen, I.; Ljung, K.; Nemhauser, J.L. An endogenous carbon-sensing pathway triggers increased auxin flux and hypocotyl elongation. Plant Physiol. 2012, 160, 2261–2270. [Google Scholar] [CrossRef] [PubMed]
- Giehl, R.F.; von Wiren, N. Root nutrient foraging. Plant Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wirén, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Ristova, D.; Busch, W. Natural variation of root traits: From development to nutrient uptake. Plant Physiol. 2014, 166, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Remans, T.; Nacry, P.; Pervent, M.; Girin, T.; Tillard, P.; Lepetit, M.; Gojon, A. A central role for the nitrate transporter NRT2.1 in the integrated morphological and physiological responses of the root system to nitrogen limitation in Arabidopsis. Plant Physiol. 2006, 140, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Forde, B.G. Regulation of Arabidopsis root development by nitrate availability. J. Exp. Bot. 2000, 51, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Jennings, A.; Barlow, P.W.; Forde, B.G. Dual pathways for regulation of root branching by nitrate. Proc. Natl. Acad. Sci. USA 1999, 96, 6529–6534. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Chen, F.; Liu, J.; Zhang, F.; Mi, G. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots. J. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Walch-Liu, P.; Ivanov, I.I.; Filleur, S.; Gan, Y.; Remans, T.; Forde, B.G. Nitrogen regulation of root branching. Ann. Bot. 2005, 97, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.Y.; Li, J.J.; Qu, B.Y.; He, X.; Zhao, X.Q.; Li, B.; Fu, X.D.; Tong, Y.P. Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in Arabidopsis. Plant J. 2014, 78, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Shao, A.; Ma, W.; Zhao, X.; Hu, M.; He, X.; Teng, W.; Li, H.; Tong, Y. The auxin biosynthetic TRYPTOPHAN AMINOTRANSFERASE RELATED TaTAR2.1-3A increases grain yield of wheat. Plant Physiol. 2017, 174, 2274–2288. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.H.; Miao, Z.Q.; Qi, G.F.; Wu, J.; Cai, X.T.; Mao, J.L.; Xiang, C.B. MADS-box transcription factor AGL21 regulates lateral root development and responds to multiple external and physiological signals. Mol. Plant 2014, 7, 1653–1669. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Lee, J.; Gong, Q.; Ma, S.; Jin, J.B.; Yoo, C.Y.; Miura, T.; Sato, A.; Bohnert, H.J.; Hasegawa, P.M. SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation. Plant Physiol. 2011, 155, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Qu, B.; He, X.; Wang, J.; Zhao, Y.; Teng, W.; Shao, A.; Zhao, X.; Ma, W.; Wang, J.; Li, B.; et al. A wheat CCAAT box-binding transcription factor increases the grain yield of wheat with less fertilizer input. Plant Physiol. 2015, 167, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Kutz, A.; Muller, A.; Hennig, P.; Kaiser, W.M.; Piotrowski, M.; Weiler, E.W. A role for nitrilase 3 in the regulation of root morphology in sulphur-starving Arabidopsis thaliana. Plant J. 2002, 30, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Blancaflor, E.B.; Masson, P.H. Plant gravitropism. Unraveling the ups and downs of a complex process. Plant Physiol. 2003, 133, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Band, L.R.; Wells, D.M.; Larrieu, A.; Sun, J.Y.; Middleton, A.M.; French, A.P.; Brunoud, G.; Sato, E.M.; Wilson, M.H.; Peret, B.; et al. Root gravitropism is regulated by a transient lateral auxin gradient controlled by a tipping-point mechanism. Proc. Natl. Acad. Sci. USA 2012, 109, 4668–4673. [Google Scholar] [CrossRef] [PubMed]
