
Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis

 ,
,
Abstract
:1. Introduction
2. Results
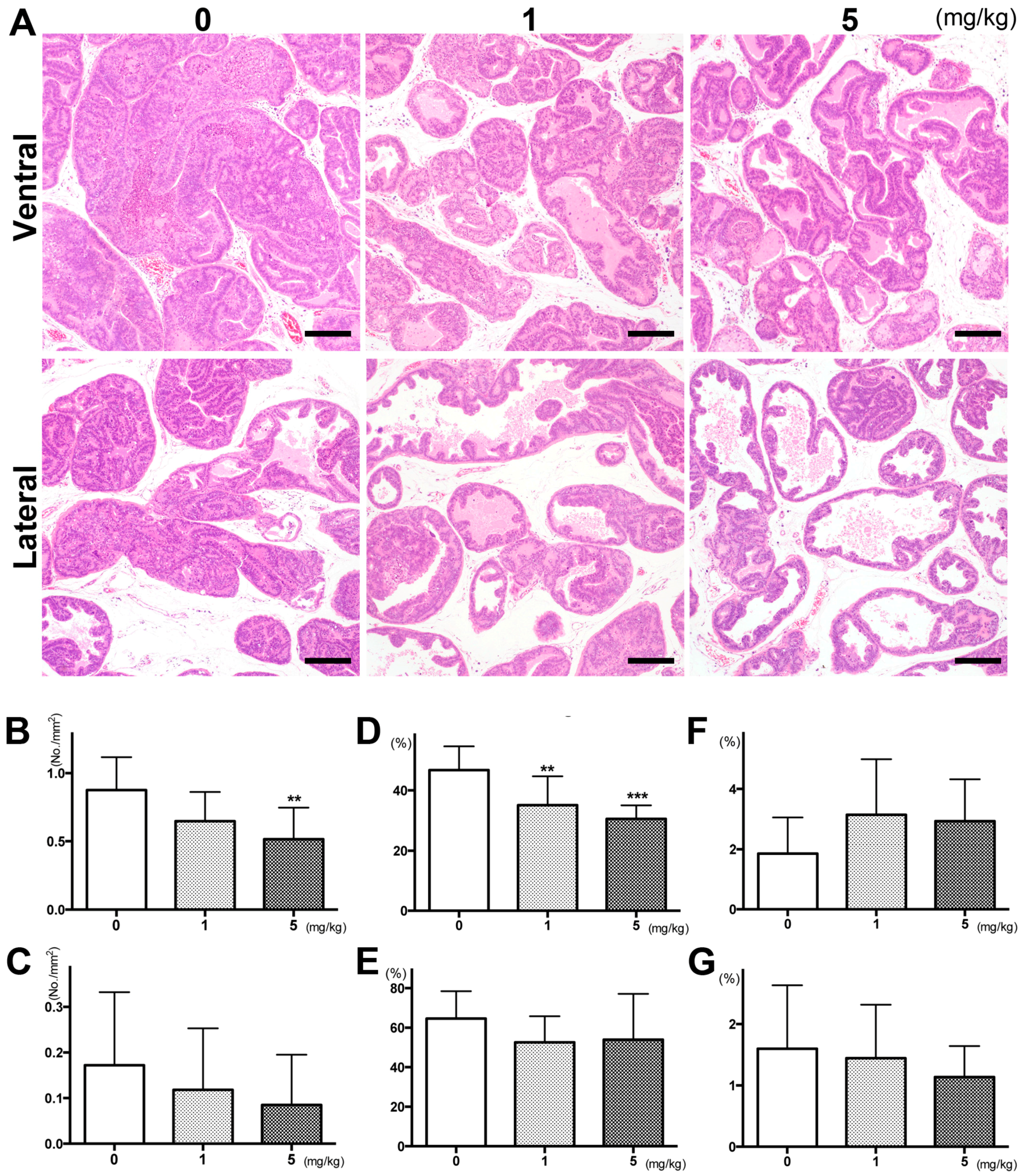
2.1. PGZ Inhibited the Progression of Prostate Tumorigenesis as Well as Cell Proliferation in TRAP Rats
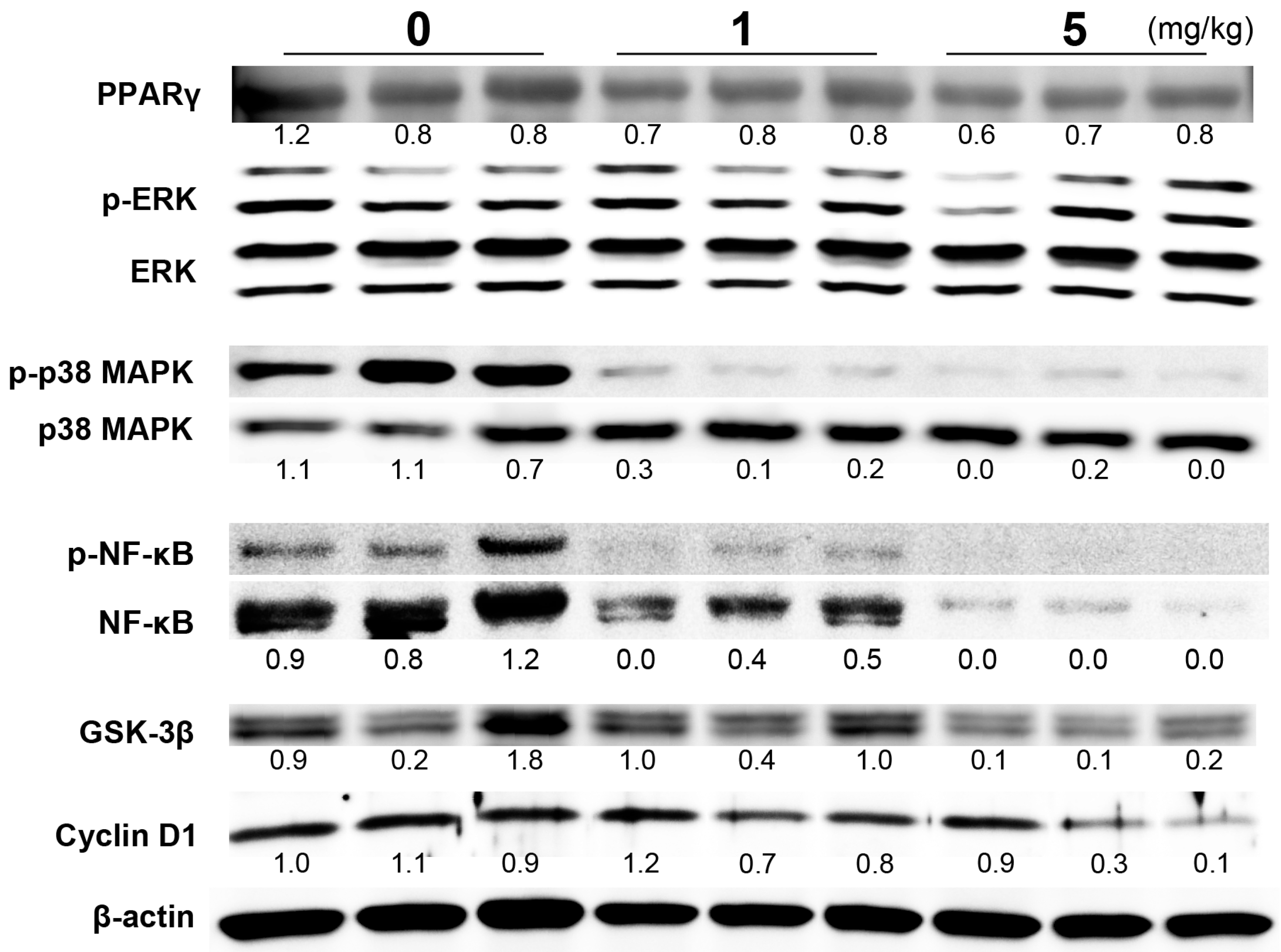
2.2. Reduction of Cell Proliferation by PGZ Treatment Was Associated with Changes in the NFκB and p38 Pathways
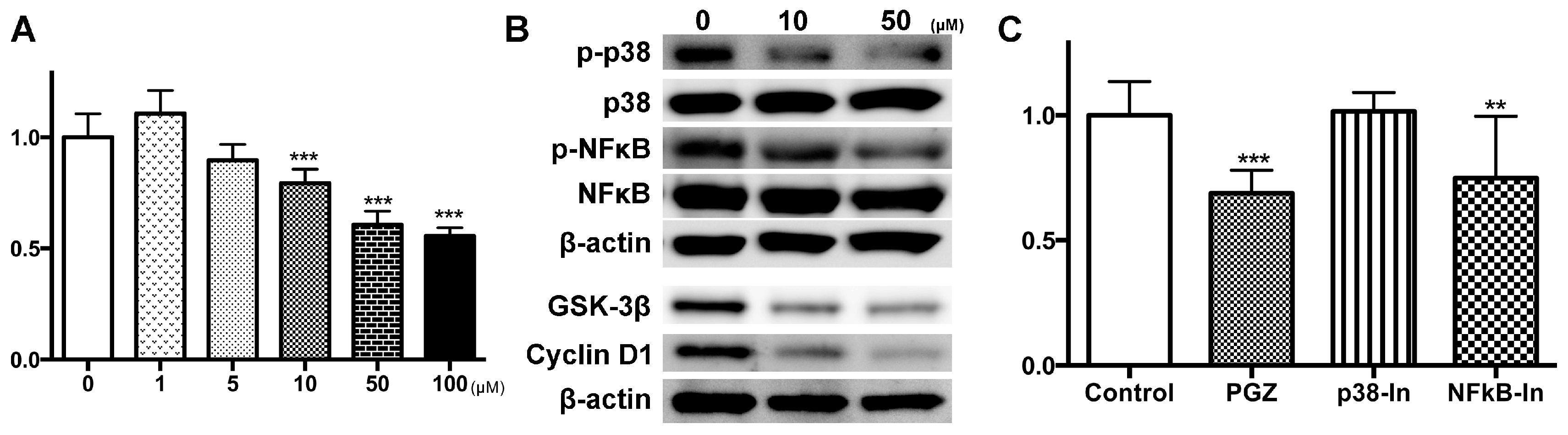
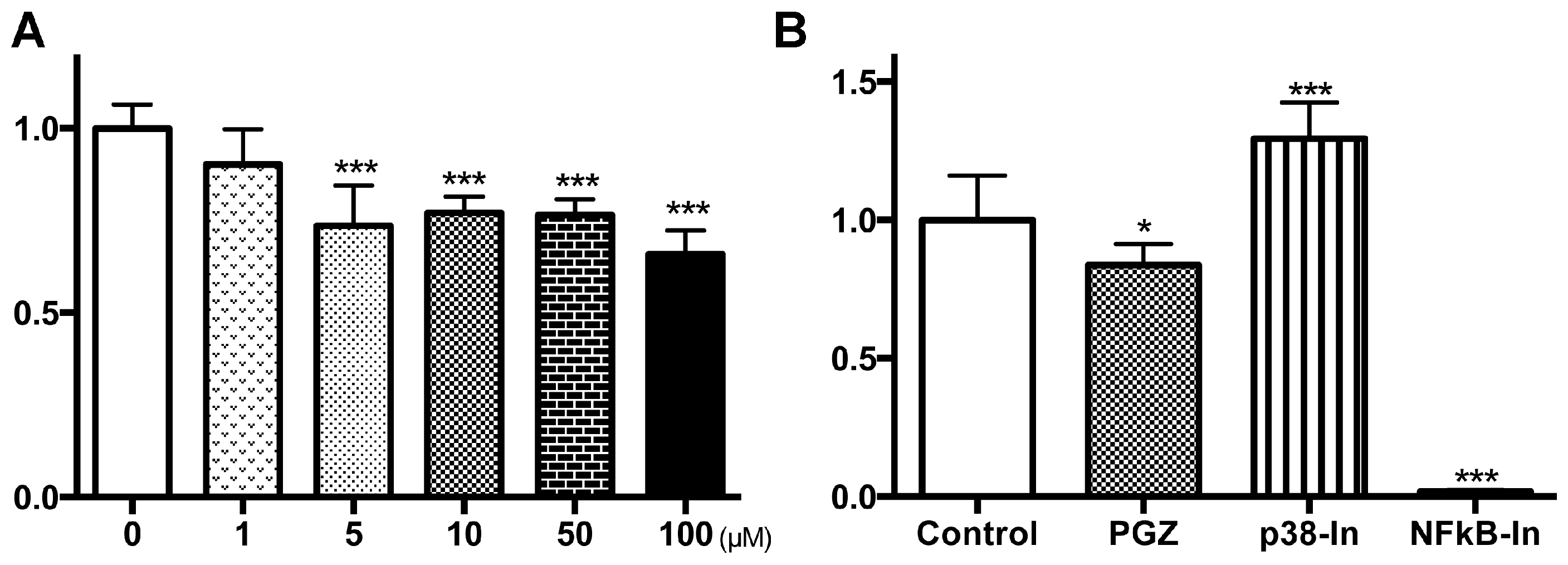
2.3. PGZ Affected the Reduction of Cell Growth in Human Prostate Cancer Cell Lines
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Histopathology, Immunohistochemistry and TUNEL Assays
4.3. Cell Lines
4.4. Cell Proliferation Assay
4.5. Immunoblot Analyses
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Torre, L.A.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global cancer incidence and mortality rates and trends—An update. Can. Epidemiol. Biomed. Prev. 2016, 25, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Hori, M.; Matsuda, T.; Shibata, A.; Katanoda, K.; Sobue, T.; Nishimoto, H. Cancer incidence and incidence rates in Japan in 2009: A study of 32 population-based cancer registries for the monitoring of cancer incidence in Japan (MCIJ) project. Jpn. J. Clin. Oncol. 2015, 45, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, G.S.; Nepple, K.G.; Tanagho, Y.S.; Andriole, G.L. Prostate cancer chemoprevention. Semin. Oncol. 