
Analysis of Protein Composition and Bioactivity of Neoponera villosa Venom (Hymenoptera: Formicidae)

 , , and
, , and
Abstract
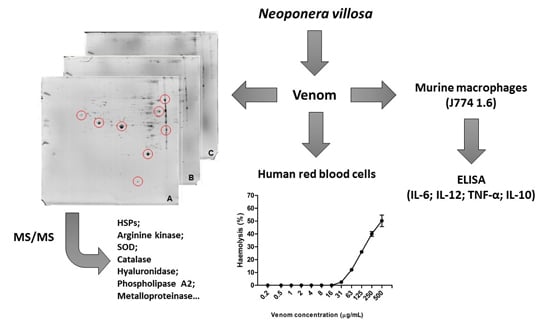
:
1. Introduction
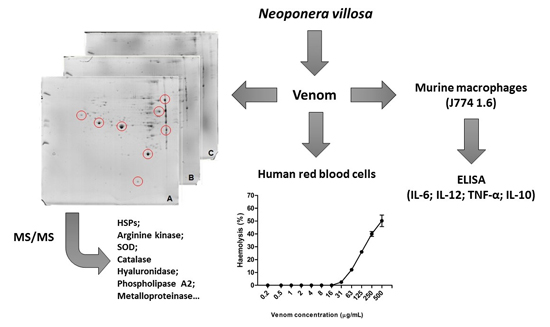
2. Results
2.1. Doses of Protein
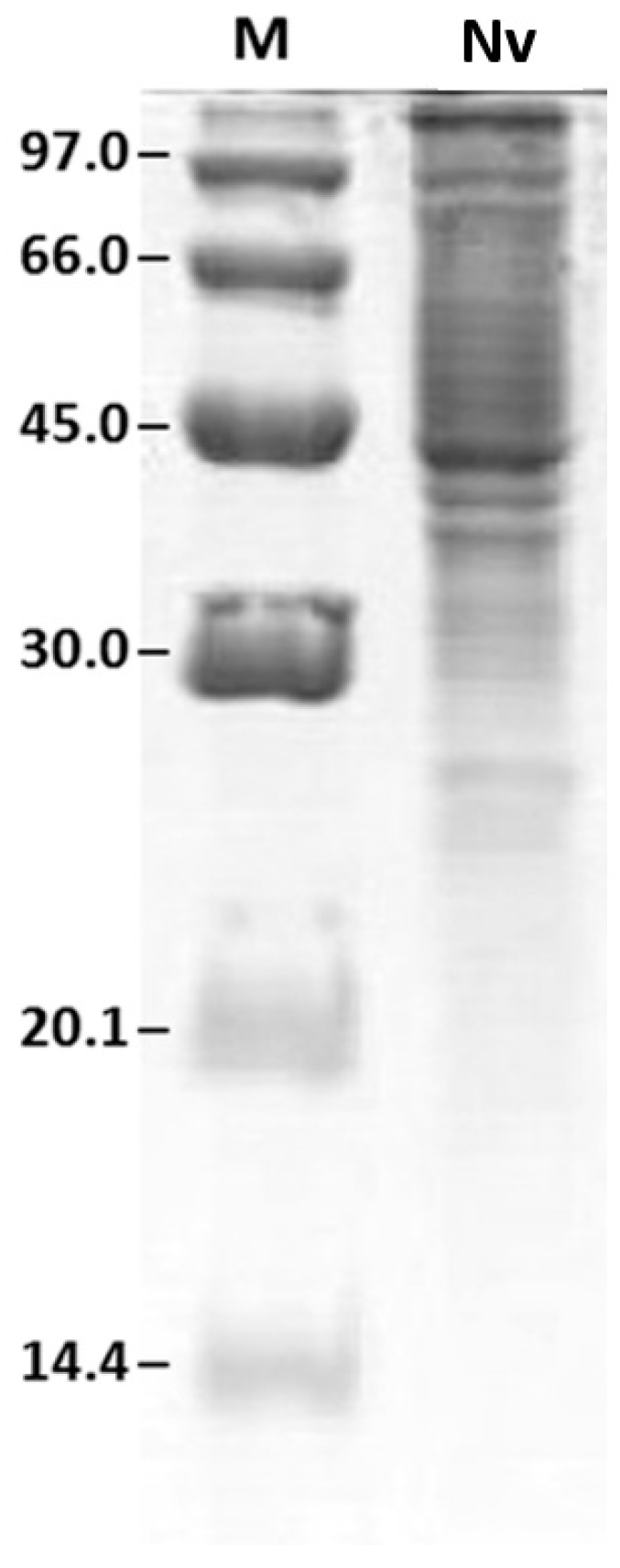
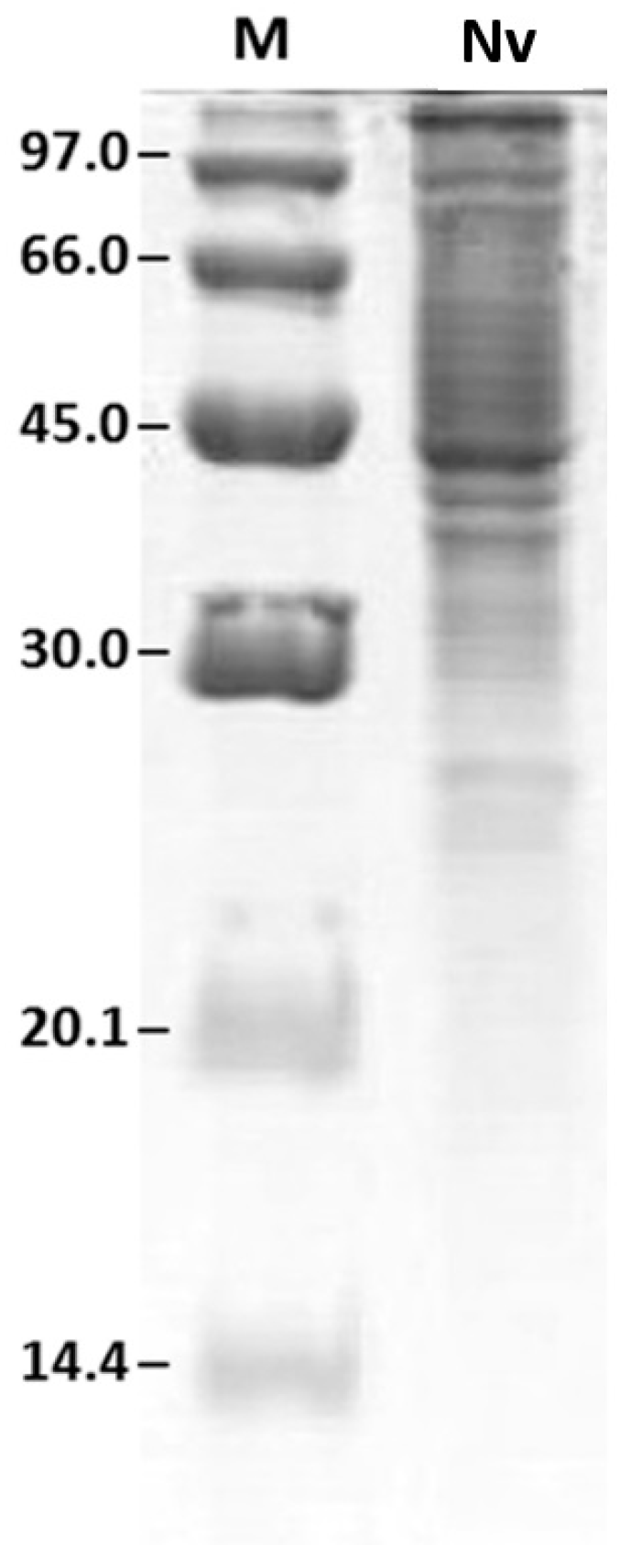
2.2. Protein Profile Obtained by SDS-PAGE
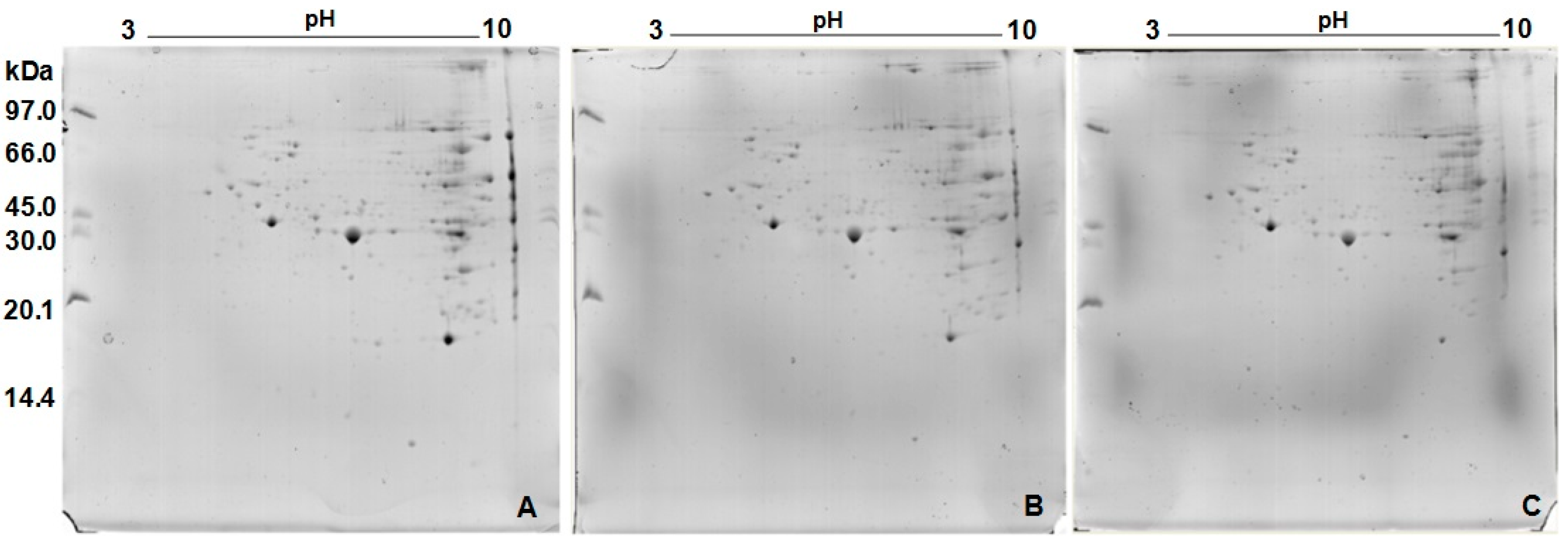
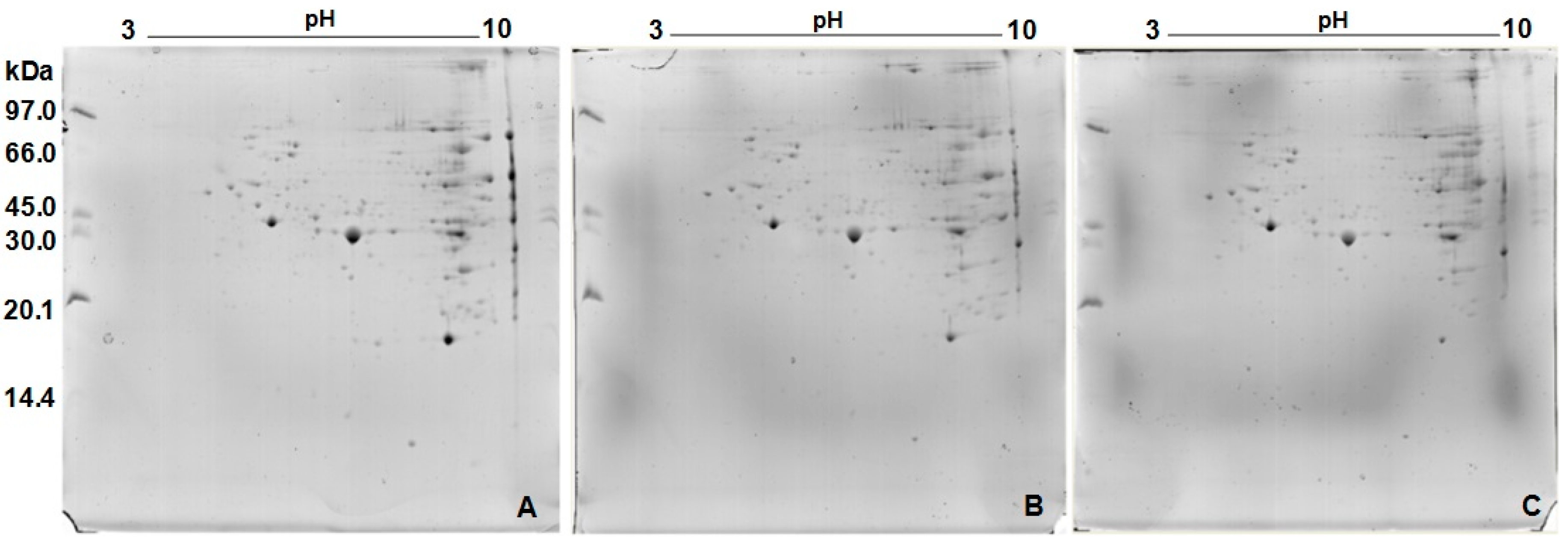
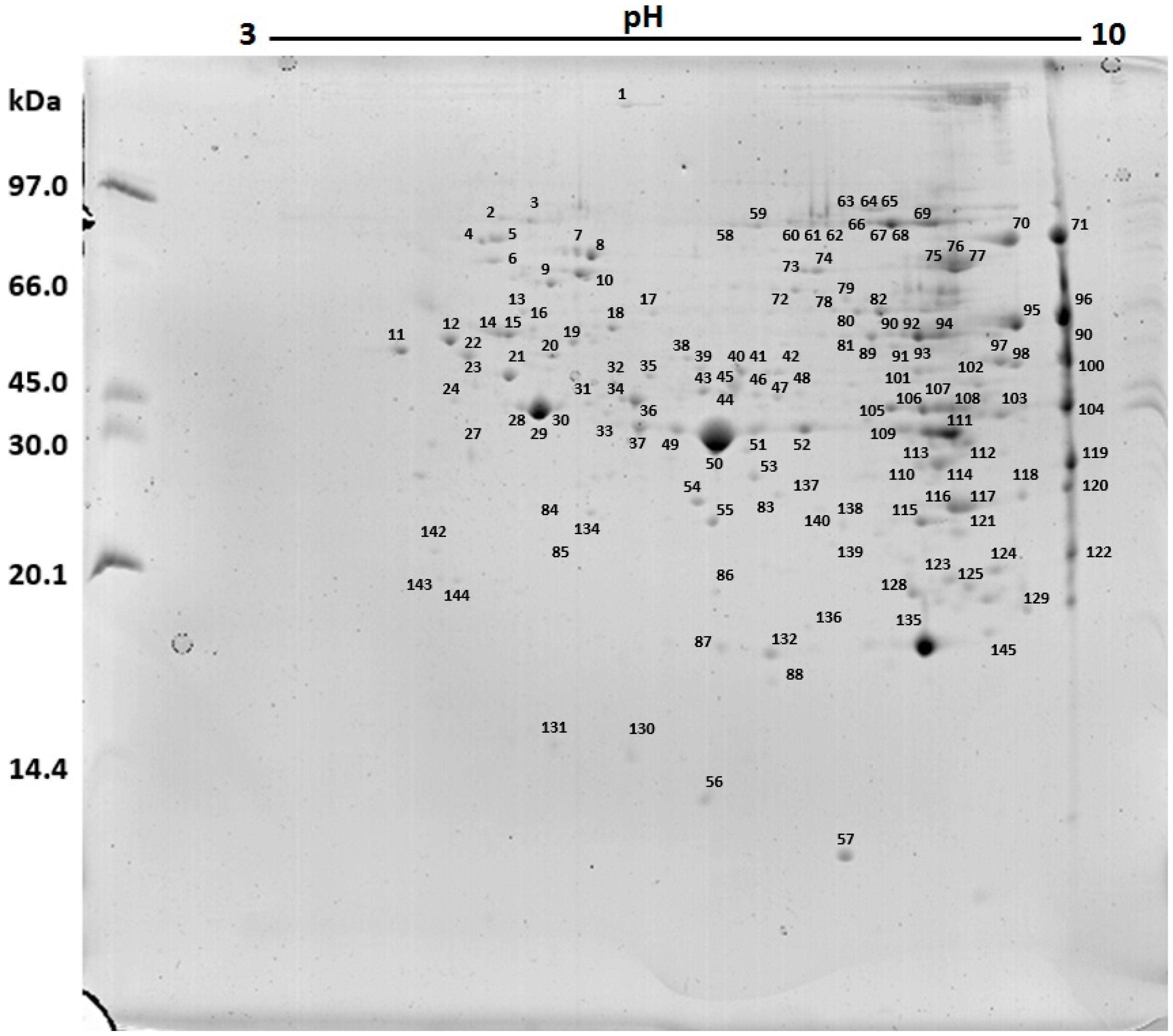
2.3. Protein Profile Obtained by Two-Dimensional Electrophoresis (2D)
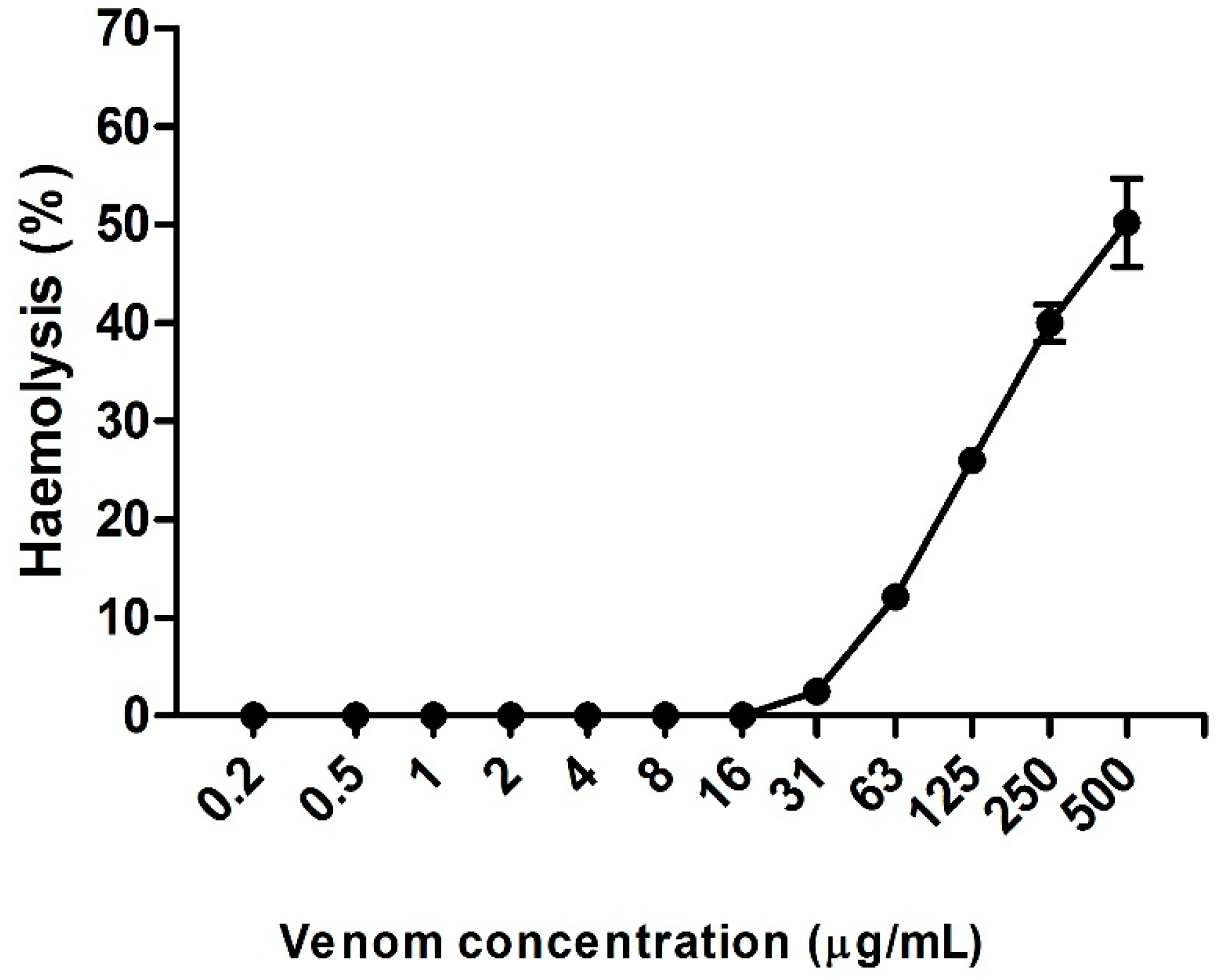
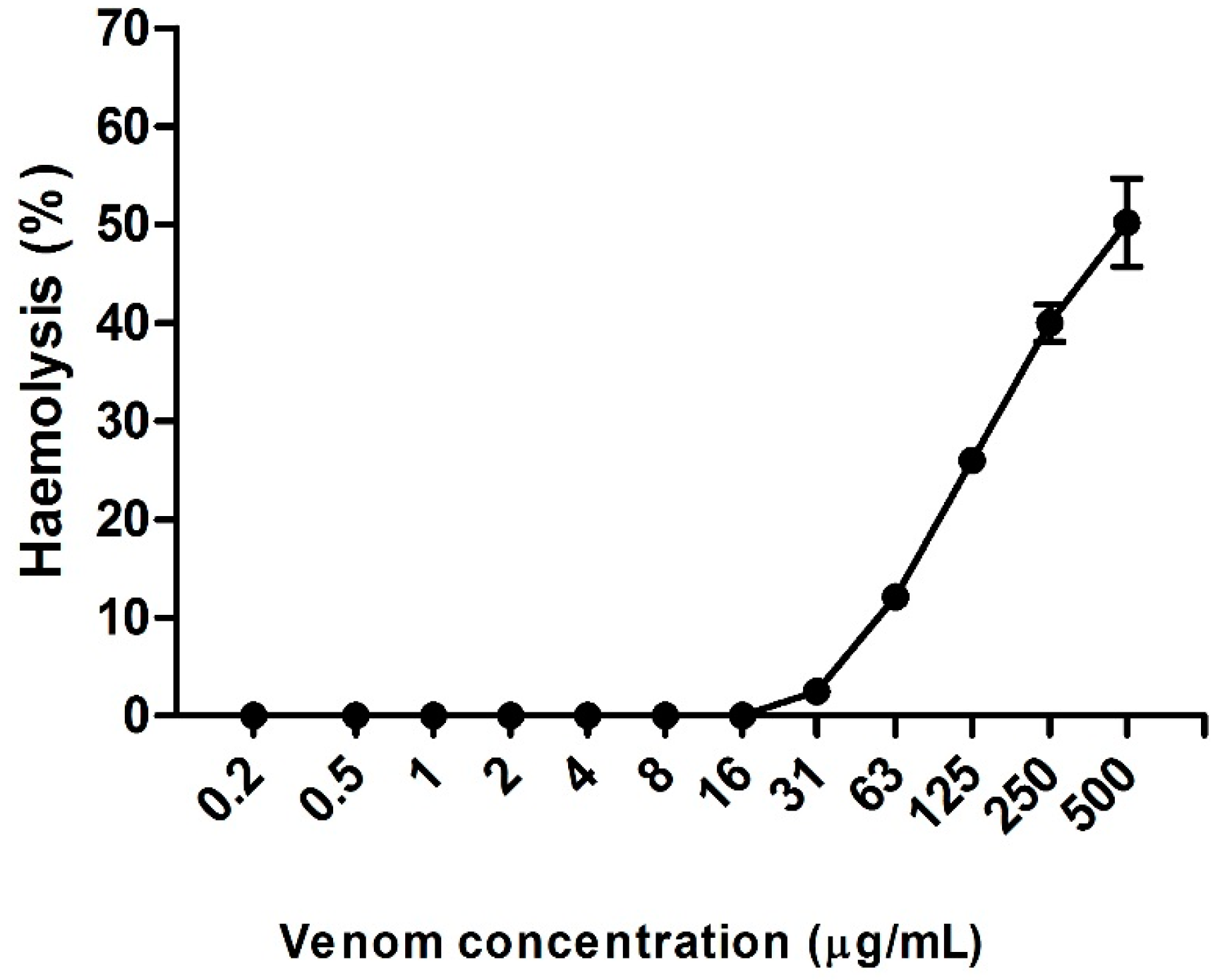
2.4. Hemolytic Activity Induced by Neoponera villosa Venom
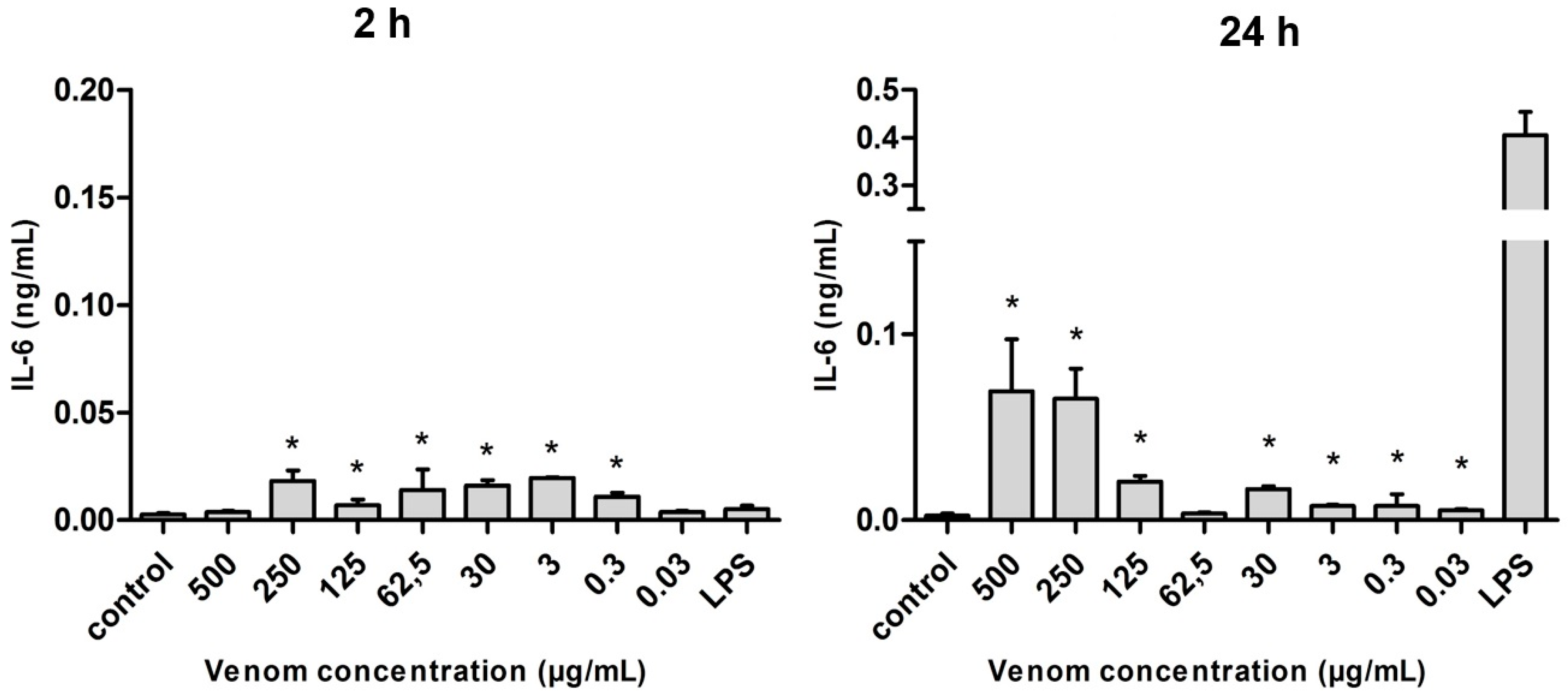
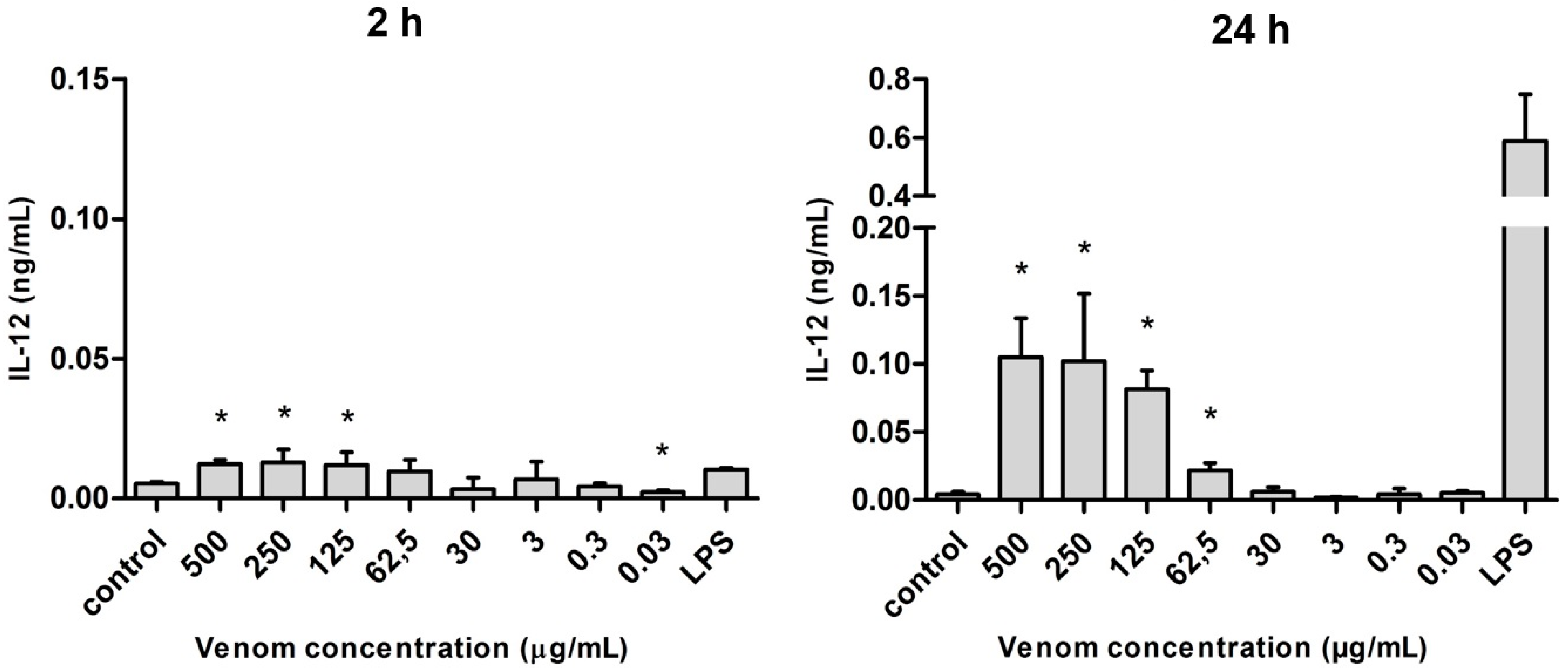
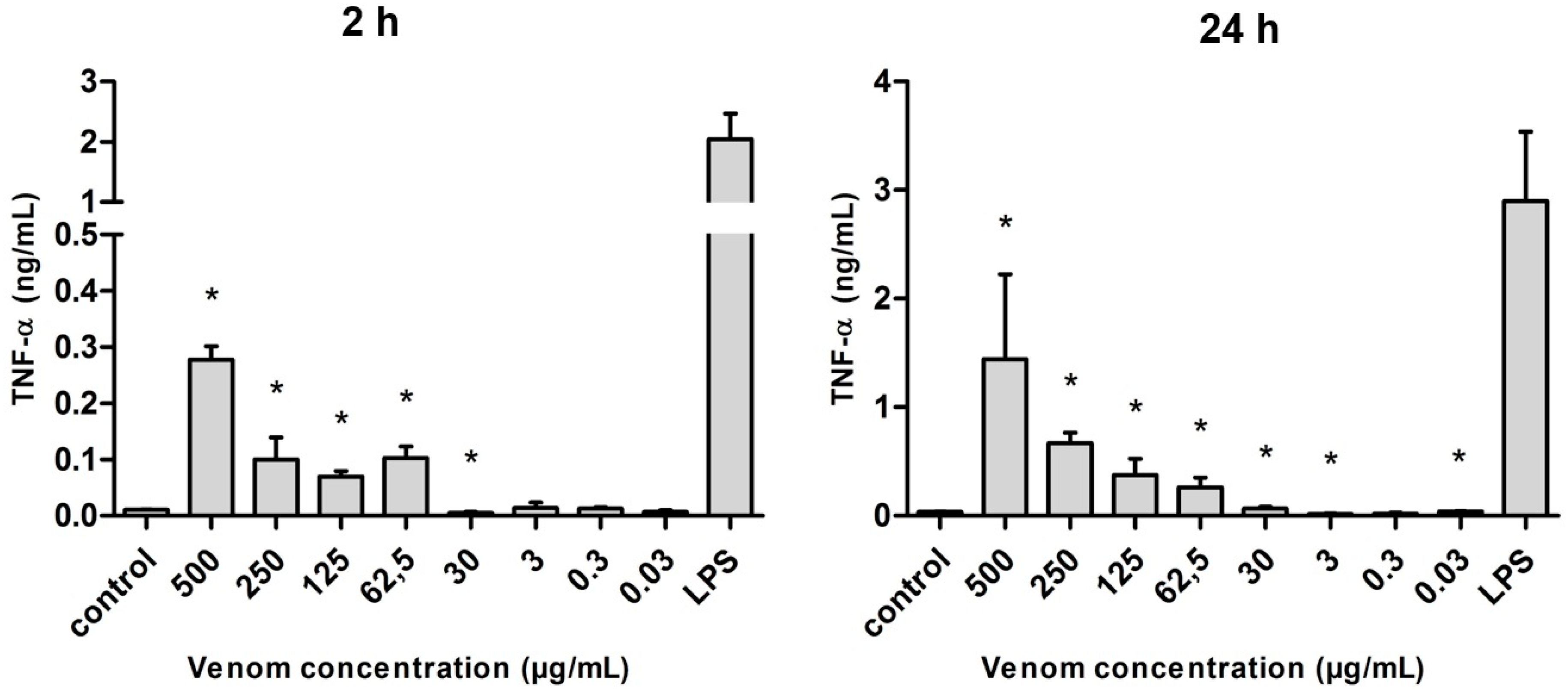
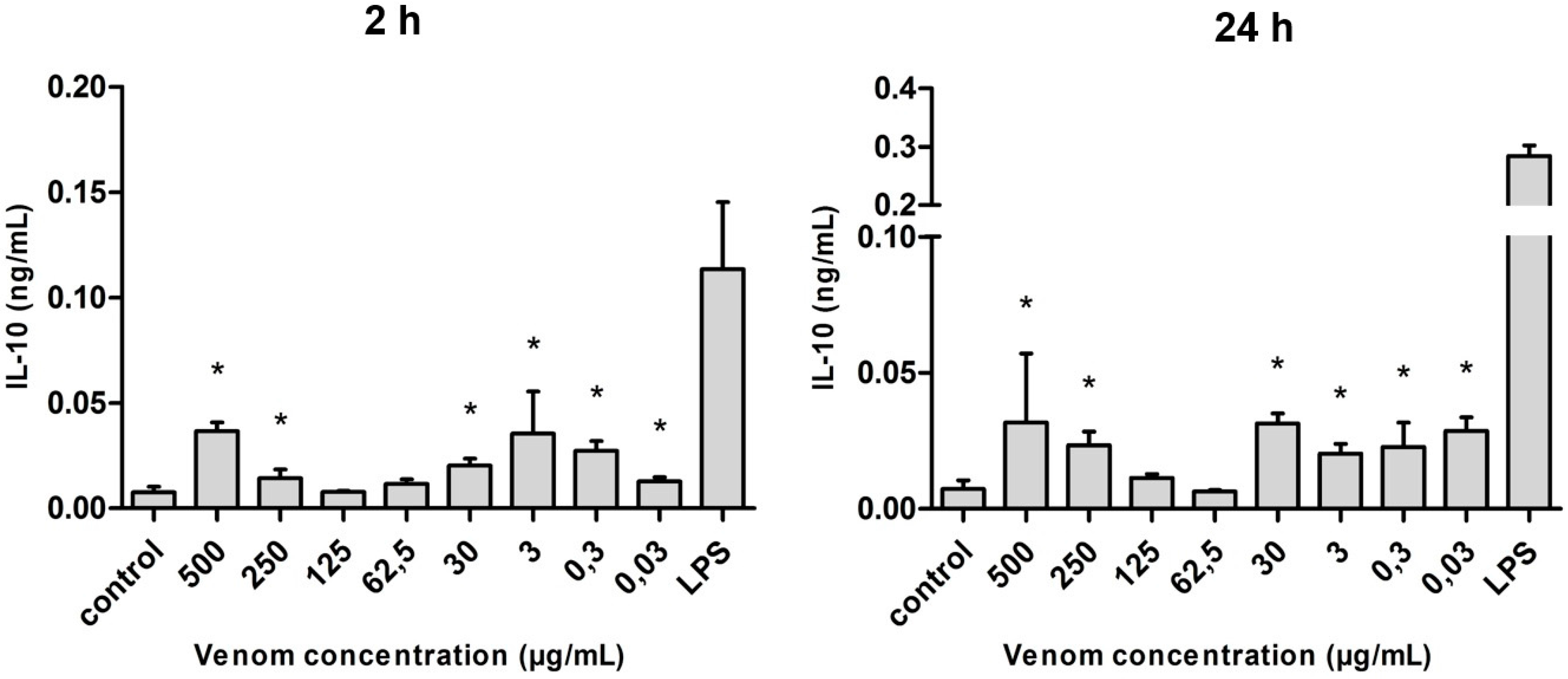
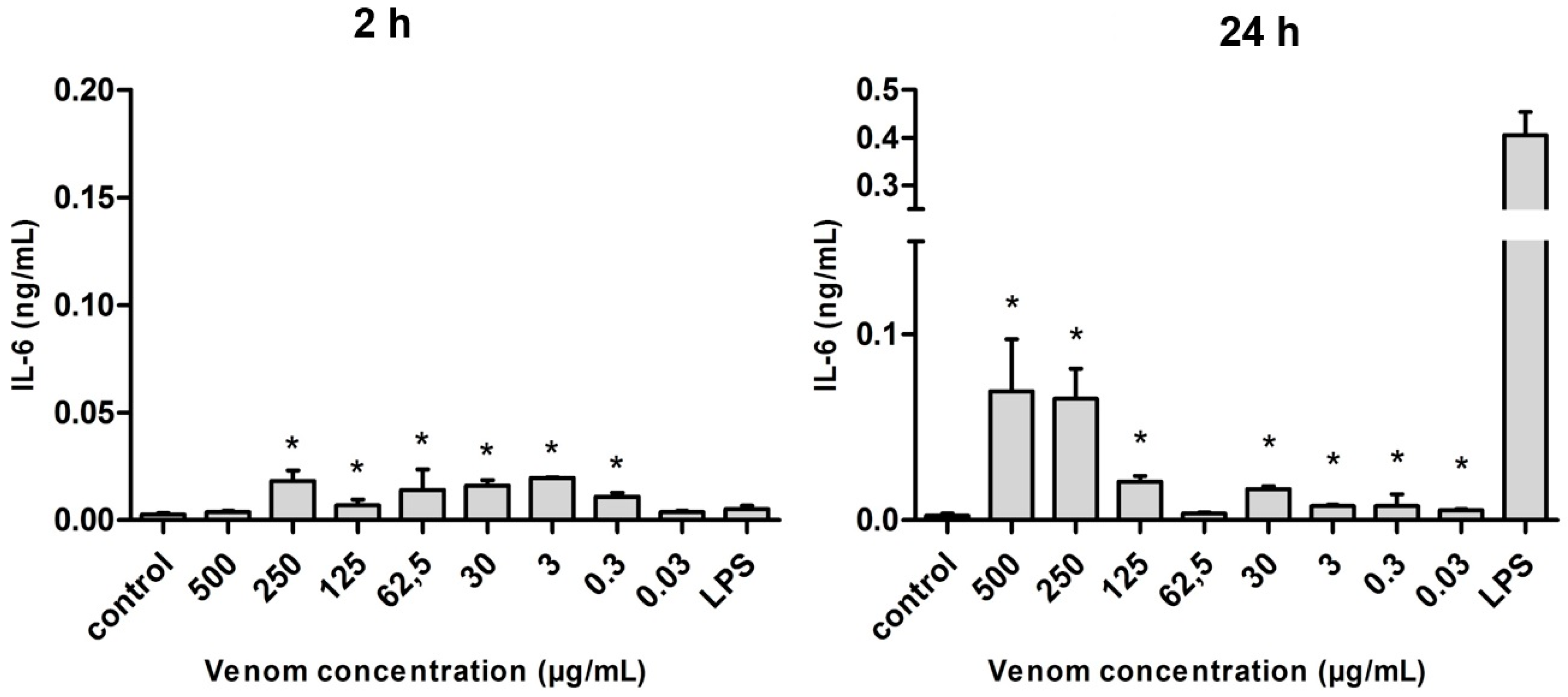
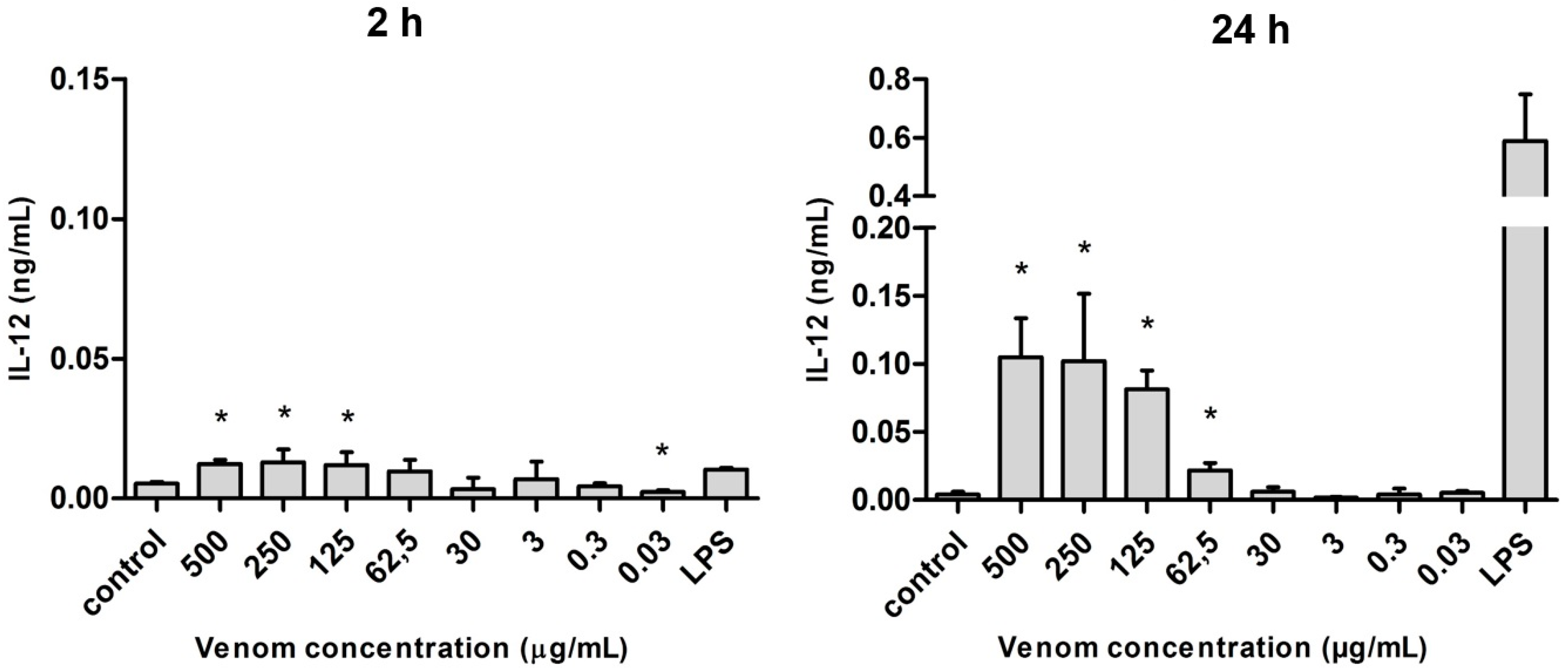
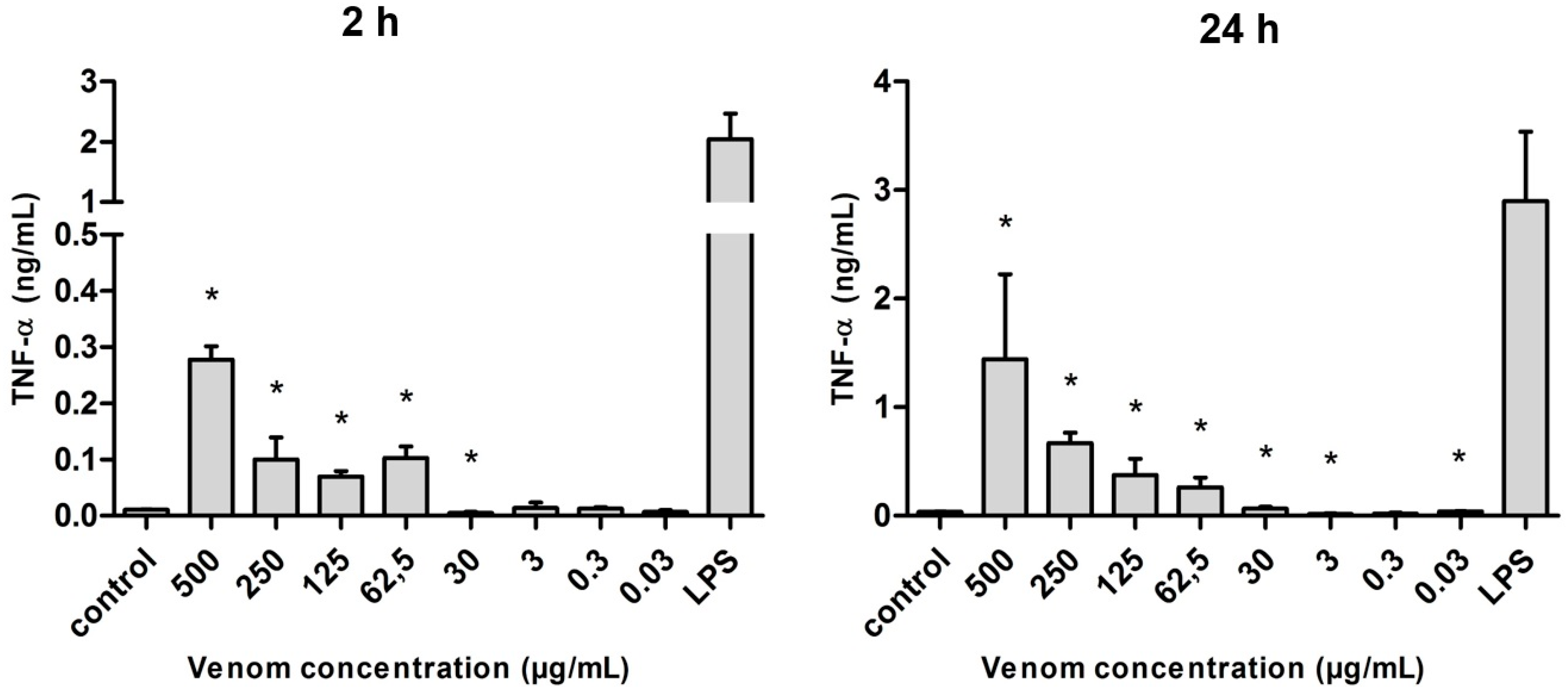
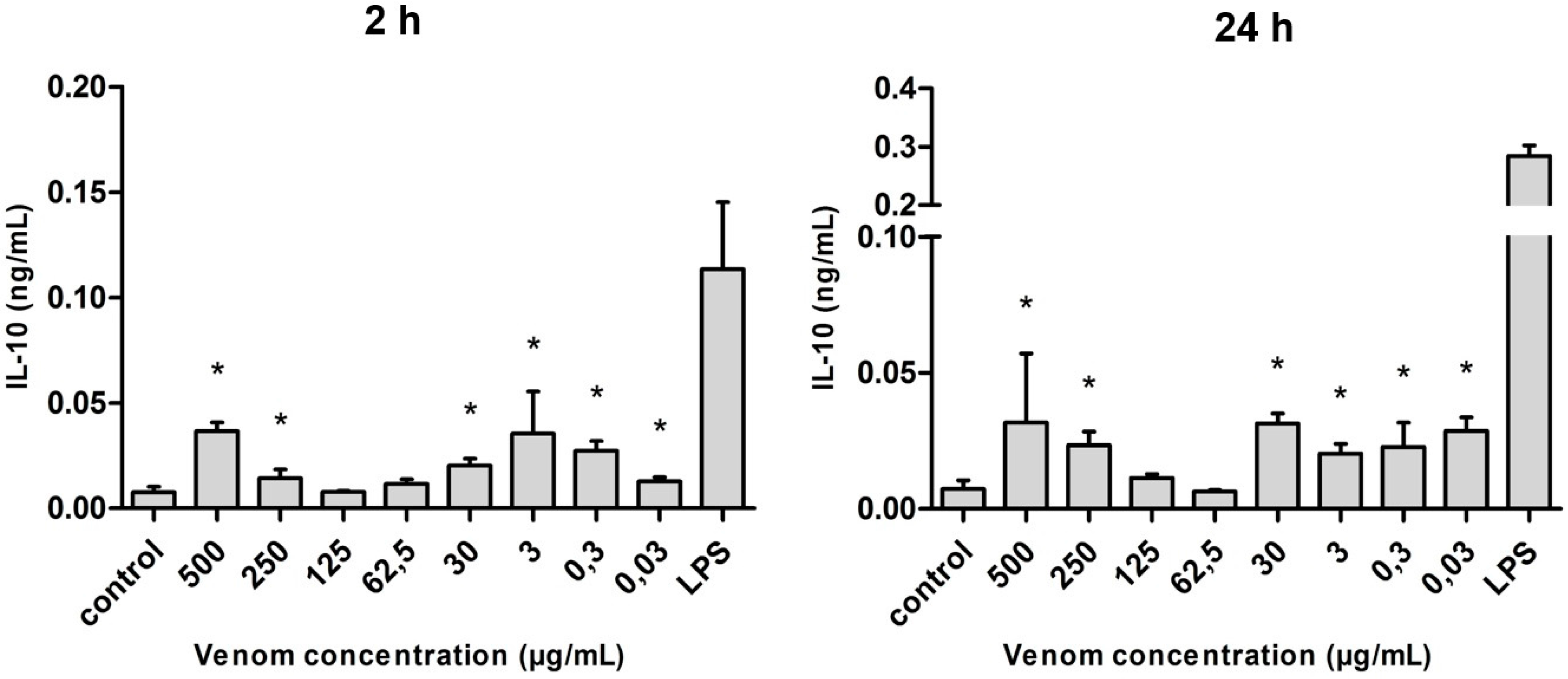
2.5. Induction of Cytokines in J774 Macrophages in the Presence of Neoponera villosa Venom
3. Discussion
4. Materials and Methods
4.1. Venom Extraction
4.2. Protein Extraction
4.3. Protein Quantification
4.4. Electrophoresis (SDS-PAGE)
4.5. Two-Dimensional Electrophoresis (2D-SDS-PAGE)
4.6. Protein Visualization and Image Analysis
4.7. Spot Excision and Triptych Digestion
4.8. Peptide Sequencing and Protein Identification
4.9. Hemolytic Activity
4.10. Murine Macrophages Culture and Stimulus with Neoponera villosa Venom
4.11. Cytokines Doses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 2D | Two-dimensional |
| BCRJ | Rio de Janeiro Cell Bank |
| CEPLAC | Executive Committee of the Cocoa Crop |
| CHAPS | 3-((3-Cholamidopropyl)dimethylammonio)-1-propanesulfonate |
| CI | Confidence interval |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DTT | Dithiothreitol |
| ELISA | Enzyme linked immuno sorbent assay |
| FBS | Fetal bovine serum |
| HSP | Heat shock proteins |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| MALDI-TOF | Matrix-Assisted Laser Desorption/Ionization Time-Of-Flight |
| MAPK | Mitogen-activated protein kinase |
| MW | Molecular weight |
| NCBI | National Center for Biotechnology Information |
| NO | Nitric oxide |
| PAGE | Polyacrylamide gel electrophoresis |
| PBS | Phosphate buffered saline |
| pI | Isoelectric point |
| SOD | Superoxide dismutase |
| SDS | Sodium dodecyl sulfate |
| TCA | Trichloroacetic acid |
| TNF | Tumor necrosis factor |
| UESC | State University of Santa Cruz |
| UPLC | Ultra performance liquid chromatography |
References
- Steen, C.J.; Janniger, C.K.; Schutzer, S.E.; Schwartz, R.A. Insect sting reactions to bees, wasps, and ant. Int. J. Dermatol. 2005, 44, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, K.T.; Flood, A.A. Hymenoptera Stings. Clin. Tech. Small Anim. Pract. 2006, 21, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Escoubas, P.; King, G.F. Venomics as a drug discovery platform. Expert Rev. Proteom. 2009, 6, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Nentwig, L.; Stöcklin, R.; Nentwig, W. Venom composition and strategies in spiders: Is everything possible? Adv. Insect Physiol. 2011, 40, 1–86. [Google Scholar]
- Georgieva, D.; Seifert, J.; Öhler, M.; von Bergen, M.; Spencer, P.; Arni, R.K.; Genov, N.; Betzel, C. Pseudechis australis venomics: Adaptation for a defense against microbial pathogens and recruitment of body transferrin. J. Proteome Res. 2011, 10, 2440–2464. [Google Scholar] [CrossRef] [PubMed]
- De Graaf, D.C.; Aerts, M.; Danneels, E.; Devreese, B. Bee, wasp and ant venomics pave the way for a component-resolved diagnosis of sting allergy. J. Proteom. 2009, 72, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.D.; Pieroni, M.; Menegasso, A.R.S.; Pinto, J.R.A.S.; Palma, M.S. A new scenario of bioprospecting of Hymenoptera venoms through proteomic approach. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 364–377. [Google Scholar]
- Hoffman, D.R.; Sakell, R.H.; Schmidt, M. Sol i 1, the phospholipase allergen of imported fire ant venom. J. Allergy Clin. Immunol. 2005, 115, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, E.F.; Camargos, T.S.; Zamudio, F.Z.; Silva, L.P.; Bloch, C., Jr.; Caixeta, F.; Schwartz, C.A.; Possani, L.D. Mass spectrometry analysis, amino acid sequence and biological activity of venom components from the Brazilian scorpion Opisthacanthus cayaporum. Toxicon 2008, 15, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Rates, B.; Bemquerer, M.P.; Richardson, M.; Borges, M.H.; Morales, R.A.; de Lima, M.E.; Pimenta, A.M. Venomic analyses of Scolopendra viridicornis nigra and Scolopendra angulata (Centipede, Scolopendromorpha): Shedding light on venoms from a neglected group. Toxicon 2007, 49, 810–826. [Google Scholar] [CrossRef] [PubMed]