- Geisler, M.; Wang, B.; Zhu, J. Auxin transport during root gravitropism: Transporters and techniques. Plant Biol. 2014, 16 (Suppl. S1), 50–57. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Kim, W.Y.; Kang, S.B.; Kim, J.I.; Baek, D.; Jung, I.J.; Kim, M.R.; Li, N.; Kim, H.J.; Nakajima, M.; et al. A novel thiol-reductase activity of Arabidopsis YUC6 confers drought tolerance independently of auxin biosynthesis. Nat. Commun. 2015, 6, 8041. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Wang, Z.; Ji, C.Y.; Jeong, J.C.; Lee, H.S.; Li, H.; Xu, B.; Deng, X.; Kwak, S.S. Transgenic poplar expressing Arabidopsis YUCCA6 exhibits auxin-overproduction phenotypes and increased tolerance to abiotic stress. Plant Physiol. Biochem. 2015, 94, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Baek, D.; Park, H.C.; Chun, H.J.; Oh, D.H.; Lee, M.K.; Cha, J.Y.; Kim, W.Y.; Kim, M.C.; Chung, W.S.; et al. Overexpression of Arabidopsis YUCCA6 in potato results in high auxin developmental phenotypes and enhanced resistance to water deficit. Mol. Plant 2013, 6, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Jung, J.-H.; Han, D.Y.; Seo, P.J.; Park, W.J.; Park, C.M. Activation of a flavin monooxygenase gene YUCCA7 enhances drought resistance in Arabidopsis. Planta 2012, 235, 923–938. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Wang, H.; Kasahara, H.; Liu, J.; MacPherson, C.; Machida, Y.; Kamiya, Y.; Hannah, M.A.; Chua, N.H. IAA-Ala Resistant 3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 2012, 24, 3590–3602. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.E.; Dinneny, J.R. The divining root: Moisture-driven responses of roots at the micro- and macro-scale. J. Exp. Bot. 2015, 66, 2145–2154. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Aggarwal, P.; Robbins, N.E.; Sturrock, C.J.; Thompson, M.C.; Tan, H.Q.; Tham, C.; Duan, L.N.; Rodriguez, P.L.; Vernoux, T.; et al. Plant roots use a patterning mechanism to position lateral root branches toward available water. Proc. Natl. Acad. Sci. USA 2014, 111, 9319–9324. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Tripathi, D.K.; Singh, S.; Sharma, S.; Dubey, N.K.; Chauhan, D.K.; Vaculik, M. Toxicity of aluminium on various levels of plant cells and organism: A review. Environ. Exp. Bot. 2017, 137, 177–193. [Google Scholar] [CrossRef]
- Sun, L.; Tian, J.; Zhang, H.; Liao, H. Phytohormone regulation of root growth triggered by P deficiency or Al toxicity. J. Exp. Bot. 2016, 67, 3655–3664. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Tian, Q.Y.; Chen, J.; Zhang, W.H. Aluminium-induced inhibition of root elongation in Arabidopsis is mediated by ethylene and auxin. J. Exp. Bot. 2010, 61, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Ren, X.Y.; Huang, B.R.; Wang, G.; Zhou, P.; An, Y. Aluminium-induced reduction of plant growth in alfalfa (Medicago sativa) is mediated by interrupting auxin transport and accumulation in roots. Sci. Rep. 2016, 6, 30079. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.B.; Geng, X.Y.; He, C.M.; Zhang, F.; Wang, R.; Horst, W.J.; Ding, Z.J. TAA1-regulated local auxin biosynthesis in the root apex transition zone mediates the Aluminum-induced inhibition of root growth in Arabidopsis. Plant Cell 2014, 26, 2889–2904. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.B.; Liu, G.; Liu, J.; Zhang, B.; Meng, W.; Müller, B.; Hayashi, K.I.; Zhang, X.; Zhao, Z.; De Smet, I.; et al. Synergistic action of auxin and cytokinin mediates aluminum-induced root growth inhibition in Arabidopsis. EMBO Rep. 2017, 18, 1213–1230. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.C.; Gao, S.; Tian, H.Y.; Wu, W.W.; Robert, H.S.; Ding, Z.J. Local transcriptional control of YUCCA regulates auxin promoted root growth inhibition in response to Aluminium stress in Arabidopsis. PLoS Genet. 2016, 12, e1006360. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.F.; Lei, G.J.; Wang, Z.W.; Shi, Y.Z.; Braam, J.; Li, G.X.; Zheng, S.J. Coordination between apoplastic and symplastic detoxification confers plant aluminum resistance. Plant Physiol. 2013, 162, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Hou, N.; Schlicht, M.; Wan, Y.; Mancuso, S.; Baluska, F. Aluminium toxicity targets PIN2 in Arabidopsis root apices: Effects on PIN2 endocytosis, vesicular recycling, and polar auxin transport. Chin. Sci. Bull. 2008, 53, 2480. [Google Scholar] [CrossRef]
