
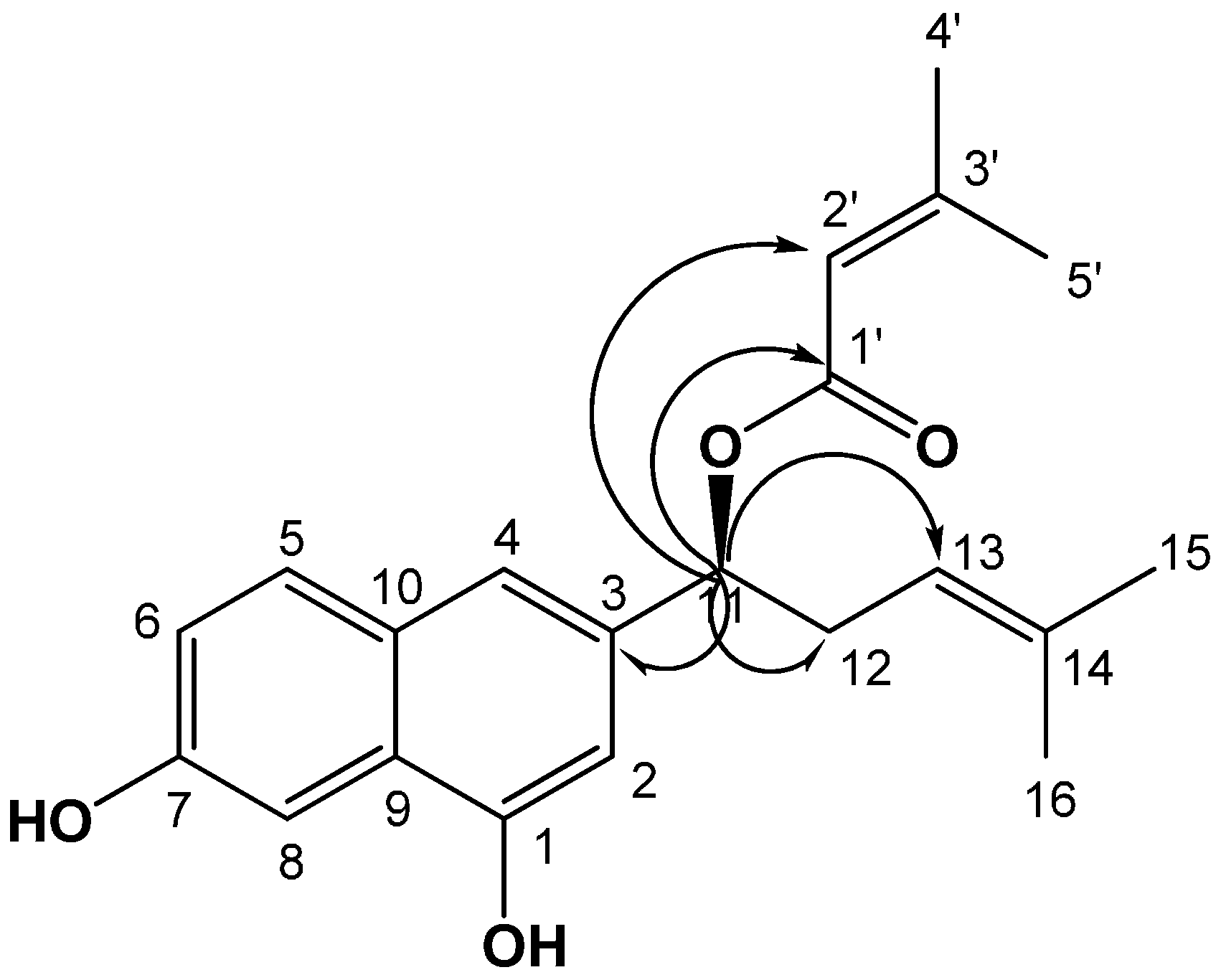
Potential of Pseudoshikonin I Isolated from Lithospermi Radix as Inhibitors of MMPs in IL-1β-Induced SW1353 Cells

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
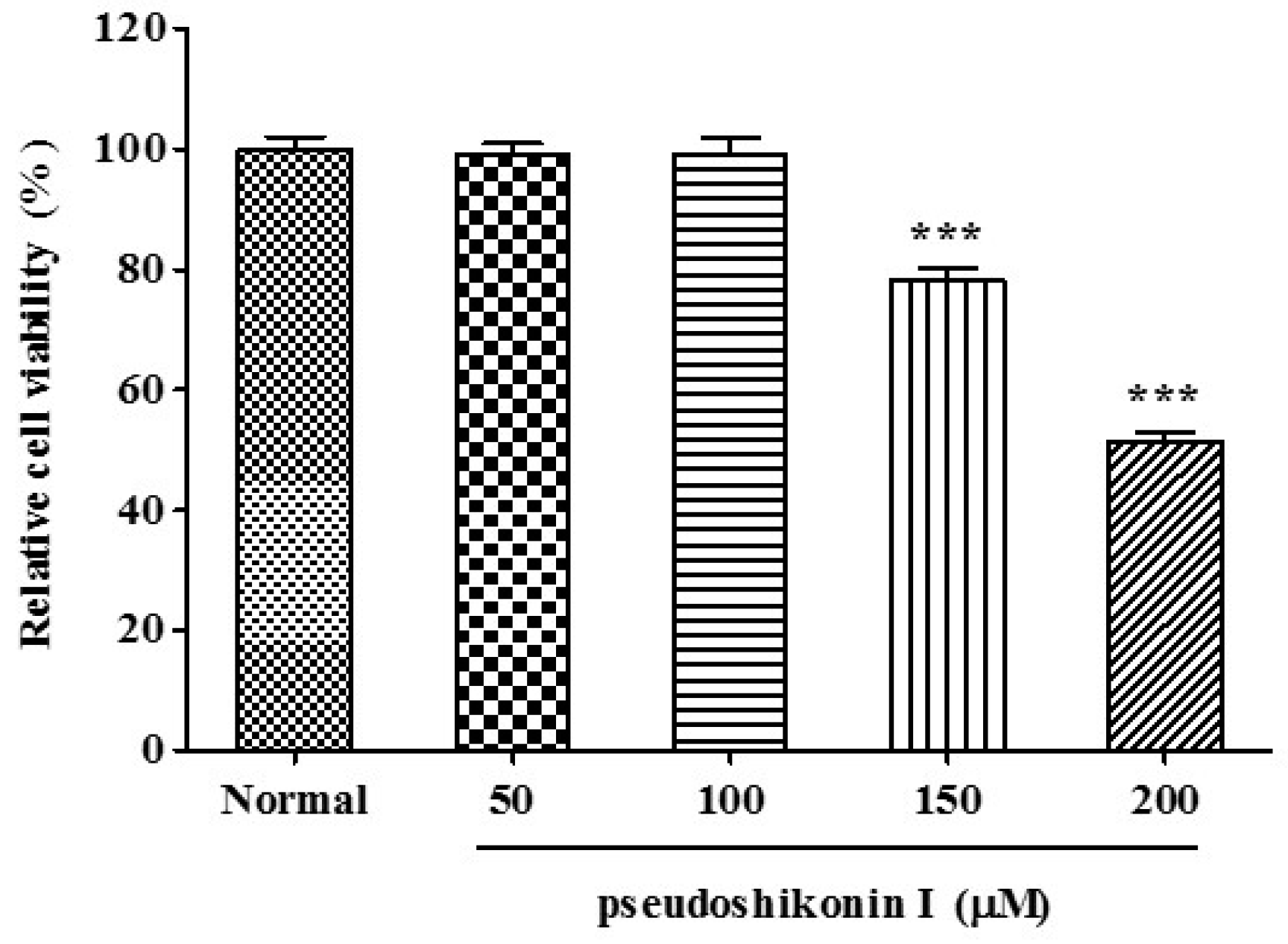
2.1. Cell Viability of Pseudoshikonin I on SW1353 Cells
2.2. Effect of Pseudoshikonin I on MMPs Activity in IL-1β-Induced SW1353 Cells
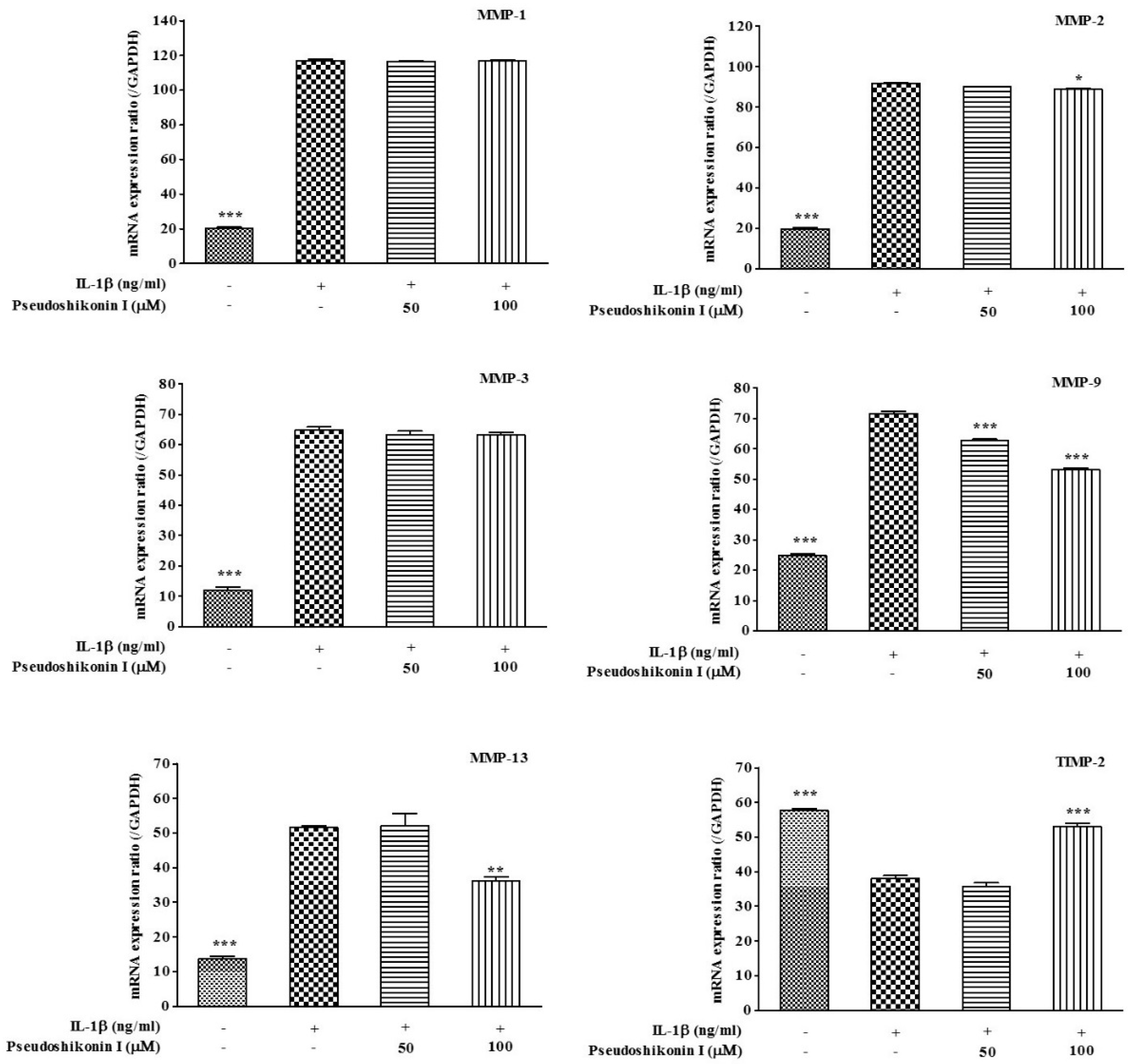
2.3. Effect of Pseudoshikonin I on the mRNA Expression of MMPs in IL-1β-Induced SW1353 Cells
3. Experiment
3.1. General
3.2. Plant Materials
3.3. Extraction and Isolation
3.4. Cell Culture
3.5. Determination of Cell Viability
3.6. Measurement of MMPs Production in IL-1β-Induced SW1353 Cells
3.7. mRNA Isolation and RT-PCR in IL-1β-Induced SW1353 Cells
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cho, M.H.; Paik, Y.S.; Hahn, T.R. Physical stability of shikonin derivatives from the roots of Lithospermm erythrorhizon cultivated in Korea. J. Agric. Food Chem. 1999, 47, 4117–4120. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.H.; Jin, Y.H.; Choe, C.Y.; Lee, M.K.; Lee, S. Ecological characteristics of Lithospermum erythrorhizon population in habitats. Korean J. Physiol. Pharmacol. 2009, 40, 289–297. [Google Scholar]
- Rajasekar, S.; Park, D.J.; Park, C.; Park, S.; Park, Y.H.; Kim, S.T.; Choi, Y.H.; Choi, Y.W. In vitro and in vivo anticancer effects of Lithospermum erythrorhizon extract on B16F10 murinemelanoma. J. Ethnopharmcol. 