- Rates, B.; Silva, L.P.; Ireno, I.C.; Leite, F.S.; Borges, M.H.; Bloch, C., Jr.; de Lima, M.E.; Pimenta, A.M. Peptidomic dissection of the skin secretion of Phasmahyla jandaia (Bokermann and Sazima, 1978) (Anura, Hylidae, Phyllomedusinae). Toxicon 2011, 57, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Fialho, E.M.; Maciel, M.C.; Silva, A.C.; Reis, A.S.; Assunção, A.K.; Fortes, T.S.; Silva, L.A.; Guerra, R.N.; Kwasniewski, F.H.; Nascimento, F.R. Immune cells recruitment and activation by Tityus serrulatus scorpion venom. Toxicon 2011, 58, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.B.; McClendon, D.; Desaiah, D.; Goddard, J.; Lister, A.; Moffitt, J.; Meer, R.K.; de Shazo, R.; Lee, K.S.; Rockhold, R.W. Fire ant venom alkaloid, isosolenopsin A, a potent and selective inhibitor of neuronal nitric oxide synthase. Int. J. Toxicol. 2003, 22, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L.; Hernández Cruz, A.; Coronas, F.I.; Possani, L.D. Toxin gamma from Tityus serrulatus scorpion venom plays an essential role in immunomodulation of macrophages. Toxicon 2007, 50, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Ponvert, C.; le Courvoisier, C.; Weill, B.; Bloch, E.; Paupe, J.; Scheinmann, P. Kinetics of plasma cytokine levels in children hyposensitized with wasp venom. Cytokine 2001, 15, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.S.; Kim, S.K.; Han, J.B.; Ahn, H.J.; Bae, H.; Min, B.I. Effects of bee venom on the pro-inflammatory responses in RAW264.7 macrophage cell line. J. Ethnopharmacol. 2005, 99, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Zanchet, E.M.; Longo, I.; Cury, Y. Involvement of spinal neurokinins, excitatory amino acids, proinflammatory cytokines, nitric oxide and prostanoids in pain facilitation induced by Phoneutria nigriventer spider venom. Brain Res. 2004, 1021, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, A.W.; Avigad, L.S.; Schmidt, J.O. A hemolytic polypeptide from the venom of the red harvester ant, Pogonomyrmex barbatus. Toxicon 1980, 18, 271–278. [Google Scholar] [CrossRef]
- De Souza, A.L.; Malaque, C.M.; Sztajnbok, J.; Romano, C.C.; Duarte, A.J.; Seguro, A.C. Loxosceles venom-induced cytokine activation, haemolysis, and acute kidney injury. Toxicon 2008, 51, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.A.; Shattuck, S.O. The higher classification of the ant subfamily ponerinae (Hymenoptera: Formicidae), with a review of ponerine ecology and behavior. Zootaxa 2014, 3817, 1–242. [Google Scholar] [CrossRef] [PubMed]
- Schimidt, J.O.; Blum, M.; Overal, W. Hemolytic activities of stinging insect venoms. Arch. Insect Biochem. Physiol. 1984, 1, 155–160. [Google Scholar] [CrossRef]
- Lopez, L.C.; Morgan, E.D. Explanation of bitter taste of venom of ponerine ant, Pachycondyla apicalis. J. Chem. Ecol. 1997, 23, 705–712. [Google Scholar] [CrossRef]
- Orivel, J.; Dejean, A. Comparative effect of the venoms of ants of the genus Pachycondyla (Hymenoptera: Ponerinae). Toxicon 2001, 39, 195–201. [Google Scholar] [CrossRef]
- Mill, A.E. Predation by the ponerine ant Pachycondyla commutata on termites of the genus Syntermes in Amazonian rain forest. J. Nat. Hist. 1982, 18, 405–410. [Google Scholar] [CrossRef]
- Lee, E.K.; Jeong, K.Y.; Lyu, D.P.; Lee, Y.W.; Sohn, J.H.; Lim, K.J.; Hong, C.S.; Park, J.W. Characterization of the major allergens of Pachycondyla chinensis in ant sting anaphylaxis patients. Clin. Exp. Allergy 2009, 39, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Dkhil, M.A.; Abdel-Baki, A.S.; Al-Quraishi, S.; Al-Khalifa, M. Anti-inflammatory activity of the venom from samsum ants Pachycondyla sennaarensis. Afr. J. Pharmacol. 2010, 4, 115–118. [Google Scholar]
- Mackay, W.P.; Mackay, E.E. The Systematics and Biology of the New World Ants of the Genus Pachycondyla (Hymenoptera: Formicidae), 1st ed.; The Edwin Mellen Press: Lewiston, NY, USA, 2010; pp. 571–579. [Google Scholar]
- Lucas, C.; Fresneau, D.; Kolmer, K.; Heinze, J.; Delabie, J.H.C.; Pho, D.B. A multidisciplinary approach to discriminating different taxa in the species complex Pachycondyla villosa (Formicidae). Biol. J. Linn. Soc. 2002, 65, 249–259. [Google Scholar] [CrossRef]
- Mariano, C.S.F.; Delabie, J.H.C.; Santos, J.R.M.; Pompolo, S.G. Karyotype evolution in Neotropical Pachycondyla spp. (Ponerinae). Biológico 2007, 69, 409–412. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Silva, M.F.; Mota, C.M.; Miranda, V.S.; Cunha, A.O.; Silva, M.C.; Naves, K.S.C.; Oliveira, F.; Silva, D.A.O.; Mineo, T.W.P.; Santiago, F.M. Biological and enzymatic characterization of proteases from crude venom of the ant Odontomachus bauri. Toxins 2015, 7, 5114–5128. [Google Scholar] [CrossRef] [PubMed]