- Wu, D.; Shen, H.; Yokawa, K.; Baluška, F. Alleviation of aluminium-induced cell rigidity by overexpression of OsPIN2 in rice roots. J. Exp. Bot. 2014, 65, 5305–5315. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, W.; Zhou, H.; Wang, R.; Zhang, P.; Wang, H.; Pan, X.; Xu, J. Manganese toxicity inhibited root growth by disrupting auxin biosynthesis and transport in Arabidopsis. Front. Plant Sci. 2017, 8, 272. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, Y.; Pan, Z.; Xie, S.; Peng, S.A. Boron deficiency alters root growth and development and interacts with auxin metabolism by influencing the expression of auxin synthesis and transport genes. Biotechnol. Biotechnol. Equip. 2016, 30, 661–668. [Google Scholar] [CrossRef]
- Sukumar, P.; Legue, V.; Vayssieres, A.; Martin, F.; Tuskan, G.A.; Kalluri, U.C. Involvement of auxin pathways in modulating root architecture during beneficial plant–microorganism interactions. Plant Cell Environ. 2013, 36, 909–919. [Google Scholar] [CrossRef] [PubMed]
- Felten, J.; Legue, V.; Ditengou, F.A. Lateral root stimulation in the early interaction between Arabidopsis thaliana and the ectomycorrhizal fungus Laccaria bicolor. Plant Signal. Behav. 2010, 5, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.; Zhang, N.; Li, Z.; Zhang, G.; Xu, Y.; Shen, Q.; Zhang, R. Plant-microbe communication enhances auxin biosynthesis by a root-associated bacterium, Bacillus amyloliquefaciens SQR9. Mol. Plant-Microbe Interact. 2016, 29, 324–330. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, P.J.; Schmelz, E.A.; Moussatche, P.; Lund, S.T.; Jones, J.B.; Klee, H.J. usceptible to intolerance—A range of hormonal actions in a susceptible Arabidopsis pathogen response. Plant J. 2003, 33, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Rampey, R.A.; LeClere, S.; Kowalczyk, M.; Ljung, K.; Sandberg, G.; Bartel, B. A family of auxin-conjugate hydrolases that contributes to free indole-3-acetic acid levels during Arabidopsis germination. Plant Physiol. 2004, 135, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Normanly, J. Approaching cellular and molecular resolution of auxin biosynthesis and metabolism. Cold Spring Harb. Perspect. Biol. 2010, 2, a001594. [Google Scholar] [CrossRef] [PubMed]
- LeClere, S.; Tellez, R.; Rampey, R.A.; Matsuda, S.P.; Bartel, B. Characterization of a family of IAA-amino acid conjugate hydrolases from Arabidopsis. J. Biol. Chem. 2002, 277, 20446–20452. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Piotrowska, A. Conjugates of auxin and cytokinin. Phytochemistry 2009, 70, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Riov, J.; Bangerth, F. Metabolism of auxin in tomato fruit tissue: Formation of high molecular weight conjugates of oxindole-3-acetic acid via the oxidation of indole-3-acetylaspartic acid. Plant Physiol. 1992, 100, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Östin, A.; Kowalyczk, M.; Bhalerao, R.P.; Sandberg, G. Metabolism of indole-3-acetic acid in Arabidopsis. Plant Physiol. 1998, 118, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, M.; Sandberg, G. Quantitative analysis of indole-3-acetic acid metabolites in Arabidopsis. Plant Physiol. 2001, 127, 1845–1853. [Google Scholar] [CrossRef] [PubMed]