2012, 144, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Andújar, I.; Recio, M.C.; Giner, R.M.; Ríos, J.L. Traditional Chinese medicine remedy to jury: The pharmacological basis for the use of shikonin as an anticancer therapy. Curr. Med. Chem. 2013, 20, 2892–2898. [Google Scholar]
- Pan, Y.M.; Liang, Y.; Wang, H.S.; Liang, M. Antioxidant activities of several Chinese medicine herbs. Food Chem. 2004, 88, 347–350. [Google Scholar]
- Weng, X.C.; Xiang, G.Q.; Jiang, A.L.; Liu, Y.P.; Wu, L.L.; Dong, X.W.; Duan, S. Antioxidant properties of components extracted from puccoon (Lithospermum erythrorhizon Sieb. et Zucc.). Food Chem. 2000, 69, 143–146. [Google Scholar] [CrossRef]
- Han, J.; Weng, X.; Bi, K. Antioxidants from a Chinese medicinal herb-Lithospermum erythrorhizon. Food Chem. 2008, 106, 2–10. [Google Scholar] [CrossRef]
- Kim, G.S.; Park, C.G.; Lee, K.H.; Choi, J.H.; Lee, S.E.; Noh, H.J.; Lee, J.H.; Kim, S.Y. Investigatin of shikonin pigments and antioxidant activity of the roots from Lithospermum erythrorhizon according to the different growth stages and areas of cultivation. Korean J. Med. Crop Sci. 2011, 19, 435–440. [Google Scholar] [CrossRef]
- Chung, H.S.; Kang, M.; Cho, C.; Park, S.; Kim, H.; Yoon, Y.S.; Kang, J.; Shin, M.K.; Hong, M.C.; Bae, H. Inhibition of lipopolysaccharide and interferon-gamma-induced expression of inducible nitric oxide synthase and tumor necrosis factor-alpha by Lithospermi radix in mouse peritoneal macrophages. J. Ethnopharmacol. 