- Costa Manso, E.; Croce, M.; Pinto, J.R.; Souza Santos, K.; Delazari Santos, L.; Baptista Dias, N.; Palma, M.S. Anaphylaxis due to Pachycondyla goeldii ant: A case report. J. Investig. Allergol. Clin. Immunol. 2010, 20, 352–353. [Google Scholar] [PubMed]
- Dos Santos Pinto, J.R.; Fox, E.G.; Saidemberg, D.M.; Santos, L.D.; da Silva Menegasso, A.R.; Costa-Manso, E.; Machado, E.A.; Bueno, O.C.; Palma, M.S. Proteomic view of the venom from the fire ant Solenopsis invicta Buren. J. Proteome Res. 2012, 11, 4643–4653. [Google Scholar] [CrossRef] [PubMed]
- Leluk, J.; Schmidt, J.; Jones, D. Comparative studies on the protein composition of hymenopteran venom reservoirs. Toxicon 1989, 27, 105–114. [Google Scholar] [CrossRef]
- Peiren, N.; de Graaf, D.C.; Vanrobaeys, F.; Danneels, E.L.; Devreese, B.; Van Beeumen, J.; Jacobs, F.J. Proteomic analysis of the honey bee worker venom gland focusing on the mechanisms of protection against tissue damage. Toxicon 2008, 52, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Tsan, M.F.; Gao, B. Cytokine function of heat shock proteins. Am. J. Physiol. Cell Physiol. 2004, 286, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Tsan, M.F.; Gao, B. Heat shock protein and innate immunity. Cell. Mol. Immunol. 2004, 1, 274–279. [Google Scholar] [PubMed]
- Asea, A.; Kraeft, S.K.; Kurt-Jones, E.A.; Stevenson, M.A.; Chen, L.B.; Finberg, R.W.; Koo, G.C.; Calderwood, S.K. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat. Med. 2000, 6, 435–442. [Google Scholar] [PubMed]
- Retzlaff, C.; Yamamoto, Y.; Hoffman, P.S.; Friedman, H.; Klein, T.W. Bacterial heat shock proteins directly induce cytokine mRNA and interleukin-1 secretion in macrophage cultures. Infect. Immun. 1994, 62, 5689–5693. [Google Scholar] [PubMed]
- Galdiero, M.; de l’Ero, G.C.; Marcatili, A. Cytokine and adhesion molecule expression in human monocytes and endothelial cells stimulated with bacterial heat shock proteins. Infect. Immun. 1997, 65, 699–707. [Google Scholar] [PubMed]
- Zhu, A.; Wang, Z.K.; Beavis, R. Structural studies of α-N-acetylgalactosaminidase: Effect of glycosylation on the level of expression, secretion efficiency, and enzyme activity. Arch. Biochem. Biophys. 1998, 352, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, S.B.; Nagasawa, H.; Uto, Y.; Hori, H. Tumor cell α-N-acetylgalactosaminidase activity and its involvement in GcMAF-related macrophage activation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 132, 1–8. [Google Scholar] [CrossRef]
- Cho, W.; Dhadialla, T.S.; Raikhel, A.S. Purification and characterization of a lysosomal aspartic protease with cathepsin D activity from the mosquito. Insect Biochem. 1991, 21, 165–176. [Google Scholar] [CrossRef]
- Lkhider, M.; Castino, R.; Bouguyon, E.; Isidoro, C.; Ollivier-Bousquet, M. Cathepsin D released by lactating rat mammary epithelial cells is involved in prolactin cleavage under physiological conditions. J. Cell Sci. 2004, 117, 5155–5164. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhu, J.Y.; Fang, Q.; Wang, L.; Hu, C.; Ye, G.Y. Proteomic analysis of the venom from the endoparasitoid wasp Pteromalus puparum (Hymenoptera: Pteromalidae). Arch. Insect Biochem. Physiol. 2010, 75, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.D.; Santos, K.S.; Pinto, J.R.; Dias, N.B.; de Souza, B.M.; dos Santos, M.F.; Perales, J.; Domont, G.B.; Castro, F.M.; Kalil, J.E.; et al. Profiling the proteome of the venom from the social wasp Polybia paulista: A clue to understand the envenoming mechanism. J. Proteome Res. 2010, 9, 3867–3877. [Google Scholar] [CrossRef] [PubMed]
- García-Orozco, K.D.; Aispuro-Hernández, E.; Yepiz-Plascencia, G.; Calderón-de-la-Barca, A.M.; Sotelo-Mundo, R.R. Molecular characterization of arginine kinase, an allergen from the shrimp Litopenaeus vannamei. Int. Arch. Allergy Immunol. 2007, 144, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.W.; Cao, M.J.; Cai, Q.F.; Ruan, M.M.; Mao, H.Y.; Su, W.J.; Liu, G.M. Purification, cloning, and immunological characterization of arginine kinase, a novel allergen of Octopus fangsiao. J. Agric. Food Chem. 2012, 60, 2190–2199. [Google Scholar] [CrossRef] [PubMed]
- Rosmilah, M.; Shahnaz, M.; Zailatul, H.M.; Noormalin, A.; Normilah, I. Identification of tropomyosin and arginine kinase as major allergens of Portunus pelagicus (blue swimming crab). Trop. Biomed. 2012, 29, 467–478. [Google Scholar] [PubMed]
- Guarneri, F.; Guarneri, C.; Guarneri, B.; Benvenga, S. In silico identification of potential new latex allergens. Clin. Exp. Allergy 2006, 36, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Marković-Housley, Z.; Miglierini, G.; Soldatova, L.; Rizkallah, P.J.; Müller, U.; Schirmer, T. Crystal structure of hyaluronidase, a major allergen of bee venom. Structure 2000, 8, 1025–1035. [Google Scholar] [CrossRef]