- Eyer, L.; Vain, T.; Pařízková, B.; Oklestkova, J.; Barbez, E.; Kozubíková, H.; Pospíšil, T.; Wierzbicka, R.; Kleine-Vehn, J.; Fránek, M.; et al. 2,4-D and IAA amino acid conjugates show distinct metabolism in Arabidopsis. PLoS ONE 2016, 11, e0159269. [Google Scholar] [CrossRef] [PubMed]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, C.; Dennis, E.G.; Booker, G.W.; Polyak, S.W.; Boss, P.K.; Davies, C. A novel tool for studying auxin-metabolism: The inhibition of grapevine indole-3-acetic acid-amido synthetases by a reaction intermediate analogue. PLoS ONE 2012, 7, e37632. [Google Scholar] [CrossRef] [PubMed]
- Bartel, B.; Fink, G. ILR1, an amidohydrolase that releases active indole-3-acetic acid from conjugates. Science 1995, 268, 1745–1748. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.T.; Goetz, D.H.; Lasswell, J.; Anderson, M.N.; Bartel, B. IAR3 encodes an auxin conjugate hydrolase from Arabidopsis. Plant Cell 1999, 11, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Magidin, M.; Pittman, J.K.; Hirschi, K.D.; Bartel, B. ILR2, a novel gene regulating IAA conjugate sensitivity and metal transport in Arabidopsis thaliana. Plant J. 2003, 35, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Gu, H.; Zhao, Y.; Ma, Z.; Shi, G.; Yang, Y.; Pichersky, E.; Chen, H.; Liu, M.; Chen, Z.; et al. An indole-3-acetic acid carboxyl methyltransferase regulates Arabidopsis leaf development. Plant Cell 2005, 17, 2693–2704. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, R.; Ma, C.-J.; Vlot, A.C.; Klessig, D.F.; Pichersky, E. Inactive methyl indole-3-acetic acid ester can be hydrolyzed and activated by several esterases belonging to the AtMES esterase family of Arabidopsis. Plant Physiol. 2008, 147, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Spiess, G.M.; Hausman, A.; Yu, P.; Cohen, J.D.; Rampey, R.A.; Zolman, B.K. Auxin input pathway disruptions are mitigated by changes in auxin biosynthetic gene expression in Arabidopsis. Plant Physiol. 2014, 165, 1092–1104. [Google Scholar] [CrossRef] [PubMed]
- Bertoni, G. Indolebutyric acid–derived auxin and plant development. Plant Cell 2011, 23, 845. [Google Scholar] [CrossRef]
- Frick, E.M.; Strader, L.C. Roles for IBA-derived auxin in plant development. J. Exp. Bot. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Peer, W.A. Auxin homeostasis: The DAO of catabolism. J. Exp. Bot. 2017, 68, 3145–3154. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.M. Destruction of Auxin. Ann. Rev. Plant Physiol. 1958, 9, 81–118. [Google Scholar] [CrossRef]
- Pěnčík, A.; Simonovik, B.; Petersson, S.V.; Henyková, E.; Simon, S.; Greenham, K.; Zhang, Y.; Kowalczyk, M.; Estelle, M.; Zažímalová, E.; et al. Regulation of auxin homeostasis and gradients in Arabidopsis roots through the formation of the indole-3-acetic acid catabolite 2-oxindole-3-acetic acid. Plant Cell 2013, 25, 3858–3870. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.N.; Alonso, J.M. Auxin catabolism unplugged: Role of IAA oxidation in auxin homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 10742–10744. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lin, J.E.; Harris, C.; Campos Mastrotti Pereira, F.; Wu, F.; Blakeslee, J.J.; Peer, W.A. DAO1 catalyzes temporal and tissue-specific oxidative inactivation of auxin in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 11010–11015. [Google Scholar] [CrossRef] [PubMed]
- Porco, S.; Pěnčík, A.; Rashed, A.; Voß, U.; Casanova-Sáez, R.; Bishopp, A.; Golebiowska, A.; Bhosale, R.; Swarup, R.; Swarup, K.; et al. Dioxygenase-encoding AtDAO1 gene controls IAA oxidation and homeostasis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 11016–11021. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, M.; Ten Tusscher, K.H. Periodic lateral root priming: What makes it tick? Plant Cell 2017, 29, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, Y.; Liu, X.; Zhang, X.; Liu, S.; Yu, X.; Ren, Y.; Zheng, X.; Zhou, K.; Jiang, L.; et al. A Role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell 2013, 27, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.D.; Gallagher, T.F. Characterization of auxin-induced ARRO-1 expression in the primary root of Malus domestica. J. Exp. Bot. 2000, 51, 1765–1766. