
Prolactin: Friend or Foe in Central Nervous System Autoimmune Inflammation?

Abstract
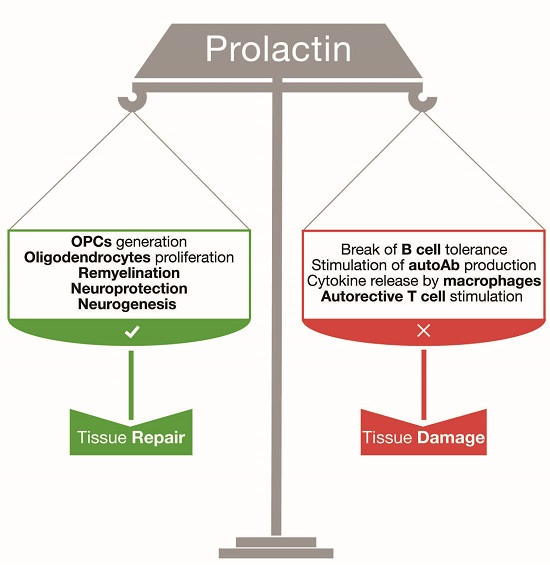
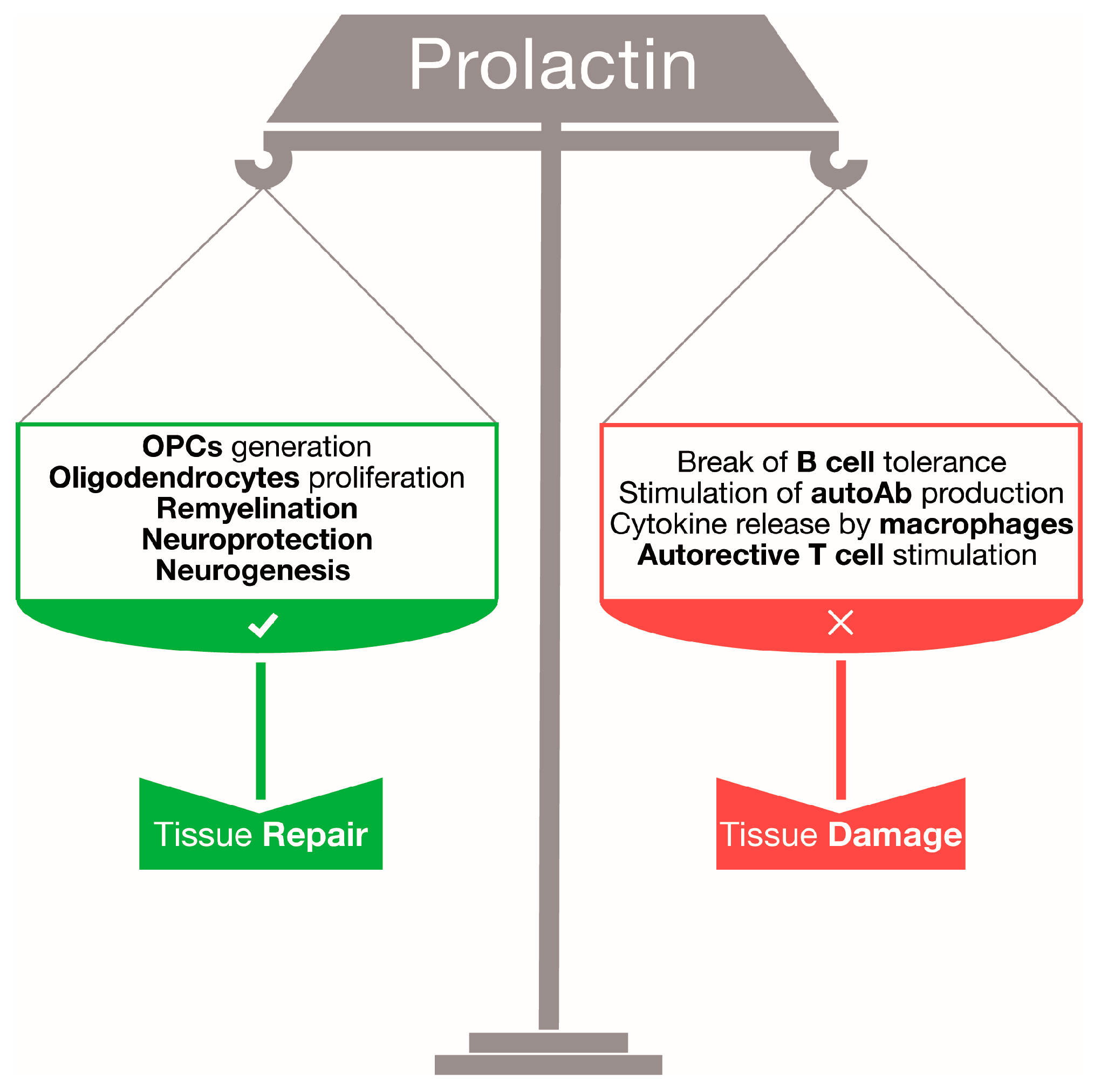
:
{kind=link}
{kind=link}
1. Introduction
2. Prolactin: A Regenerative Hormone for Central Nervous System Tissue
2.1. Prolactin Sustains Adult Neurogenesis
2.2. Prolactin as a Neuroprotective Factor
2.3. Promyelinating Effects of Prolactin
3. Prolactin and Central Nervous System Autoimmune Responses
3.1. The Immune-Stimulating Properties of Prolactin
3.2. Prolactin and Multiple Sclerosis
3.3. Prolactin and Experimental Autoimmune Encephalomyelitis
4. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| BCR | Bromocriptine |
| CNS | Central Nervous System |
| EAE | Experimental Autoimmune Encephalomyelitis |
| IFN | Interferon |
| IL | Interleukin |
| MOG | Myelin Oligodendrocyte Glycoprotein |
| MS | Multiple Sclerosis |
| OPC | Oligodendrocyte Progenitor Cell |
| PRL | Prolactin |
| Treg | Regulatory T cells |
References
- Steinman, L. Immunology of relapse and remission in multiple sclerosis. Annu. Rev. Immunol. 2014, 32, 257–281. [Google Scholar] [CrossRef] [PubMed]
- Goverman, J. Autoimmune T cell responses in the central nervous system. Nat. Rev. Immunol. 2009, 9, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Compston, A.; Coles, A. Multiple sclerosis. Lancet 2008, 372, 1502–1517. [Google Scholar] [CrossRef]
- Mahad, D.H.; Trapp, B.D.; Lassmann, H. Pathological mechanisms in progressive multiple sclerosis. Lancet Neurol. 2015, 14, 183–193. [Google Scholar] [CrossRef]
- Koch-Henriksen, N.; Sorensen, P.S. The changing demographic pattern of multiple sclerosis epidemiology. Lancet Neurol. 2010, 9, 520–532. [Google Scholar] [CrossRef]
- Confavreux, C.; Hutchinson, M.; Hours, M.M.; Cortinovis-Tourniaire, P.; Moreau, T. Rate of pregnancy-related relapse in multiple sclerosis. Pregnancy in multiple sclerosis group. N. Engl. J. Med. 1998, 339, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Young, J.; Chanson, P.; Binart, N. New insights in prolactin: Pathological implications. Nat. Rev. Endocrinol. 2015, 11, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Roelfsema, F.; Pijl, H.; Keenan, D.M.; Veldhuis, J.D. Prolactin secretion in healthy adults is determined by gender, age and body mass index. PLoS ONE 2012, 7, e31305. [Google Scholar] [CrossRef] [PubMed]
- Costanza, M.; Binart, N.; Steinman, L.; Pedotti, R. Prolactin: A versatile regulator of inflammation and autoimmune pathology. Autoimmun. Rev. 2015, 14, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Hellwig, K.; Haghikia, A.; Agne, H.; Beste, C.; Gold, R. Protective effect of breastfeeding in postpartum relapse rate of mothers with multiple sclerosis. Arch. Neurol. 2009, 66, 1580–1581. [Google Scholar] [CrossRef] [PubMed]