- Gmachl, M.; Kreil, G. Bee venom hyaluronidase is homologous to a membrane protein of mammalian sperm. Proc. Natl. Acad. Sci. USA 1993, 90, 3569–3573. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R. Hymenoptera venom allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef]
- Marques, P.P.; Esteves, A.; Lancellotti, M.; Ponce-Soto, L.A.; Marangoni, S. Novel acidic phospholipase A2 from Porthidium hyoprora causes inflammation with mast cell rich infiltrate. Biochem. Biophys. Rep. 2015, 1, 78–84. [Google Scholar] [CrossRef]
- Sampaio, S.C.; Rangel-Santos, A.C.; Peres, C.M.; Curi, R.; Cuty, Y. Inhibitory effect of phospholipase A2 isolated from Crotalus durissus terrificus venom on macrophage function. Toxicon 2011, 45, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A. Snake venom metalloproteinases: Their role in the pathogenesis of local tissue damage. Biochimie 2000, 82, 841–850. [Google Scholar] [CrossRef]
- Koya, S.; Crenshaw, D.; Agarwal, A. Rhabdomyolysis and acute renal failure after fire ant bites. J. Gen. Intern. Med. 2007, 22, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Díaz, C. Hemorrhage induced by snake venom metalloproteinases: Biochemical and biophysical mechanisms involved in microvessel damage. Toxicon 2005, 45, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Woo, H.A.; Kil, I.S.; Bae, S.H. Peroxiredoxin functions as a peroxidase and a regulator and sensor of local peroxides. J. Biol. Chem. 2012, 287, 4403–4410. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Fang, Y.; Han, B.; Lu, X.; Zhou, T.; Feng, M.; Li, J. Proteome and phosphoproteome analysis of honeybee (Apis mellifera) venom collected from electrical stimulation and manual extraction of the venom gland. BMC Genom. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.F.C.; Huang, C.; Chong, C.M.; Leung, S.W.; Prieto-da-Silva, A.R.B.; Havt, A.; Quinet, Y.P.; Martins, A.M.C.; Lee, S.M.Y.; Rádis-Baptista, G. Transcriptome analysis in venom gland of the predatory giant ant Dinoponera quadriceps: Insights into the polypeptide toxin arsenal of hymenopterans. PLoS ONE 2014, 9, e87556. [Google Scholar]
- Touchard, A.; Dauvois, M.; Arguel, M.J.; Petitclerc, F.; Leblanc, M.; Dejean, A.; Orivel, J.; Nicholson, G.M.; Escoubas, P. Elucidation of the unexplored biodiversity of ant venom peptidomes via MALDI-TOF mass spectrometry and its application for chemotaxonomy. J. Proteom. 2014, 105, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Xing, R.; Liu, S.; Qing, Y.; Li, K.; Li, B.; Meng, X.; Cui, J.; Li, P. Isolation and in vitro partial characterization of hemolytic proteins from the nematocyst venom of the jellyfish Stomolophus meleagris. Toxicol. Vitro 2013, 27, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Moreira, D.; Chaim, O.M.; Sade, Y.B.; Paludo, K.S.; Gremski, L.H.; Donatti, L.; de Moura, J.; Mangili, O.C.; Gremski, W.; da Silveira, R.B.; et al. Identification of a direct hemolytic effect dependent on the catalytic activity induced by phospholipase-D (dermonecrotic toxin) from brown spider venom. J. Cell. Biochem. 2009, 107, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Borchani, L.; Sassi, A.; Shahbazzadeh, D.; Strub, J.M.; Tounsi-Guetteti, H.; Boubaker, M.S.; Akbari, A.; van Dorsselaer, A.; El Ayeb, M. Heminecrolysin, the first hemolytic dermonecrotic toxin purified from scorpion venom. Toxicon 2011, 58, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Biardi, J.E.; Coss, R.G. Rock squirrel (Spermophilus variegatus) blood sera affects proteolytic and hemolytic activities of rattlesnake venoms. Toxicon 2011, 57, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Matuszek, M.A.; Hodgson, W.C.; Sutherland, S.K.; King, R.G. Pharmacological studies of jumper ant (Myrmecia pilosula) venom: Evidence for the presence of histamine, and haemolytic and eicosanoid-releasing factors. Toxicon 1992, 30, 1081–1091. [Google Scholar] [CrossRef]
- Pluzhnikov, K.; Nosyreva, E.; Shevchenko, L.; Kokoz, Y.; Schmalz, D.; Hucho, F.; Grishin, E. Analysis of ectatomin action on cell membranes. Eur. J. Biochem. 1999, 262, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, A.W.; Avigad, L.S.; Schmidt, J.O.; Ishay, J.S. Proteins in venoms of two wasps, Polistes comanchus navajoe and Wasp orientalis. Comp. Biochem. Physiol. C 1982, 71, 203–207. [Google Scholar] [CrossRef]
- Hink, W.F.; Pappas, P.W.; Jaworski, D.C. Partial biochemical characterization of venom from the ant, Pseudomyrmex triplarinus. Toxicon 1994, 32, 763–772. [Google Scholar] [CrossRef]