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Hayashi, K.-I.; Natsume, M.; Kamiya, Y.; Sakakibara, H.; Kawaide, H.; Kasahara, H. UGT74D1 catalyzes the glucosylation of 2-oxindole-3-acetic acid in the auxin metabolic pathway in Arabidopsis. Plant Cell Physiol. 2014, 55, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Saito, K. Functional genomics for plant natural product biosynthesis. Nat. Prod. Rep. 2009, 26, 1466–1487. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Ma, X.M.; Han, P.; Wang, B.; Sun, Y.G.; Zhang, G.Z.; Li, Y.J.; Hou, B.K. UGT74D1 Is a novel auxin glycosyltransferase from Arabidopsis thaliana. PLoS ONE 2013, 8, e61705. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Van Aken, O.; Morreel, K.; Vandenbroucke, K.; van de Cotte, B.; De Clercq, I.; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szerszen, J.; Szczyglowski, K.; Bandurski, R. iaglu, a gene from Zea mays involved in conjugation of growth hormone indole-3-acetic acid. Science 1994, 265, 1699–1701. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Baldauf, S.; Lim, E.K.; Bowles, D.J. Phylogenetic analysis of the UDP-glycosyltransferase multigene family of Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 4338–4343. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Li, Y.; Lim, E.K.; Bowles, D.J. Higher plant glycosyltransferases. Genome Biol. 2001, 2, 1–6. [Google Scholar] [CrossRef]
- Jackson, R.G.; Lim, E.-K.; Li, Y.; Kowalczyk, M.; Sandberg, G.; Hoggett, J.; Ashford, D.A.; Bowles, D.J. Identification and biochemical characterization of an Arabidopsis indole-3-acetic acid glucosyltransferase. J. Biol. Chem. 2001, 276, 4350–4356. [Google Scholar] [CrossRef] [PubMed]
- Mravec, J.; Skupa, P.; Bailly, A.; Hoyerova, K.; Krecek, P.; Bielach, A.; Petrasek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.-D.; et al. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Auxin biosynthetic pathways in Arabidopsis thaliana. The IPyA pathway is shown in blue, the TAM pathway in green, the IAOx pathway in pink, the IAM pathway in yellow, the Trp-independent pathway in gray and the proposed metabolic pathways in purple. Enzymatic reactions without a known catalyzing enzyme are shown by dashed arrows. AO: aldehyde oxidases; IAA: 3-indole acetic acid; IAD: indole-3-acetaldehyde; IAM: indole-3-acetamide; IAN: indole-3-acetonitrile; IAOx: indole-3-acetaldoxime; IBA: indole-3-butyric acid; IG: indole glucosinolates; IGS: Indole-3-glycerol phosphate synthase; IPyA: indole-3-pyruvic acid; TAA1: TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1; TAR1-4: TRYPTOPHAN AMINOTRANSFERASE RELATED 1-4; TAM: tryptamine; TRP: tryptophan; TSA1: Trp synthase a; TSB: Trp synthase b; YUC1-11: yucca genes encoding flavin-containing monooxygenases.
Figure 1.
Auxin biosynthetic pathways in Arabidopsis thaliana. The IPyA pathway is shown in blue, the TAM pathway in green, the IAOx pathway in pink, the IAM pathway in yellow, the Trp-independent pathway in gray and the proposed metabolic pathways in purple. Enzymatic reactions without a known catalyzing enzyme are shown by dashed arrows. AO: aldehyde oxidases; IAA: 3-indole acetic acid; IAD: indole-3-acetaldehyde; IAM: indole-3-acetamide; IAN: indole-3-acetonitrile; IAOx: indole-3-acetaldoxime; IBA: indole-3-butyric acid; IG: indole glucosinolates; IGS: Indole-3-glycerol phosphate synthase; IPyA: indole-3-pyruvic acid; TAA1: TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS 1; TAR1-4: TRYPTOPHAN AMINOTRANSFERASE RELATED 1-4; TAM: tryptamine; TRP: tryptophan; TSA1: Trp synthase a; TSB: Trp synthase b; YUC1-11: yucca genes encoding flavin-containing monooxygenases.