- Langer-Gould, A.; Huang, S.M.; Gupta, R.; Leimpeter, A.D.; Greenwood, E.; Albers, K.B.; van den Eeden, S.K.; Nelson, L.M. Exclusive breastfeeding and the risk of postpartum relapses in women with multiple sclerosis. Arch. Neurol. 2009, 66, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Vukusic, S.; Hutchinson, M.; Hours, M.; Moreau, T.; Cortinovis-Tourniaire, P.; Adeleine, P.; Confavreux, C. The pregnancy in multiple sclerosis (the prims study): Clinical predictors of post-partum relapse. Brain 2004, 127, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Iorio, R.; Nociti, V.; Frisullo, G.; Patanella, A.K.; Tonali, P.A.; Batocchi, A.P. Breastfeeding and multiple sclerosis. Arch. Neurol. 2009, 66, 1580–1581. [Google Scholar] [CrossRef] [PubMed]
- Airas, L.; Jalkanen, A.; Alanen, A.; Pirttila, T.; Marttila, R.J. Breast-feeding, postpartum and prepregnancy disease activity in multiple sclerosis. Neurology 2010, 75, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Ben-Jonathan, N.; LaPensee, C.R.; LaPensee, E.W. What can we learn from rodents about prolactin in humans? Endocr. Rev. 2008, 29, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Bole-Feysot, C.; Goffin, V.; Edery, M.; Binart, N.; Kelly, P.A. Prolactin (PRL) and its receptor: Actions, signal transduction pathways and phenotypes observed in PRL receptor knockout mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef] [PubMed]
- Clevenger, C.V.; Gadd, S.L.; Zheng, J. New mechanisms for PRLR action in breast cancer. Trends Endocrinol. Metab. 2009, 20, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.L. Molecular mechanisms of prolactin and its receptor. Endocr. Rev. 2012, 33, 504–525. [Google Scholar] [CrossRef] [PubMed]
- Dorshkind, K.; Horseman, N.D. The roles of prolactin, growth hormone, insulin-like growth factor-I, and thyroid hormones in lymphocyte development and function: Insights from genetic models of hormone and hormone receptor deficiency. Endocr. Rev. 2000, 21, 292–312. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.S.; Wyatt, A.K.; Herbison, R.E.; Knowles, P.J.; Ladyman, S.R.; Binart, N.; Banks, W.A.; Grattan, D.R. Prolactin transport into mouse brain is independent of prolactin receptor. FASEB J. 2016, 30, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Shingo, T.; Gregg, C.; Enwere, E.; Fujikawa, H.; Hassam, R.; Geary, C.; Cross, J.C.; Weiss, S. Pregnancy-stimulated neurogenesis in the adult female forebrain mediated by prolactin. Science 2003, 299, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Mak, G.K.; Weiss, S. Paternal recognition of adult offspring mediated by newly generated CNS neurons. Nat. Neurosci. 2010, 13, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.L.; Vukovic, J.; Koudijs, M.M.; Blackmore, D.G.; Mackay, E.W.; Sykes, A.M.; Overall, R.W.; Hamlin, A.S.; Bartlett, P.F. Prolactin stimulates precursor cells in the adult mouse hippocampus. PLoS ONE 2012, 7, e44371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torner, L.; Karg, S.; Blume, A.; Kandasamy, M.; Kuhn, H.G.; Winkler, J.; Aigner, L.; Neumann, I.D. Prolactin prevents chronic stress-induced decrease of adult hippocampal neurogenesis and promotes neuronal fate. J. Neurosci. 