2013, 40, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Vemana, G.; Hamilton, R.J.; Andriole, G.L.; Freedland, S.J. Chemoprevention of prostate cancer. Annu. Rev. Med. 2014, 65, 111–123. [Google Scholar] [PubMed]
- Tachibana, K.; Yamasaki, D.; Ishimoto, K.; Doi, T. The role of PPARs in cancer. PPAR Res. 2008, 2008, 102737. [Google Scholar] [CrossRef] [PubMed]
- Robbins, G.T.; Nie, D. PPAR γ, bioactive lipids, and cancer progression. Front. Biosci. 2012, 17, 1816–1834. [Google Scholar] [CrossRef]
- Yin, F.; Bruemmer, D.; Blaschke, F.; Hsueh, W.A.; Law, R.E.; Herle, A.J. Signaling pathways involved in induction of Gadd45 gene expression and apoptosis by troglitazone in human MCF-7 breast carcinoma cells. Oncogene 2004, 23, 4614–4623. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.O.; Oh, J.H.; Son, S.M.; Won, D.; Song, H.S.; Han, S.B.; Moon, D.C.; Kang, K.W.; Song, M.J.; Hong, J.T. Troglitazone, a PPAR agonist, inhibits human prostate cancer cell growth through inactivation of NFκB via suppression of Gsk-3β expression. Cancer Biol. Ther. 2011, 12, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kohno, H.; Yoshitani, S.; Takashima, S.; Okumura, A.; Murakami, A.; Hosokawa, M. Ligands for peroxisome proliferator-activated receptors α and γ inhibit chemically induced colitis and formation of aberrant crypt foci in rats. Cancer Res. 2001, 61, 2424–2428. [Google Scholar] [PubMed]