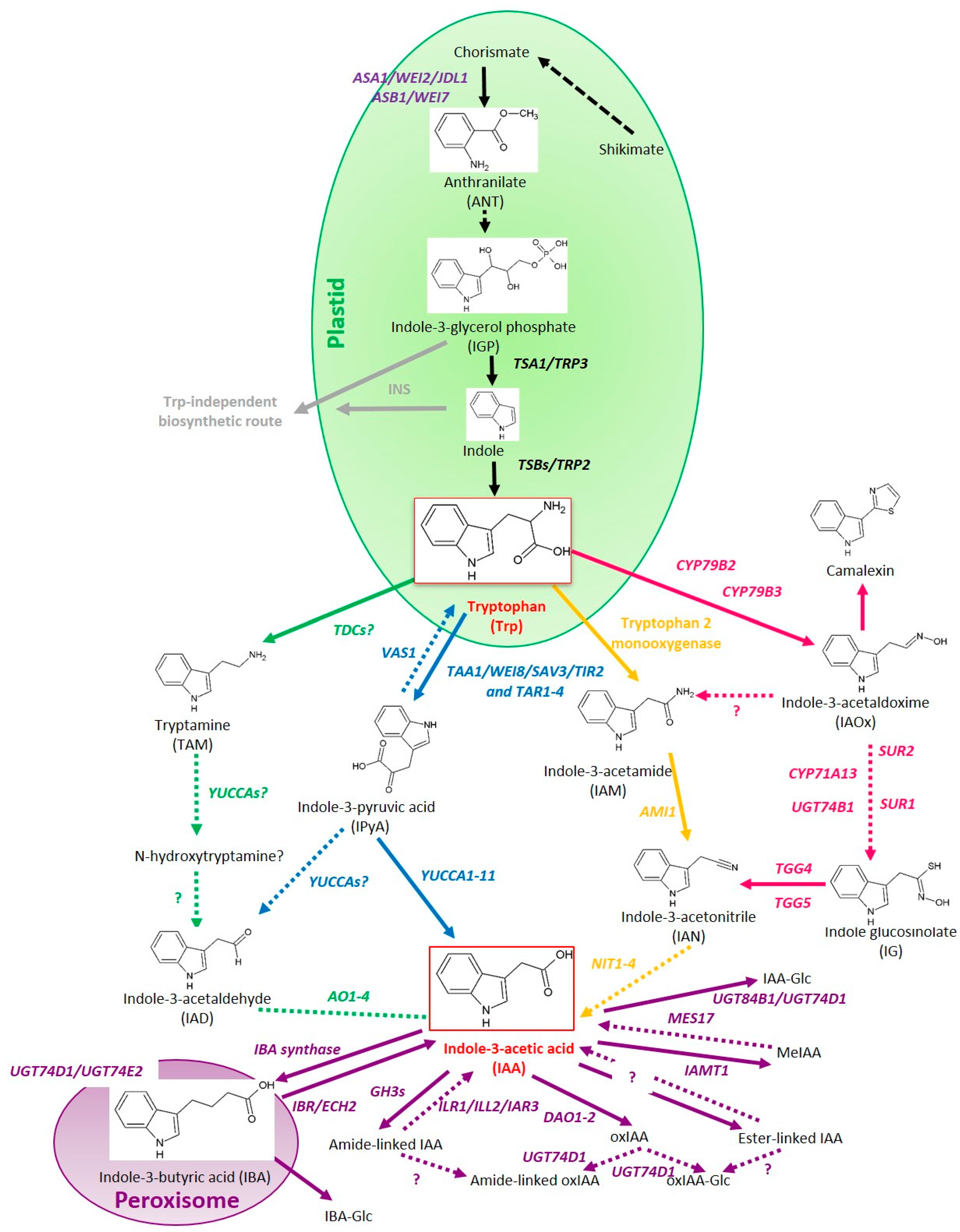

{kind=link}
{kind=link}
Table 1.