2005, 102, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Staniforth, V.; Wang, S.Y.; Shyur, L.F.; Yang, N.S. Shikonins, phytocompounds from Lithospermum erythrorhizon, inhibit the transcriptional activation of human TNF-α promoter in vivo. J. Biol. Chem. 2004, 279, 5877–5885. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Xu, Z.T.; Zhu, C.L.; Wang, J. Effect of different derivatives of shikonin from Lithospermum erythrorhizon against the pathogenic dental bacteria. Curr. Pharm. Anal. 2012, 8, 255–260. [Google Scholar] [CrossRef]
- Sasaki, K.; Abe, H.; Yoshizaki, F. In vitro antifungal activity of naphthoquinone derivatives. Biol. Pharm. Bull. 2002, 25, 669–670. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Yoon, J.S.; Song, S.Y. Protective effect of Lithospermum erythrorhizon on galactosamine induced liver injury. Korean J. Microsc. 2010, 40, 29–35. [Google Scholar]
- Wang, L.; Li, Z.; Zhang, X.; Wang, S.; Zhu, C.; Miao, J.; Chen, L.; Cui, L.; Qiao, H. Protective effect of shikonin in experimental ischemic stroke: Attenuated TLR4, p-p38MAPK, NF-κB, TNF-α and MMP-9 expression, up-regulated claudin-5 expression, ameliorated BBB permeability. Neurochem. Res. 2014, 39, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Seo, Y.C.; No, R.H.; Lee, H.Y. Improved cosmetic activity by optimizing the Lithospermum erythrorhizon extraction process. Cytotechnology 2015, 67, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Hisamichi, S.J.; Yoshizaki, F. Structures of new minor pigments and isolation of two isomers of shikonin derivatives from Lithospermum erythrorhizon Sieb. et Zucc. Shoyakugaku Zasshi 1982, 36, 154–159. [Google Scholar]