- Yoshida, K.; Hirose, Y.; Tanaka, T.; Yamada, Y.; Kuno, T.; Kohno, H.; Katayama, M.; Qiao, Z.; Sakata, K.; Sugie, S.; et al. Inhibitory effects of troglitazone, a peroxisome proliferator-activated receptor γ ligand, in rat tongue carcinogenesis initiated with 4-nitroquinoline 1-oxide. Cancer Sci. 2003, 94, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.T.; Leng, X.S.; Li, T.; Zhao, J.M.; Lin, X.H. Peroxisome proliferator-activated receptor γ ligands suppress liver carcinogenesis induced by diethylnitrosamine in rats. World J. Gastroenterol. 2004, 10, 3419–3423. [Google Scholar] [CrossRef] [PubMed]
- Nicol, C.J.; Yoon, M.; Ward, J.M.; Yamashita, M.; Fukamachi, K.; Peters, J.M.; Gonzalez, F.J. PPARγ influences susceptibility to DMBA-induced mammary, ovarian and skin carcinogenesis. Carcinogenesis 2004, 25, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.M. Effects of PPARγ and combined agonists on the urinary tract of rats and other species. Toxicol. Sci. 2005, 87, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Asamoto, M.; Hokaiwado, N.; Cho, Y.M.; Takahashi, S.; Ikeda, Y.; Imaida, K.; Shirai, T. Prostate carcinomas developing in transgenic rats with SV40 T antigen expression under probasin promoter control are strictly androgen dependent. Cancer Res. 2001, 61, 4693–4700. [Google Scholar] [PubMed]