- Tambourgi, D.V.; Morgan, B.P.; de Andrade, R.M.; Magnoli, F.C.; van Den Berg, C.W. Loxosceles intermedia spider envenomation induces activation of an endogenous metalloproteinase, resulting in cleavage of glycophorins from the erythrocyte surface and facilitating complement-mediated lysis. Blood 2000, 95, 683–691. [Google Scholar] [PubMed]
- Lam, P.D.; Mandal, P.K.; Hak, S.Y.; Hwang, S.G. Study of the molecular mechanism of anti-inflammatory activity of bee venom in lipopolysaccharide stimulated RAW 264.7 macrophages. Trop. J. Pharm. Res. 2010, 10, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Tambourgi, D.V.; Petricevich, V.L.; Magnoli, F.C.; Assaf, S.L.; Jancar, S.; Dias Da Silva, W. Endotoxemic-like shock induced by Loxosceles spider venoms: Pathological changes and putative cytokine mediators. Toxicon 1998, 36, 391–403. [Google Scholar] [CrossRef]
- Malaque, C.M.S.; Ori, M.; Santos, S.A.; Andrade, D.H. Production of TNF-α by primary cultures of human keratinocytes challenged with Loxosceles gaucho venom. Rev. Inst. Med. Trop. Sao Paulo 1999, 41, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Barbaro, K.C.; Lira, M.S.; Araújo, C.A.; Pareja-Santos, A.; Távora, B.C.; Prezotto-Neto, J.P.; Kimura, L.F.; Lima, C.; Lopes-Ferreira, M.; Santoro, M.L. Inflammatory mediators generated at the site of inoculation of Loxosceles gaucho spider venom. Toxicon 2010, 56, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Zoccal, K.F.; Bitencourt, C.S.; Secatto, A.; Sorgi, C.A.; Bordon, K.C.; Sampaio, S.V.; Arantes, E.C.; Faccioli, L.H. Tityus serrulatus venom and toxins Ts1, Ts2 and Ts6 induce macrophage activation and production of immune mediators. Toxicon 2011, 57, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.W.; O’Garra, A.; de Waal Malefyt, R.; Vieira, P.; Mosmann, T.R. Interleukin-10. Annu. Rev. Immunol. 1993, 11, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.V.; Lisboa, F.A.; Portugal, A.L.; Arantes, R.M.; Cunha-Melo, J.R. Scorpion venom increases mRNA expression of lung cytokines. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.B.; Dos Santos, J.C.; Silva, J.M.; de Godoi, P.H.; Magalhães, M.R.; Spadafora-Ferreira, M.; Fonseca, S.G.; Pfrimer, I.A. Crotalus durissus collilineatus venom induces TNF-α and IL-10 production in human peripheral blood mononuclear cells. ISRN Inflamm. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L.; Teixeira, C.F.; Tambourgi, D.V.; Gutiérrez, J.M. Increments in serum cytokine and nitric oxide levels in mice injected with Bothrops asper and Bothrops jararaca snake venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Protein/Source | Accession Code | Mw (Da) | pI | Coverage/Score |
|---|---|---|---|---|---|
| 1 | Myosin heavy chain, muscle-like isoform 2/Bombus terrestris (bee) | gi|340718034 | 251,060 | 5.58 | 15%/1509 |
| 2 | Endoplasmin/Harpegnathos saltator (ant) | gi|307192149 | 90,928 | 4.96 | 22%/907 |
| 3 | Transitional endoplasmic reticulum ATPase TER94/Camponotus floridanus (ant) | gi|307174120 | 89,511 | 5.14 | 37%/1213 |
| 4 | Hemocyte protein-glutamine gamma-glutamyltransferase/Harpegnathos saltator (ant) | gi|307215415 | 79,590 | 4.89 | 36%/1291 |
| 5 | Heat shock protein HSP 90-α/Camponotus floridanus (ant) | gi|307186382 | 83,640 | 4.98 | 40%/1410 |
| 6 | Heat shock 70 kDa protein cognate 3/Harpegnathos saltator (ant) | gi|307210158 | 72,646 | 5.02 | 44%/1709 |
| 7 | Transferrin/Harpegnathos saltator (ant) | gi|307215135 | 82,014 | 5.72 | 15%/617 |
| 8 | Transferrin/Harpegnathos saltator (ant) | gi|307215135 | 82,014 | 5.72 | 17%/718 |
| 9 | Vacuolar ATP synthase catalytic subunit A/Harpegnathos saltator (ant) | gi|307196686 | 68,591 | 5.25 | 60%/2338 |
| 10 | Heat shock protein cognate 4 (70 kDa)/Apis mellifera (bee) | gi|229892210 | 71,383 | 5.43 | 50%/2204 |
| 11 | Calreticulin/Harpegnathos saltator (ant) | gi|307191961 | 47,000 | 4.42 | 38%/810 |
| 12 | Tubulin β-1 chain/Harpegnathos saltator (ant) | gi|307201205 | 49,010 | 4.67 | 40%/818 |
| 13 | 60 kDa heat shock protein, mitochondrial/Harpegnathos saltator (ant) | gi|307199045 | 64,756 | 5.32 | 35%/1012 |
| 14 | Tubulin α-1 chain-like/Nasonia vitripennis (wasp) | gi|156548149 | 50,647 | 5.01 | 56%/1598 |
| 15 | Tubulin α-1 chain/Harpegnathos saltator (ant) | gi|307208702 | 52,080 | 5.00 | 51%/1453 |
| 16 | Putative α-tubulin/Maconellicoccus hirsutus (mealybug) | gi|121544009 | 50,633 | 5.01 | 25%/451 |
| 17 | Phosphoglucomutase/Acromyrmex echinatior (ant) | gi|332024861 | 66,241 | 6.20 | 12%/287 |
| 18 | Protein disulfide-isomerase A3/Harpegnathos saltator (ant) | gi|307194521 | 56,272 | 5.53 | 11%/260 |
| 19 | Vacuolar ATP synthase subunit B/Camponotus floridanus (ant) | gi|307174076 | 55,263 | 5.37 | 58%/1628 |
| 20 | Eukaryotic initiation factor 4A-II/Camponotus floridanus (ant) | gi|307189936 | 48,514 | 5.30 | 47%/1002 |