- Kim, G.S.; Kim, H.J.; Lee, D.Y.; Choi, S.M.; Lee, S.E.; Noh, H.J.; Choi, J.G.; Choi, S.I. Effects of supercritical fluid extract, shikonin and acetylshikonin from Lithospermum erythrorhizon on chondrocytes and MIA-induced osteoarthritis in rats. Korean J. Med. Crop Sci. 2013, 21, 466–473. [Google Scholar] [CrossRef]
- Zhou, W.; Jiang, H.D.G.L.; Peng, Y.; Li, S.S. Comparative study on enantiomeric excess of main akannin/shikonin derivatives isolated from the roots of three endemic Boraginaceae plants in China. Biomed. Chromatogr. 2011, 25, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.G.; Lu, Q.; Wen, H.D.; Cai, J.C. New asymmetric synthesis of alkannin and shikonin. Chin. Chem. Lett. 2005, 16, 465–467. [Google Scholar]
- Gebauer, M.; Saas, J.; Sohler, F.; Haag, J.; Söder, S.; Pieper, M.; Bartnik, E.; Beninga, J.; Zimmer, R.; Aigner, T. Comparison of the chondrosarcoma cell line SW1353 with primary human adult articular chondrocytes with regard to their gene expression profile and reactivity to IL-1β1. Osteoarthr. Cartil. 2005, 13, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Tetlow, L.C.; Adlam, D.J.; Woolley, D.E. Matrix metalloproteinase and proinflammatory cytokine production by chondrocytes of human osteoarthritic cartilage: Associations with degenerative changes. Arthritis Rheum. 2001, 44, 585–594. [Google Scholar] [CrossRef]