- Cho, Y.M.; Takahashi, S.; Asamoto, M.; Suzuki, S.; Inaguma, S.; Hokaiwado, N.; Shirai, T. Age-dependent histopathological findings in the prostate of probasin/SV40 T antigen transgenic rats: Lack of influence of carcinogen or testosterone treatment. Cancer Sci. 2003, 94, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Uemura, H.; Seeni, A.; Tang, M.; Komiya, M.; Long, N.; Ishiguro, H.; Kubota, Y.; Shirai, T. Therapeutic targeting of angiotensin II receptor type 1 to regulate androgen receptor in prostate cancer. Prostate 2012, 72, 1559–1572. [Google Scholar] [CrossRef] [PubMed]
- Long, N.; Suzuki, S.; Sato, S.; Naiki-Ito, A.; Sakatani, K.; Shirai, T.; Takahashi, S. Purple corn color inhibition of prostate carcinogenesis by targeting cell growth pathways. Cancer Sci. 2013, 104, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Shiraga, K.; Sato, S.; Punfa, W.; Naiki-Ito, A.; Yamashita, Y.; Shirai, T.; Takahashi, S. Apocynin, an NADPH oxidase inhibitor, suppresses rat prostate carcinogenesis. Cancer Sci. 2013, 104, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Berquin, I.M.; Min, Y.; Wu, R.; Wu, H.; Chen, Y.Q. Expression signature of the mouse prostate. J. Biol. Chem. 2005, 280, 36442–36451. [Google Scholar] [CrossRef] [PubMed]