Summary auxin biosynthetic genes in root development.
| Biosynthetic Pathway | Biosynthetic Pathway | Biosynthetic Pathway | Biosynthetic Pathway |
|---|---|---|---|
| iaaM | Pseudomonas/Agrobacterium tryptophan-2-monooxygenase | Auxin biosynthesis in transgenic overexpression lines | Trp → IAM |
| iaaH | Pseudomonas/Agrobacterium IAM hydrolase | Auxin biosynthesis in transgenic overexpression lines | IAM → IAA |
| IPDC | IPA decarboxylase | Bacterial pathway | IPyA synthesis |
| ASA1/WEI2/JDL1/CKRC6/ASA2 ASB1/WEI7 | Subunits of anthranilate synthase | Root tip expression, inhibitor of sur1 Hormone interactions (cytokinin/ethylene/jasmonic acid) | Chorismate → anthranilate |
| IGS | Indole-3-glycerol phosphate synthase | Catalyzes formation of key intermediate of Trp-dependent and Trp-independent pathways | Anthranilate → indole-3-glycerol phosphate |
| TSA1/TSB1-2 | Trp synthase a and Trp synthase b | Trp synthesis | Indole-3-glycerol phosphate → indole → Trp |
| INS | TSA homolog | Early expression in embryo | |
| PDX1 | vitamin B6 biosynthesis mutant | Trp → IAA | |
| TAA1/TAR1-4 | Arabidopsis tryptophan aminotransferases | Stem cell niche specification De novo root organogenesis Control root architecture nitrogen dependently | Trp → IPyA |
| TAAI/WEI8/SAV3/TIR2 | Arabidopsis | Trp → IPyA | |
| TaTAR2 | wheat | Lateral root formation Nitrate and phosphate starvation | IPyA |
| FISHBONE | Rice Orthologue TAA1 | ||
| OsYUC1 OsCOW1 | Rice Flavin-containing monooxygenases | Post embryonic root development Crown and adventitious roots | IPyA |
| YUC1-11 | Arabidopsis Flavin-containing monooxygenases | Root embryogenesis RAM maintenance Drought resistance De novo root organogenesis Root growth in Al3+ stress | IPyA → IAA TAM → N-hydroxytryptamine? |
| YUC (ZM2G141383) | Maize Flavin-containing monooxygenases | Root cap-mediated gravitropic U-turn in response to light | IPyA → IAA |
| MYROSINASE AtTGG4/AtTGG5 | Arabidopsis | Local auxin distribution in the root | IG → IAN |
| Nitrilases NIT1-4 | Arabidopsis | Root development and growth | IAN → IAA |
| CYP79B2/CYP79B3 | Arabidopsis Cytochrome P450 | Auxin gradients in the root | Trp → IAOx |
| IBR1,3,10/ECH2 | INDOLE- 3-BUTYRIC ACID RESPONSE (IBR) ENOYL–COA HYDRATASE2 | IBA to IAA conversion Priming LR founder cells | IBA → IAA |
| AO | Aldehyde oxidase | IPyA to IAD pathway? | IDA → IAA |
| AMI1 | IAM HYDROLASE orthologue of iaaH | Required for BY2 growth on IAM medium | IAM → IAA PAM → PAA |
| SUR1/SUR2 | Glucosinolate biosynthesis genes SUPERROOT1/2 | Overproduction LR and AR | IAOx → IG |
| UGT74B1 | UDP-glycosyltransferase | High auxin levels in mutant Mutation in IG synthesis | |
| CYP71A13 | IAOx → IAN | ||
| ILR1/ILL2/IAR3 | IAA-leucine resistant1 IAA-leucine resistant-like 2 IAA-Ala Resistant3 hydrolase | Conjugated IAA → IAA | |
| GH3 | GRETCHEN HAGEN 3 (GH3) protein family | IAA-conjugation to amino acids | IAA → amide conjugated IAA |
| DAO | DIOXYGENASE FOR AUXIN OXIDATION 1 | IAA → oxIAA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Olatunji, D.; Geelen, D.; Verstraeten, I. Control of Endogenous Auxin Levels in Plant Root Development. Int. J. Mol. Sci. 2017, 18, 2587. https://doi.org/10.3390/ijms18122587
AMA Style
Olatunji D, Geelen D, Verstraeten I. Control of Endogenous Auxin Levels in Plant Root Development. International Journal of Molecular Sciences. 2017; 18(12):2587. https://doi.org/10.3390/ijms18122587
Chicago/Turabian StyleOlatunji, Damilola, Danny Geelen, and Inge Verstraeten. 2017. "Control of Endogenous Auxin Levels in Plant Root Development" International Journal of Molecular Sciences 18, no. 12: 2587. https://doi.org/10.3390/ijms18122587
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.