2009, 29, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Farooq, F.; Molina, F.A.; Hadwen, J.; MacKenzie, D.; Witherspoon, L.; Osmond, M.; Holcik, M.; MacKenzie, A. Prolactin increases SMN expression and survival in a mouse model of severe spinal muscular atrophy via the STAT5 pathway. J. Clin. Investig. 2011, 121, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Tejadilla, D.; Cerbon, M.; Morales, T. Prolactin reduces the damaging effects of excitotoxicity in the dorsal hippocampus of the female rat independently of ovarian hormones. Neuroscience 2010, 169, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Castaneda, E.; Grattan, D.R.; Pasantes-Morales, H.; Perez-Dominguez, M.; Cabrera-Reyes, E.A.; Morales, T.; Cerbon, M. Prolactin mediates neuroprotection against excitotoxicity in primary cell cultures of hippocampal neurons via its receptor. Brain Res. 2016, 1636, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Arnold, E.; Thebault, S.; Baeza-Cruz, G.; Arredondo Zamarripa, D.; Adan, N.; Quintanar-Stephano, A.; Condes-Lara, M.; Rojas-Piloni, G.; Binart, N.; Martinez de la Escalera, G.; et al. The hormone prolactin is a novel, endogenous trophic factor able to regulate reactive glia and to limit retinal degeneration. J. Neurosci. 2014, 34, 1868–1878. [Google Scholar] [CrossRef] [PubMed]
- Gregg, C.; Shikar, V.; Larsen, P.; Mak, G.; Chojnacki, A.; Yong, V.W.; Weiss, S. White matter plasticity and enhanced remyelination in the maternal CNS. J. Neurosci. 2007, 27, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Majumder, B.; Biswas, R.; Chattopadhyay, U. Prolactin regulates antitumor immune response through induction of tumoricidal macrophages and release of IL-12. Int. J. Cancer 2002, 97, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Musumeci, M.; Licata, F.; Di Rosa, M.; Messina, A.; Musumeci, S. Prolactin induces chitotriosidase gene expression in human monocyte-derived macrophages. Immunol. Lett. 2004, 94, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, A.; Tripathi, A. Prolactin and growth hormone induce differential cytokine and chemokine profile in murine peritoneal macrophages in vitro: Involvement of p-38 MAP kinase, STAT3 and NF-κB. Cytokine 2008, 41, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Matera, L.; Galetto, A.; Geuna, M.; Vekemans, K.; Ricotti, E.; Contarini, M.; Moro, F.; Basso, G. Individual and combined effect of granulocyte-macrophage colony-stimulating factor and prolactin on maturation of dendritic cells from blood monocytes under serum-free conditions. Immunology 2000, 100, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Matera, L.; Mori, M.; Galetto, A. Effect