- Shirai, T.; Sakata, T.; Fukushima, S.; Ikawa, E.; Ito, N. Rat prostate as one of the target organs for 3,2’-dimethyl-4-aminobiphenyl-induced carcinogenesis: Effects of dietary ethinyl estradiol and methyltestosterone. Jpn. J. Cancer Res. 1985, 76, 803–808. [Google Scholar] [PubMed]
- Shirai, T.; Sano, M.; Tamano, S.; Takahashi, S.; Hirose, M.; Futakuchi, M.; Hasegawa, R.; Imaida, K.; Matsumoto, K.; Wakabayashi, K.; et al. The prostate: A target for carcinogenicity of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (phip) derived from cooked foods. Cancer Res. 1997, 57, 195–198. [Google Scholar] [PubMed]
- Bosetti, C.; Rosato, V.; Buniato, D.; Zambon, A.; La Vecchia, C.; Corrao, G. Cancer risk for patients using thiazolidinediones for type 2 diabetes: A meta-analysis. Oncologist 2013, 18, 148–156. [Google Scholar] [CrossRef] [PubMed]
- EI-Hage, J. Peroxisome Proliferator-Activated Receptor Agonists: Carcinogenicity Findings and Regulatory Recommendations; International Athroscleosis Society Symposium on PPAR: Monte Carlo, Monaco, 2005. [Google Scholar]
- Lewis, J.D.; Ferrara, A.; Peng, T.; Hedderson, M.; Bilker, W.B.; Quesenberry, C.P., Jr.; Vaughn, D.J.; Nessel, L.; Selby, J.; Strom, B.L. Risk of bladder cancer among diabetic patients treated with pioglitazone: Interim report of a longitudinal cohort study. Diabetes Care 2011, 34, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Dominick, M.A.; White, M.R.; Sanderson, T.P.; van