- Kobayashi, M.; Squires, G.R.; Mousa, A.; Tanzer, M.; Zukor, D.J.; Antoniou, J.; Feige, U.; Poole, A.R. Role of interleukin-1 and tumor necrosis factor α in matrix degradation of human osteoarthritic cartilage. Arthritis Rheum. 2005, 52, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Troeberg, L.; Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Biophys. Acta. 2012, 1824, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Burrage, P.S.; Mix, K.S.; Brinckerhoff, C.E. Matrix metalloproteinases: Role in arthritis. Front. Biosci. 2006, 11, 529–543. [Google Scholar] [CrossRef] [PubMed]
- Piecha, D.; Weik, J.; Kheil, H.; Becher, G.; Timmermann, A.; Jaworski, A.; Burger, M.; Hofmann, M.W. Novel selective MMP-13 inhibitors reduce collagen degradation in bovine articular and human osteoarthritis cartilage explants. Inflamm. Res. 2010, 59, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Lipari, L.; Gerbino, A. Expression of gelatinases (MMP-2, MMP-9) in human articular cartilage. Int. J. Immunopathol. Pharmacol. 2013, 26, 817–823. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Matrix Metalloproteinase (MMP) | IC50 Value (μM) |
|---|---|
| MMP-1 | 58.7 |
| MMP-2 | 60.8 |
| MMP-3 | >100 |
| MMP-9 | >100 |
| MMP-13 | 63.3 |
| TIMP-2 | – |
| Gene Code | Accession ID | Sequence (5′→3′) | Temperature (°C) | Product Length (bp) | |
|---|---|---|---|---|---|
| MMP-1 | NM_001145938.1 | Forward | AGTGACTGGGAAACCAGATGA | 57 | 159 |
| Reverse | CGTCTTGGCAAATCTGGCCTGTAA | ||||
| MMP-2 | NM_001127891.2 | Forward | GCAGTGGGGGCTTAAGAAGA | 57 | 969 |
| Reverse | AGCCGTACTTGCCATCCTTC | ||||