of prolactin on the antigen presenting function of monocyte-derived dendritic cells. Lupus 2001, 10, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Sabharwal, P.; Glaser, R.; Lafuse, W.; Varma, S.; Liu, Q.; Arkins, S.; Kooijman, R.; Kutz, L.; Kelley, K.W.; Malarkey, W.B. Prolactin synthesized and secreted by human peripheral blood mononuclear cells: An autocrine growth factor for lymphoproliferation. Proc. Natl. Acad. Sci. USA 1992, 89, 7713–7716. [Google Scholar] [CrossRef] [PubMed]
- Bauernhofer, T.; Kuss, I.; Friebe-Hoffmann, U.; Baum, A.S.; Dworacki, G.; Vonderhaar, B.K.; Whiteside, T.L. Role of prolactin receptor and CD25 in protection of circulating T lymphocytes from apoptosis in patients with breast cancer. Br. J. Cancer 2003, 88, 1301–1309. [Google Scholar] [CrossRef] [PubMed]
- Costanza, M.; Musio, S.; Abou-Hamdan, M.; Binart, N.; Pedotti, R. Prolactin is not required for the development of severe chronic experimental autoimmune encephalomyelitis. J. Immunol. 2013, 191, 2082–2088. [Google Scholar] [CrossRef] [PubMed]
- Goffin, V.; Binart, N.; Touraine, P.; Kelly, P.A. Prolactin: The new biology of an old hormone. Annu. Rev. Physiol. 2002, 64, 47–67. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; de Rosa, V.; Carrieri, P.B.; Montella, S.; Bruzzese, D.; Porcellini, A.; Procaccini, C.; La Cava, A.; Matarese, G. Regulatory T cell proliferative potential is impaired in human autoimmune disease. Nat. Med. 2014, 20, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Legorreta-Haquet, M.V.; Chavez-Rueda, K.; Montoya-Diaz, E.; Arriaga-Pizano, L.; Silva-Garcia, R.; Chavez-Sanchez, L.; Moreno-Lafont, M.; Zenteno-Galindo, E.; Blanco-Favela, F. Prolactin down-regulates CD4+CD25highCD127low/− regulatory T cell function in humans. J. Mol. Endocrinol. 2012, 48, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Tsokos, G.C. Systemic lupus erythematosus. N. Engl. J. Med. 2011, 365, 2110–2121. [Google Scholar] [CrossRef] [PubMed]
- McMurray, R.; Keisler, D.; Kanuckel, K.; Izui, S.; Walker, S.E. Prolactin influences autoimmune disease activity in the female B/W mouse. J. Immunol. 1991, 147, 3780–3787. [Google Scholar] [PubMed]
- McMurray, R.; Keisler, D.; Izui, S.; Walker, S.E. Hyperprolactinemia in male NZB/NZW (B/W) F1 mice: Accelerated autoimmune disease with normal circulating testosterone. Clin. Immunol. Immunopathol. 1994, 71, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Peeva, E.; Michael, D.; Cleary, J.; Rice, J.; Chen, X.; Diamond, B. Prolactin modulates the naive B cell repertoire. J. Clin. Investig. 2003, 111, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Peeva, E.; Gonzalez, J.; Hicks, R.; Diamond, B. Cutting edge: Lupus susceptibility interval SLE3/5 confers responsiveness to prolactin in C57BL/6 mice. J. Immunol. 2006, 177, 1401–1405. [Google Scholar] [CrossRef] [PubMed]