Vleet, T.; Cohen, S.M.; Arnold, L.E.; Cano, M.; Tannehill-Gregg, S.; Moehlenkamp, J.D.; Waites, C.R.; et al. Urothelial carcinogenesis in the urinary bladder of male rats treated with muraglitazar, a PPAR α/γ agonist: Evidence for urolithiasis as the inciting event in the mode of action. Toxicol. Pathol. 2006, 34, 903–920. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.; Bell, S.; Sund, R.; Hartikainen, S.A.; Tuomilehto, J.; Pukkala, E.; Keskimaki, I.; Badrick, E.; Renehan, A.G.; Buchan, I.E.; et al. Pioglitazone and bladder cancer risk: A multipopulation pooled, cumulative exposure analysis. Diabetologia 2015, 58, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Habel, L.A.; Quesenberry, C.P.; Strom, B.L.; Peng, T.; Hedderson, M.M.; Ehrlich, S.F.; Mamtani, R.; Bilker, W.; Vaughn, D.J.; et al. Pioglitazone use and risk of bladder cancer and other common cancers in persons with diabetes. JAMA 2015, 314, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Daniyal, M.; Siddiqui, Z.A.; Akram, M.; Asif, H.M.; Sultana, S.; Khan, A. Epidemiology, etiology, diagnosis and treatment of prostate cancer. Asian Pac. J. Cancer Prev. 2014, 15, 9575–9578. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.T.; Niu, H.S.; Chen, L.J.; Cheng, J.T.; Tong, Y.C. Increase of human prostate cancer cell (DU145) apoptosis by telmisartan through PPAR-Δ pathway. Eur. J. Pharmacol. 