| 21 | ATP synthase subunit β, mitochondrial/Harpegnathos saltator (ant) | gi|307195440 | 55,223 | 5.32 | 64%/2075 |
| 22 | Tubulin β-1 chain/Apis mellifera (bee) | gi|48095525 | 50,599 | 4.75 | 65%/1193 |
| 23 | α-N-acetylgalactosaminidase/Harpegnathos saltator (ant) | gi|307213390 | 161,528 | 5.99 | 6%/368 |
| 24 | cAMP-dependent protein kinase type I regulatory subunit isoform 1/Apis mellifera (bee) | gi|48106841 | 41,923 | 4.86 | 34%/611 |
| 27 | Lysosomal aspartic protease/Acromyrmex echinatior (ant) | gi|332024025 | 42,132 | 5.34 | 13%/254 |
| 28 | Actin-5, muscle-specific/Harpegnathos saltator (ant) | gi|307197034 | 42,098 | 5.30 | 68%/1145 |
| 29 | Actin, muscle/Camponotus floridanus (ant) | gi|307166491 | 42,071 | 5.29 | 71%/1252 |
| 30 | Actin, muscle/Camponotus floridanus (ant) | gi|307166491 | 42,071 | 5.29 | 62%/955 |
| 31 | Actin/Aedes aegypti (mosquito) | gi|94468486 | 42,164 | 5.30 | 40%/484 |
| 32 | Rab GDP dissociation inhibitor β/Camponotus floridanus (ant) | gi|307172388 | 50,104 | 5.51 | 36%/757 |
| 33 | Arginine kinase/Acromyrmex echinatior (ant) | gi|332018357 | 40,032 | 5.86 | 18%/313 |
| 34 | Mitochondrial-processing peptidase subunit β/Harpegnathos saltator (ant) | gi|307207091 | 53,918 | 5.77 | 28%/572 |
| 35 | Enolase/Harpegnathos saltator (ant) | gi|307211488 | 47,379 | 5.79 | 40%/650 |
| 36 | Succinyl-CoA ligase (ADP-forming) subunit β, mitochondrial-like/Bombus terrestris (bee) | gi|340726331 | 49,012 | 6.63 | 31%/861 |
| 37 | Arginine kinase/Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 45%/1201 |
| 38 | RecName: Full = Arginine kinase; Short = AK/Homarus gammarus (lobster) | gi|585342 | 40,300 | 6.05 | 4%/88 |
| 39 | Retinal dehydrogenase 1/Acromyrmex echinatior (ant) | gi|332024132 | 53,459 | 6.14 | 9%/283 |
| 40 | Retinal dehydrogenase 1/Acromyrmex echinatior (ant) | gi|332024132 | 53,459 | 6.14 | 9%/333 |
| 41 | α-Tubulin at 84B/Drosophila melanogaster (fly) | gi|17136564 | 50,561 | 5.00 | 25%/402 |
| 42 | Retinal dehydrogenase 1/Acromyrmex echinatior (ant) | gi|332024132 | 53,459 | 6.14 | 9%/333 |
| 43 | Aminoacylase-1/Harpegnathos saltator (ant) | gi|307206409 | 45,783 | 5.66 | 16%/319 |
| 44 | Adenosylhomocysteinase/Harpegnathos saltator (ant) | gi|307206413 | 48,440 | 5.88 | 30%/737 |
| 45 | Enolase/Harpegnathos saltator (ant) | gi|307211488 | 47,379 | 5.79 | 44%/778 |
| 46 | Enolase/Harpegnathos saltator (ant) | gi|307211488 | 47,379 | 5.79 | 16%/307 |
| 47 | Phosphoglycerate kinase/Camponotus floridanus (ant) | gi|307177429 | 45,039 | 6.16 | 35%/1031 |
| 48 | Phosphoglycerate kinase/Camponotus floridanus (ant) | gi|307177429 | 45,039 | 6.16 | 23%/416 |
| 49 | Arginine kinase/Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 47%/1381 |
| 50 | Arginine kinase/Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 41%/1045 |
| 51 | Arginine kinase/Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 45%/637 |
| 52 | Arginine kinase/Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 39%/652 |
| 53 | Arginine kinase/Acromyrmex echinatior (ant) | gi|332018357 | 40,032 | 5.86 | 10%/211 |
| 54 | Malate dehydrogenase, cytoplasmic/Camponotus floridanus (ant) | gi|307166391 | 39,169 | 7.03 | 20%/533 |
| 55 | Glycerol-3-phosphate dehydrogenase (NAD+), cytoplasmic/Acromyrmex echinatior (ant) | gi|332024225 | 37,294 | 7.15 | 44%/1106 |
| 56 | Glycerol-3-phosphate dehydrogenase (NAD+), cytoplasmic/Acromyrmex echinatior (ant) | gi|332024225 | 37,294 | 7.15 | 25%/290 |
| 57 | Superoxide dismutase (Cu-Zn)/Harpegnathos saltator (ant) | gi|307204104 | 14,008 | 6.18 | 43%/213 |
| 58 | Glycogen phosphorylase/Harpegnathos saltator (ant) | gi|307199215 | 121,697 | 6.22 | 17%/837 |
| 59 | Glycogen phosphorylase/Harpegnathos saltator (ant) | gi|307199215 | 121,697 | 6.22 | 20%/1060 |
| 60 | Cytoplasmic aconitate hydratase/Harpegnathos saltator (ant) | gi|307196718 | 98,222 | 6.11 | 11%/319 |
| 61 | 10-formyltetrahydrofolate dehydrogenase/Acromyrmex echinatior (ant) | gi|332029989 | 99,956 | 5.88 | 4%/139 |
| 62 | Elongation factor 2/Camponotus floridanus (ant) | gi|307170298 | 94,303 | 6.11 | 16%/496 |
| 63 | 2-oxoglutarate dehydrogenase E1 component, mitochondrial/Acromyrmex echinatior (ant) | gi|332017156 | 121,347 | 6.78 | 13%/541 |
| 64 | 2-oxoglutarate dehydrogenase E1 component, mitochondrial/Acromyrmex echinatior (ant) | gi|332017156 | 121,347 | 6.78 | 25%/1252 |
| 65 | 2-oxoglutarate dehydrogenase E1 component, mitochondrial/Acromyrmex echinatior (ant) | gi|332017156 | 121,347 | 6.78 | 26%/1262 |
| 66 | Filamin-C/Harpegnathos saltator (ant) | gi|307210403 | 252,199 | 6.12 | 14%/1364 |
| 67 | Filamin-C/Harpegnathos saltator (ant) | gi|307210403 | 252,199 | 6.12 | 17%/1704 |
| 68 | Filamin-C/Harpegnathos saltator (ant) | gi|307210403 | 252,199 | 6.12 | 18%/2261 |