| MMP-3 | NM_002422.3 | Forward | ATTCCATGGAGCCAGGCTTTC | 57 | 142 |
| Reverse | CATTTGGGTCAAACTCCACTGTG | ||||
| MMP-9 | NM_004994.2 | Forward | CATCCGGCACCTCTATGGTC | 57 | 637 |
| Reverse | CATCGTCCACCGGACTCAAA | ||||
| MMP-13 | NM_002427.3 | Forward | AAATTATGGAGGAGATGCCCATT | 57 | 125 |
| Reverse | TCCTTGGAGTGGTCAAGACCTAA | ||||
| TIMP-2 | NM_003255.4 | Forward | GTAGTGATCAGGGCCAAAGC | 57 | 160 |
| Reverse | GGGGGCCGTGATAAACT | ||||
| GAPDH | NM_001256799.2 | Forward | AGAAGGCTGGGGCTCATTTG | 52 | 271 |
| Reverse | AGGGGCCATCAGTCTTC | ||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.Y.; Choi, S.-I.; Han, S.H.; Lee, Y.-J.; Choi, J.-G.; Lee, Y.-S.; Choi, J.H.; Lee, S.-E.; Kim, G.-S. Potential of Pseudoshikonin I Isolated from Lithospermi Radix as Inhibitors of MMPs in IL-1β-Induced SW1353 Cells. Int. J. Mol. Sci. 2016, 17, 1350. https://doi.org/10.3390/ijms17081350
Lee DY, Choi S-I, Han SH, Lee Y-J, Choi J-G, Lee Y-S, Choi JH, Lee S-E, Kim G-S. Potential of Pseudoshikonin I Isolated from Lithospermi Radix as Inhibitors of MMPs in IL-1β-Induced SW1353 Cells. International Journal of Molecular Sciences. 2016; 17(8):1350. https://doi.org/10.3390/ijms17081350
Chicago/Turabian StyleLee, Dae Young, Soo-Im Choi, Se Hee Han, Ye-Joo Lee, Jong-Gil Choi, Young-Seob Lee, Je Hun Choi, Seung-Eun Lee, and Geum-Soog Kim. 2016. "Potential of Pseudoshikonin I Isolated from Lithospermi Radix as Inhibitors of MMPs in IL-1β-Induced SW1353 Cells" International Journal of Molecular Sciences 17, no. 8: 1350. https://doi.org/10.3390/ijms17081350