2016, 775, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Seeni, A.; Takahashi, S.; Takeshita, K.; Tang, M.; Sugiura, S.; Sato, S.Y.; Shirai, T. Suppression of prostate cancer growth by resveratrol in the transgenic rat for adenocarcinoma of prostate (TRAP) model. Asian Pac. J. Cancer Prev. 2008, 9, 7–14. [Google Scholar] [PubMed]
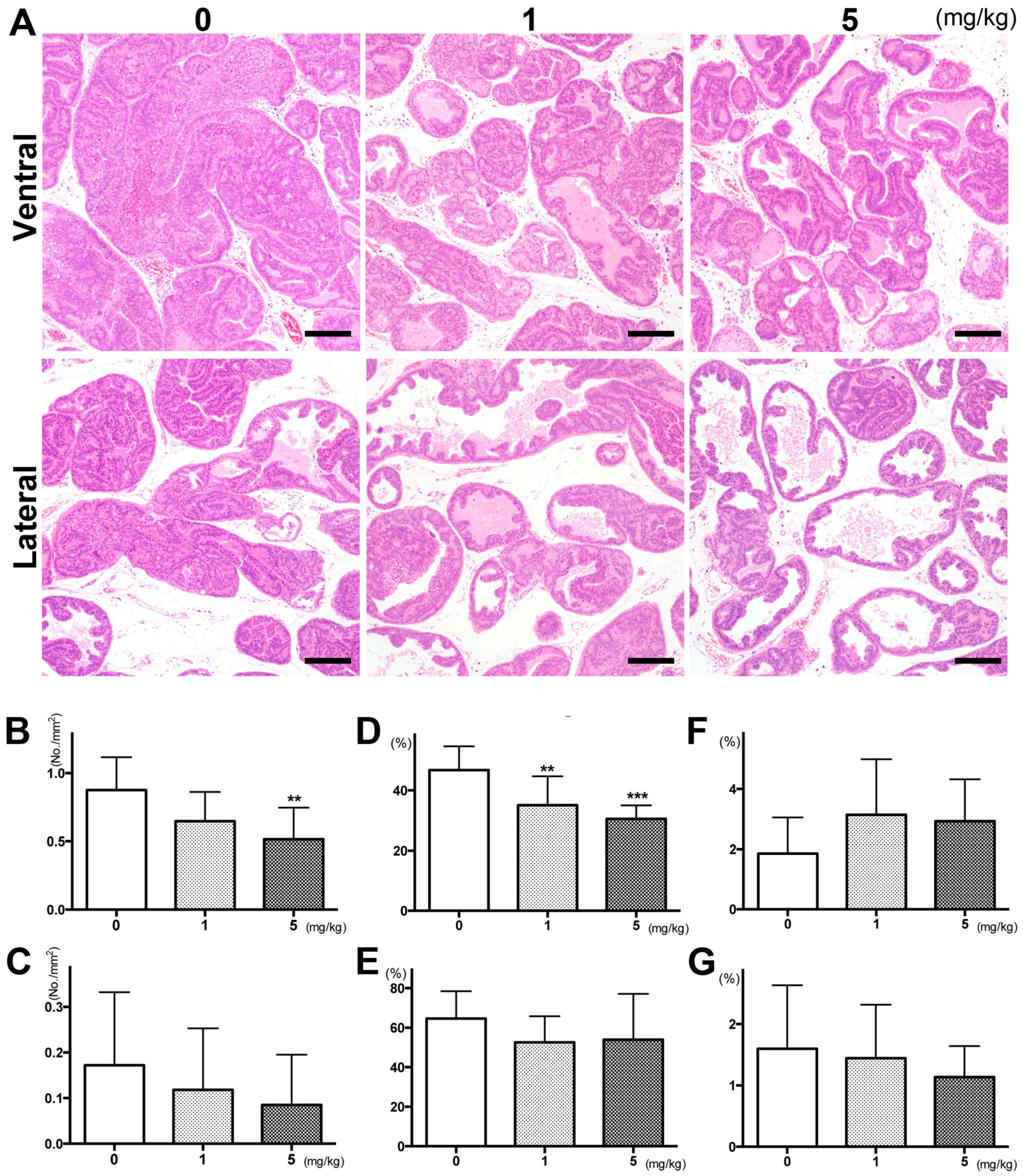
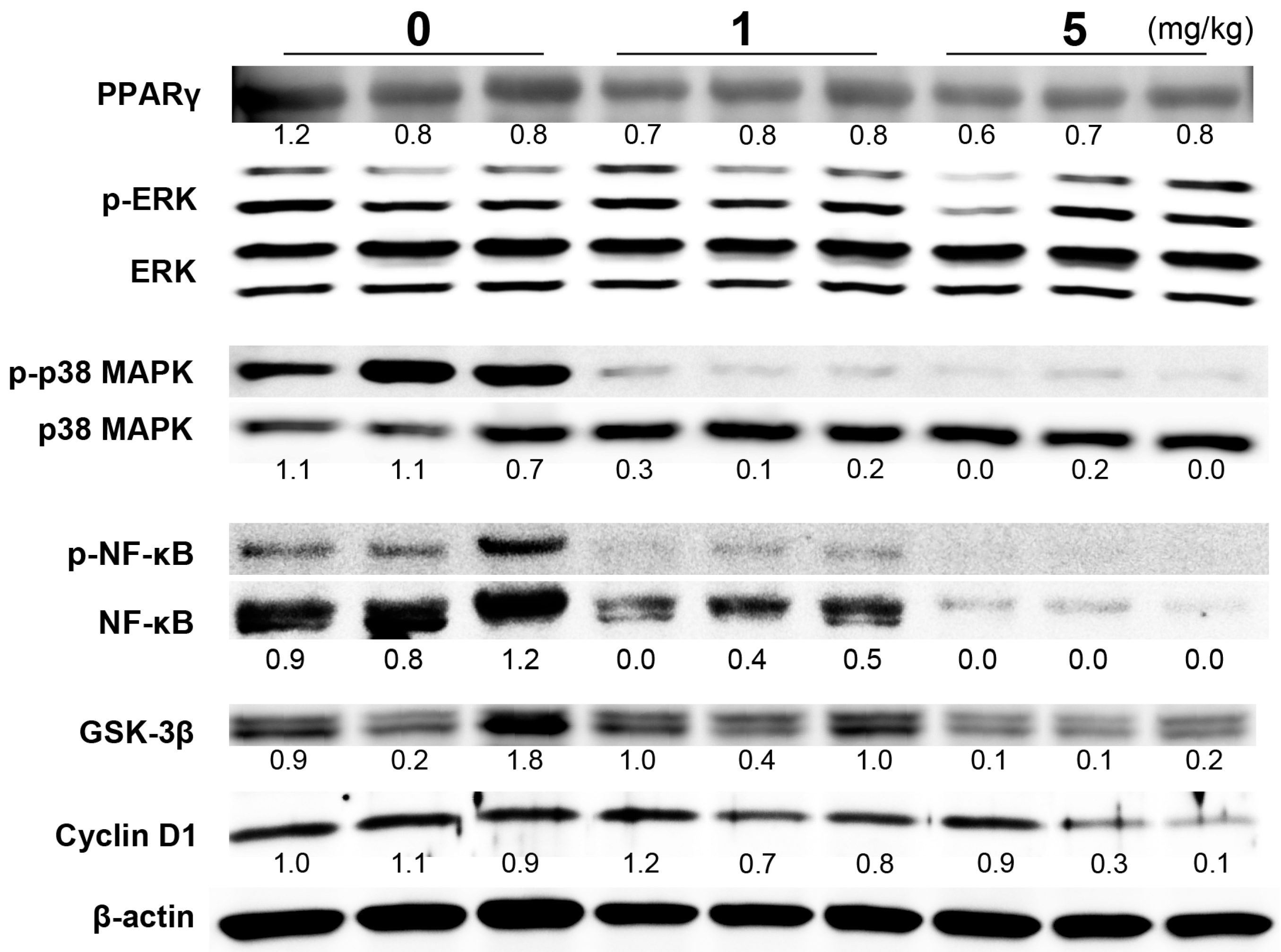
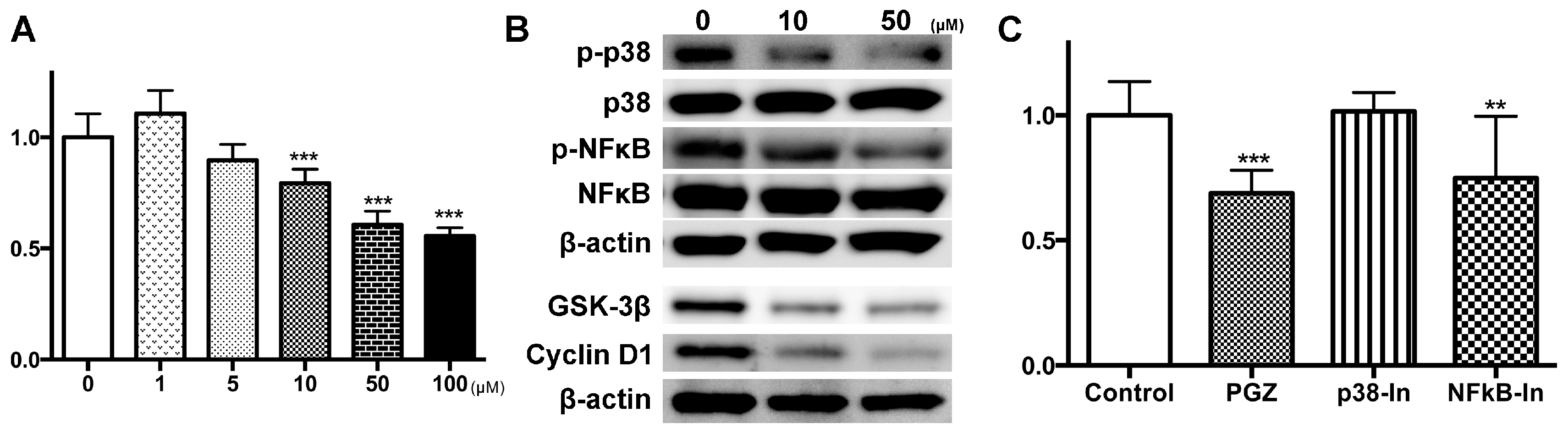
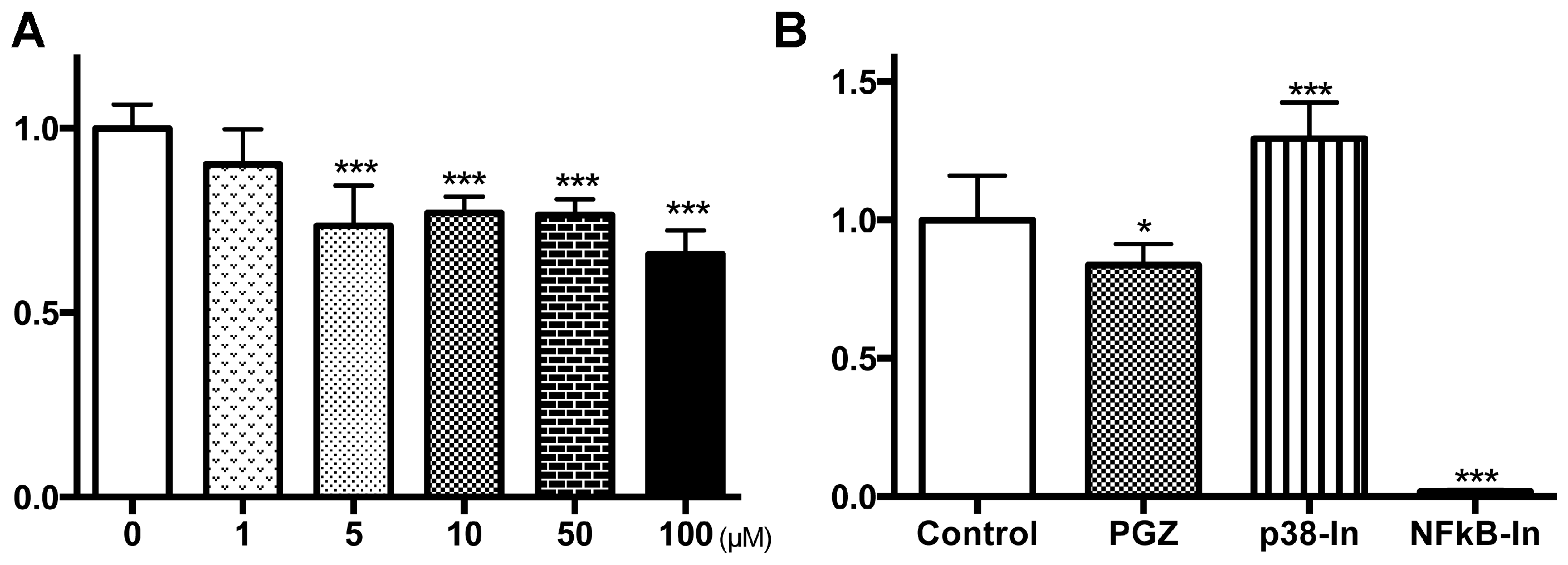

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGZ Treatment | No. of Rats | Body Weight (g) | Liver (g) | Kidneys (g) | Spleen (g) | Ventral Prostate (g) |
|---|---|---|---|---|---|---|
| Control | 10 | 506.6 ± 54.3 | 15.9 ± 1.5 | 3.0 ± 0.2 | 0.8 ± 0.1 | 0.30 ± 0.07 |
| 1 mg/kg | 8 | 531.9 ± 69.7 | 16.9 ± 2.6 | 3.0 ± 0.3 | 0.8 ± 0.2 | 0.27 ± 0.06 |
| 5 mg/kg | 9 | 531.4 ± 25.7 | 16.2 ± 1.5 | 3.0 ± 0.2 | 0.8 ± 0.1 | 0.24 ± 0.04 * |
| PGZ Treatment | No. of Rats | Ventral | Lateral | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Incidence of Carcinoma | % of Lesions in Prostate a | Incidence of Carcinoma | % of Lesions in Prostate a | ||||||
| LG-PIN | HG-PIN | Carcinoma | LG-PIN | HG-PIN | Carcinoma | ||||
| Control | 10 | 10 (100%) | 4.7 ± 1.9 | 80.7 ± 3.9 | 14.6 ± 3.7 | 8 (80%) | 18.6 ± 7.9 | 78.6 ± 10.0 | 2.7 ± 3.8 |