| 69 | Filamin-C/Harpegnathos saltator (ant) | gi|307210403 | 252,199 | 6.12 | 18%/1008 |
| 70 | Aconitate hydratase, mitochondrial/Harpegnathos saltator (ant) | gi|307201595 | 85028 | 8.36 | 25%/839 |
| 71 | Aconitate hydratase, mitochondrial/Harpegnathos saltator (ant) | gi|307201595 | 85028 | 8.36 | 36%/1333 |
| 72 | Succinate dehydrogenase (ubiquinone) flavoprotein subunit, mitochondrial/Acromyrmex echinatior (ant) | gi|332019677 | 73506 | 6.75 | 27%/886 |
| 73 | Putative actin-interacting protein 1/Harpegnathos saltator (ant) | gi|307210939 | 69910 | 5.69 | 36%/1180 |
| 74 | Putative actin-interacting protein 1/Harpegnathos saltator (ant) | gi|307210939 | 69910 | 5.69 | 29%/708 |
| 75 | Glycerol-3-phosphate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307212068 | 81972 | 6.51 | 16%/530 |
| 76 | Glycerol-3-phosphate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307212068 | 81972 | 6.51 | 13%/292 |
| 77 | Apolipophorins/Harpegnathos saltator (ant) | gi|307201472 | 382,345 | 6.47 | 1%/400 |
| 78 | NADP-dependent malic enzyme/Harpegnathos saltator (ant) | gi|307205633 | 66,782 | 7.19 | 29%/763 |
| 79 | NADP-dependent malic enzyme/Harpegnathos saltator (ant) | gi|307205633 | 66,782 | 7.19 | 13%/207 |
| 80 | NADP-dependent malic enzyme/Harpegnathos saltator (ant) | gi|307205633 | 66,782 | 7.19 | 49%/1238 |
| 81 | Dihydrolipoyl dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307209020 | 54,499 | 6.87 | 7%/117 |
| 82 | NADP-dependent malic enzyme/Harpegnathos saltator (ant) | gi|307205633 | 66,782 | 7.19 | 54%/1447 |
| 83 | NADP-dependent malic enzyme/Harpegnathos saltator (ant) | gi|307205633 | 66,782 | 7.19 | 12%/195 |
| 84 | Transaldolase/Harpegnathos saltator (ant) | gi|307215256 | 37,393 | 5.27 | 7%/102 |
| 85 | Pyruvate dehydrogenase E1 component subunit β, mitochondrial/Harpegnathos saltator (ant) | gi|307195718 | 39,486 | 5.87 | 34%/475 |
| 86 | Proteasome subunit β type-7/Harpegnathos saltator (ant) | gi|307192825 | 30,622 | 7.00 | 28%/250 |
| 87 | Peroxiredoxin 1/Camponotus floridanus (ant) | gi|307175821 | 21,887 | 6.30 | 32%/423 |
| 88 | Proteasome subunit β type-1/Harpegnathos saltator (ant) | gi|307214019 | 26,343 | 6.81 | 20%/108 |
| 89 | Dihydrolipoyl dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307209020 | 54,499 | 6.87 | 23%/491 |
| 90 | Glutamate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307195623 | 62,095 | 8.40 | 27%/549 |
| 91 | Selenium-binding protein 1-A/Acromyrmex echinatior (ant) | gi|332021867 | 52,719 | 6.99 | 8%/233 |
| 92 | Glutamate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307195623 | 62,095 | 8.40 | 52%/1140 |
| 93 | Glutamate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307195623 | 62,095 | 8.40 | 53%/1166 |
| 94 | Glutamate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307195623 | 62,095 | 8.40 | 52%/1356 |
| 95 | Catalase/Harpegnathos saltator (ant) | gi|307197480 | 58,234 | 8.58 | 23%/170 |
| 96 | Elongation factor-1 α, partial/Hoplitis ochraceicornis (bee) | gi|426322152 | 40,701 | 8.58 | 41%/226 |
| 97 | Alanine aminotransferase 2/Harpegnathos saltator (ant) | gi|307214462 | 61,520 | 9.26 | 24%/665 |
| 98 | ATP synthase subunit α, mitochondrial/Harpegnathos saltator (ant) | gi|307208992 | 59,687 | 9.12 | 41%/1271 |
| 99 | Elongation factor 1-α/Harpegnathos saltator (ant) | gi|307196337 | 50,620 | 9.13 | 43%/350 |
| 100 | ATP synthase subunit α, mitochondrial/Harpegnathos saltator (ant) | gi|307208992 | 59,687 | 9.12 | 32%/364 |
| 101 | 4-hydroxybutyrate coenzyme A transferase/Harpegnathos saltator (ant) | gi|307203837 | 53,231 | 7.69 | 36%/769 |
| 102 | Probable fumarate hydratase, mitochondrial/Harpegnathos saltator (ant) | gi|307202128 | 47,153 | 8.14 | 24%/478 |
| 103 | Probable citrate synthase 1, mitochondrial/Harpegnathos saltator (ant) | gi|307202019 | 49,316 | 8.94 | 34%/788 |
| 104 | Probable citrate synthase 1, mitochondrial/Harpegnathos saltator (ant) | gi|307202019 | 49,316 | 8.94 | 37%/471 |
| 105 | Isocitrate dehydrogenase (NADP) cytoplasmic-like isoform 1/Bombus terrestris (bee) | gi|340721268 | 46,518 | 8.00 | 9%/146 |
| 106 | Isocitrate dehydrogenase (NADP) cytoplasmic/Harpegnathos saltator (ant) | gi|307210166 | 50,629 | 8.20 | 44%/952 |
| 107 | Isocitrate dehydrogenase (NADP) cytoplasmic/Harpegnathos saltator (ant) | gi|307210166 | 50,629 | 8.20 | 41%/611 |
| 108 | Isocitrate dehydrogenase (NADP) cytoplasmic/Harpegnathos saltator (ant) | gi|307210166 | 50,629 | 8.20 | 19%/422 |
| 109 | Arginine kinase /Harpegnathos saltator (ant) | gi|307197996 | 39,996 | 5.75 | 46%/1213 |
| 110 | Fructose-bisphosphate aldolase-like/Megachile rotundata (bee) | gi|383861513 | 40,009 | 7.00 | 18%/470 |
| 111 | Four and a half LIM domains protein 2/Acromyrmex echinatior (ant) | gi|332021158 | 28,335 | 8.46 | 52%/758 |