- Correale, J.; Farez, M.F.; Ysrraelit, M.C. Role of prolactin in B cell regulation in multiple sclerosis. J. Neuroimmunol. 2014, 269, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Markianos, M.; Koutsis, G.; Evangelopoulos, M.E.; Mandellos, D.; Sfagos, C. Serum and cerebrospinal fluid prolactin levels in male and female patients with clinically-isolated syndrome or relapsing-remitting multiple sclerosis. J. Neuroendocrinol. 2010, 22, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Moshirzadeh, S.; Ghareghozli, K.; Harandi, A.A.; Pakdaman, H. Serum prolactin level in patients with relapsing-remitting multiple sclerosis during relapse. J. Clin. Neurosci. 2012, 19, 622–623. [Google Scholar] [CrossRef] [PubMed]
- De Giglio, L.; Marinelli, F.; Prosperini, L.; Contessa, G.M.; Gurreri, F.; Piattella, M.C.; de Angelis, F.; Barletta, V.T.; Tomassini, V.; Pantano, P.; et al. Relationship between prolactin plasma levels and white matter volume in women with multiple sclerosis. Mediat. Inflamm. 2015, 2015, 732539. [Google Scholar] [CrossRef] [PubMed]
- Shelly, S.; Boaz, M.; Orbach, H. Prolactin and autoimmunity. Autoimmun. Rev. 2012, 11, A465–A470. [Google Scholar] [CrossRef] [PubMed]
- Nociti, V.; Frisullo, G.; Tartaglione, T.; Patanella, A.K.; Iorio, R.; Tonali, P.A.; Batocchi, A.P. Multiple sclerosis attacks triggered by hyperprolactinemia. J. Neurooncol. 2010, 98, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Bissay, V.; de Klippel, N.; Herroelen, L.; Schmedding, E.; Buisseret, T.; Ebinger, G.; de Keyser, J. Bromocriptine therapy in multiple sclerosis: An open label pilot study. Clin. Neuropharmacol. 1994, 17, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Postert, T.; McMonagle, U.; Buttner, T.; Pohlau, D.; Meves, S.; Przuntek, H. Paroxysmal convergence spasm in multiple sclerosis. Acta Neurol. Scand. 1996, 94, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, K.; Oseko, F.; Morikawa, S. Immunosuppressive property of bromocriptine on human B lymphocyte function in vitro. Clin. Exp. Immunol. 1993, 93, 200–205. [Google Scholar] [CrossRef]
- Morikawa, K.; Oseko, F.; Morikawa, S. Immunosuppressive activity of bromocriptine on human T lymphocyte function in vitro. Clin. Exp. Immunol. 1994, 95, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Stromnes, I.M.; Goverman, J.M. Active induction of experimental allergic encephalomyelitis. Nat. Protoc. 2006, 1, 1810–1819. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L.; Zamvil, S.S. Virtues and pitfalls of EAE for the development of therapies for multiple sclerosis. Trends Immunol. 2005, 26, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Riskind, P.N.; Massacesi, L.; Doolittle, T.H.; Hauser, S.L. The role of prolactin in autoimmune demyelination: Suppression of experimental allergic encephalomyelitis by bromocriptine. Ann. Neurol. 1991, 29, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, C.D.; van der Voort, E.R.; de Groot, C.J.; Huitinga, I.; Uitdehaag, B.M.; Polman, C.H.; Berkenbosch, F. Therapeutic effect of the D2-dopamine agonist bromocriptine on acute and relapsing experimental allergic encephalomyelitis. Psychoneuroendocrinology 1994, 19, 135–142. [Google Scholar] [CrossRef]