| 1 mg/kg | 8 | 8 (100%) | 7.5 ± 2.2 | 83.3 ± 2.6 | 9.2 ± 2.8 ** | 5 (63%) | 28.2 ± 15.8 | 70.7 ±16.3 | 1.0 ± 1.0 |
| 5 mg/kg | 9 | 9 (100%) | 10.4 ± 3.6 *** | 81.9 ± 4.2 | 7.7 ±3.3 *** | 4 (44%) | 25.9 ± 8.6 | 73.3 ± 8.7 | 0.8 ± 0.8 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, S.; Mori, Y.; Nagano, A.; Naiki-Ito, A.; Kato, H.; Nagayasu, Y.; Kobayashi, M.; Kuno, T.; Takahashi, S. Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. Int. J. Mol. Sci. 2016, 17, 2071. https://doi.org/10.3390/ijms17122071
Suzuki S, Mori Y, Nagano A, Naiki-Ito A, Kato H, Nagayasu Y, Kobayashi M, Kuno T, Takahashi S. Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. International Journal of Molecular Sciences. 2016; 17(12):2071. https://doi.org/10.3390/ijms17122071
Chicago/Turabian StyleSuzuki, Shugo, Yukiko Mori, Aya Nagano, Aya Naiki-Ito, Hiroyuki Kato, Yuko Nagayasu, Mizuho Kobayashi, Toshiya Kuno, and Satoru Takahashi. 2016. "Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis" International Journal of Molecular Sciences 17, no. 12: 2071. https://doi.org/10.3390/ijms17122071
APA StyleSuzuki, S., Mori, Y., Nagano, A., Naiki-Ito, A., Kato, H., Nagayasu, Y., Kobayashi, M., Kuno, T., & Takahashi, S. (2016). Pioglitazone, a Peroxisome Proliferator-Activated Receptor γ Agonist, Suppresses Rat Prostate Carcinogenesis. International Journal of Molecular Sciences, 17(12), 2071. https://doi.org/10.3390/ijms17122071