| 112 | Isocitrate dehydrogenase (NAD) subunit β, mitochondrial/Harpegnathos saltator (ant) | gi|307210578 | 41,512 | 8.81 | 31%/530 |
| 113 | Sorbitol dehydrogenase/Harpegnathos saltator (ant) | gi|307204829 | 38,256 | 7.56 | 27%/488 |
| 114 | Fructose-bisphosphate aldolase-like/Megachile rotundata (bee) | gi|383861513 | 40,009 | 7.00 | 33%/635 |
| 115 | Aldose reductase/Camponotus floridanus (ant) | gi|307181859 | 35,897 | 6.11 | 17%/212 |
| 116 | Glyceraldehyde-3-phosphate dehydrogenase 2/Harpegnathos saltator (ant) | gi|307198667 | 35,769 | 7.64 | 45%/898 |
| 117 | Glyceraldehyde-3-phosphate dehydrogenase 2/Harpegnathos saltator (ant) | gi|307198667 | 35,769 | 7.64 | 45%/848 |
| 118 | Glyceraldehyde-3-phosphate dehydrogenase 2/Harpegnathos saltator (ant) | gi|307198667 | 35,769 | 7.64 | 42%/603 |
| 119 | Hyaluronoglucosaminidase/Camponotus floridanus (ant) | gi|307180582 | 40,178 | 8.45 | 10%/216 |
| 120 | Succinyl-CoA ligase (GDP-forming) subunit α, mitochondrial/Harpegnathos saltator (ant) | gi|307203732 | 34,525 | 8.70 | 23%/476 |
| 121 | Guanine nucleotide-binding protein subunit β-like protein/Harpegnathos saltator (ant) | gi|307215409 | 36,333 | 7.63 | 68%/913 |
| 122 | Malate dehydrogenase, mitochondrial/Harpegnathos saltator (ant) | gi|307214026 | 36,178 | 9.36 | 46%/670 |
| 123 | Phospholipase A2 isozymes PA3A/PA3B/PA5/Apis mellifera (bee) | gi|328778177 | 26,379 | 7.46 | 6%/103 |
| 124 | Proteasome subunit α type-7-1/Harpegnathos saltator (ant) | gi|307201582 | 28,118 | 6.97 | 32%/385 |
| 125 | Dehydrogenase/reductase SDR family member 11/Harpegnathos saltator (ant) | gi|307212910 | 17,798 | 9.61 | 9%/102 |
| 128 | Phosphoglycerate mutase 2-like/Bombus terrestris (bee) | gi|340726229 | 35,071 | 9.16 | 36%/733 |
| 129 | Glutathione S-transferase sigma 3/Locusta migratoria (locust) | gi|329564873 | 23,498 | 7.63 | 8%/94 |
| 130 | Adenylate kinase isoenzyme F38B2.4/Harpegnathos saltator (ant) | gi|307204436 | 21,291 | 5.42 | 35%/525 |
| 131 | Protein DJ-1/Camponotus floridanus (ant) | gi|307174129 | 20,023 | 5.93 | 7%/110 |
| 132 | RecName: Full = Snake venom metalloproteinase atroxlysin-1; Short = SVMP; AltName: Full = Atroxlysin-I/Bothrops atrox (snake) | gi|353526296 | 23,302 | 5.85 | 12%/135 |
| 134 | Transaldolase/Harpegnathos saltator (ant) | gi|307215256 | 37,393 | 5.27 | 7%/168 |
| 135 | Peroxiredoxin-6/Harpegnathos saltator (ant) | gi|307213913 | 25,248 | 6.65 | 18%/124 |
| 136 | Triosephosphate isomerase/Harpegnathos saltator (ant) | gi|307199049 | 26,956 | 6.00 | 49%/695 |
| 137 | Eukaryotic translation initiation factor 3 subunit I/Acromyrmex echinatior (ant) | gi|332024413 | 36,384 | 6.00 | 25%/210 |
| 138 | Glycine N-methyltransferase/Harpegnathos saltator (ant) | gi|307192516 | 33,820 | 6.26 | 19%/259 |
| 139 | Fumarylacetoacetate hydrolase domain-containing protein 2A/Harpegnathos saltator (ant) | gi|307203489 | 37,311 | 8.54 | 8%/109 |
| 140 | Ribose-phosphate pyrophosphokinase 1/Harpegnathos saltator (ant) | gi|307207320 | 37,856 | 6.26 | 23%/320 |
| 142 | 14-3-3 protein epsilon-like/Nasonia vitripennis (wasp) | gi|156553657 | 29,323 | 4.72 | 4%/123 |
| 143 | Putative 14-3-3 protein/Maconellicoccus hirsutus (mealybug) | gi|121543925 | 28,171 | 4.80 | 9%/83 |
| 144 | 14-3-3 protein ζ/Camponotus floridanus (ant) | gi|307186410 | 35,115 | 5.01 | 11%/149 |
| 145 | Glutathione S-transferase 1, isoform C/Harpegnathos saltator (ant) | gi|307196173 | 24,899 | 7.71 | 18%/175 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pessoa, W.F.B.; Silva, L.C.C.; De Oliveira Dias, L.; Delabie, J.H.C.; Costa, H.; Romano, C.C. Analysis of Protein Composition and Bioactivity of Neoponera villosa Venom (Hymenoptera: Formicidae). Int. J. Mol. Sci. 2016, 17, 513. https://doi.org/10.3390/ijms17040513
Pessoa WFB, Silva LCC, De Oliveira Dias L, Delabie JHC, Costa H, Romano CC. Analysis of Protein Composition and Bioactivity of Neoponera villosa Venom (Hymenoptera: Formicidae). International Journal of Molecular Sciences. 2016; 17(4):513. https://doi.org/10.3390/ijms17040513
Chicago/Turabian StylePessoa, Wallace Felipe Blohem, Ludimilla Carvalho Cerqueira Silva, Leila De Oliveira Dias, Jacques Hubert Charles Delabie, Helena Costa, and Carla Cristina Romano. 2016. "Analysis of Protein Composition and Bioactivity of Neoponera villosa Venom (Hymenoptera: Formicidae)" International Journal of Molecular Sciences 17, no. 4: 513. https://doi.org/10.3390/ijms17040513
APA StylePessoa, W. F. B., Silva, L. C. C., De Oliveira Dias, L., Delabie, J. H. C., Costa, H., & Romano, C. C. (2016). Analysis of Protein Composition and Bioactivity of Neoponera villosa Venom (Hymenoptera: Formicidae). International Journal of Molecular Sciences, 17(4), 513. https://doi.org/10.3390/ijms17040513