- Zhornitsky, S.; Johnson, T.A.; Metz, L.M.; Weiss, S.; Yong, V.W. Prolactin in combination with interferon-β reduces disease severity in an animal model of multiple sclerosis. J. Neuroinflamm. 2015, 12, 55. [Google Scholar] [CrossRef] [PubMed]
- Lyons, J.A.; San, M.; Happ, M.P.; Cross, A.H. B cells are critical to induction of experimental allergic encephalomyelitis by protein but not by a short encephalitogenic peptide. Eur. J. Immunol. 1999, 29, 3432–3439. [Google Scholar] [CrossRef]
- Hilfiker-Kleiner, D.; Kaminski, K.; Podewski, E.; Bonda, T.; Schaefer, A.; Sliwa, K.; Forster, O.; Quint, A.; Landmesser, U.; Doerries, C.; et al. A cathepsin d-cleaved 16 kDa form of prolactin mediates postpartum cardiomyopathy. Cell 2007, 128, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Horseman, N.D.; Gregerson, K.A. Prolactin actions. J. Mol. Endocrinol. 2014, 52, R95–R106. [Google Scholar] [CrossRef] [PubMed]
- Berwaer, M.; Martial, J.A.; Davis, J.R. Characterization of an up-stream promoter directing extrapituitary expression of the human prolactin gene. Mol. Endocrinol. 1994, 8, 635–642. [Google Scholar] [PubMed]
- Gellersen, B.; Kempf, R.; Telgmann, R.; DiMattia, G.E. Nonpituitary human prolactin gene transcription is independent of Pit-1 and differentially controlled in lymphocytes and in endometrial stroma. Mol. Endocrinol. 1994, 8, 356–373. [Google Scholar] [PubMed]
- Christensen, H.R.; Murawsky, M.K.; Horseman, N.D.; Willson, T.A.; Gregerson, K.A. Completely humanizing prolactin rescues infertility in prolactin knockout mice and leads to human prolactin expression in extrapituitary mouse tissues. Endocrinology 2013, 154, 4777–4789. [Google Scholar] [CrossRef] [PubMed]
- ‘T Hart, B.A.; van Kooyk, Y.; Geurts, J.J.; Gran, B. The primate autoimmune encephalomyelitis model; a bridge between mouse and man. Ann. Clin. Transl. Neurol. 2015, 2, 581–593. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costanza, M.; Pedotti, R. Prolactin: Friend or Foe in Central Nervous System Autoimmune Inflammation? Int. J. Mol. Sci. 2016, 17, 2026. https://doi.org/10.3390/ijms17122026
Costanza M, Pedotti R. Prolactin: Friend or Foe in Central Nervous System Autoimmune Inflammation? International Journal of Molecular Sciences. 2016; 17(12):2026. https://doi.org/10.3390/ijms17122026
Chicago/Turabian StyleCostanza, Massimo, and Rosetta Pedotti. 2016. "Prolactin: Friend or Foe in Central Nervous System Autoimmune Inflammation?" International Journal of Molecular Sciences 17, no. 12: 2026. https://doi.org/10.3390/ijms17122026