
SEPT12–NDC1 Complexes Are Required for Mammalian Spermiogenesis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. Male Infertility
1.2. SEPTINs (SEPTs) and Spermatogenesis
1.3. Nuclear Pore Complexes and NDC1
2. Results
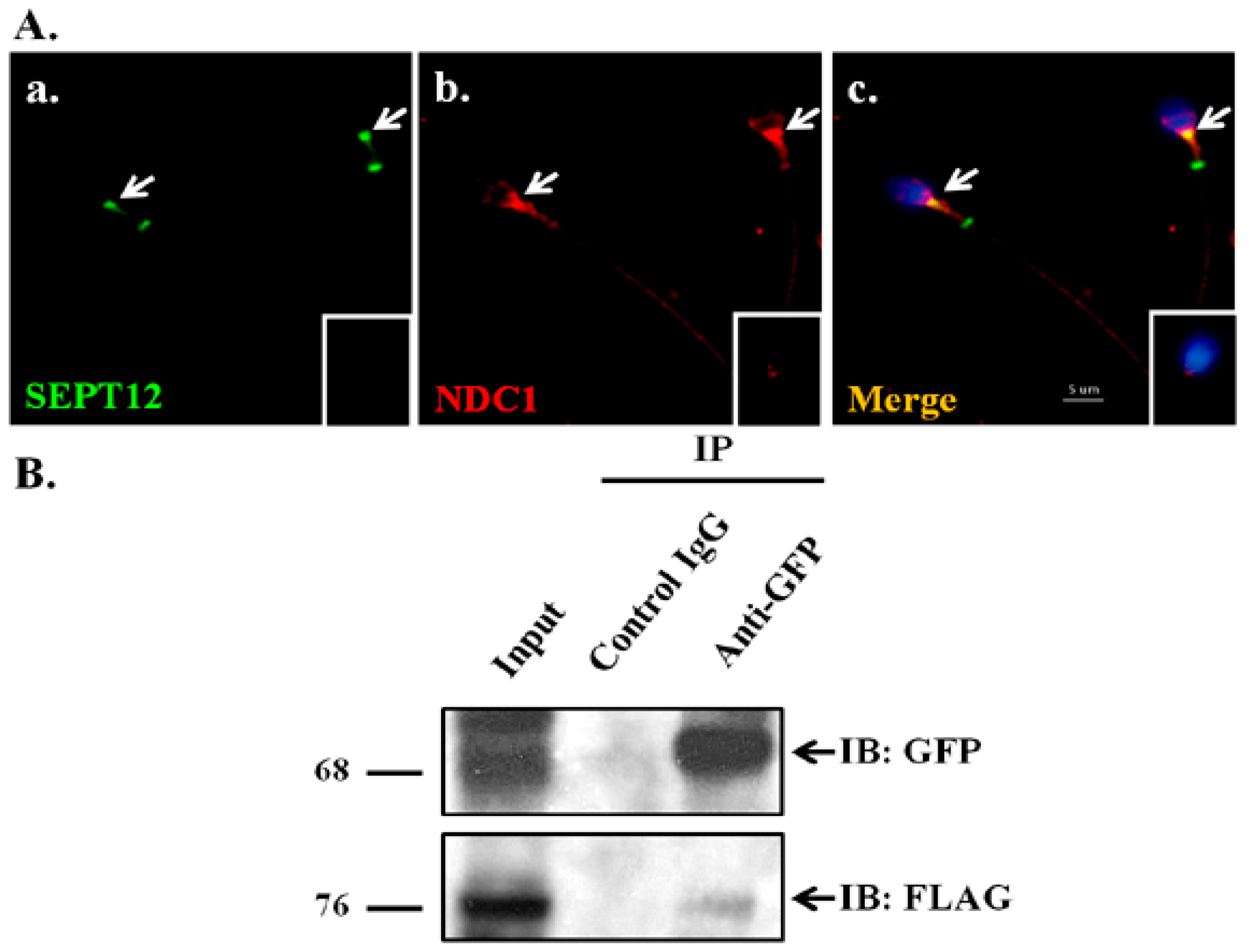
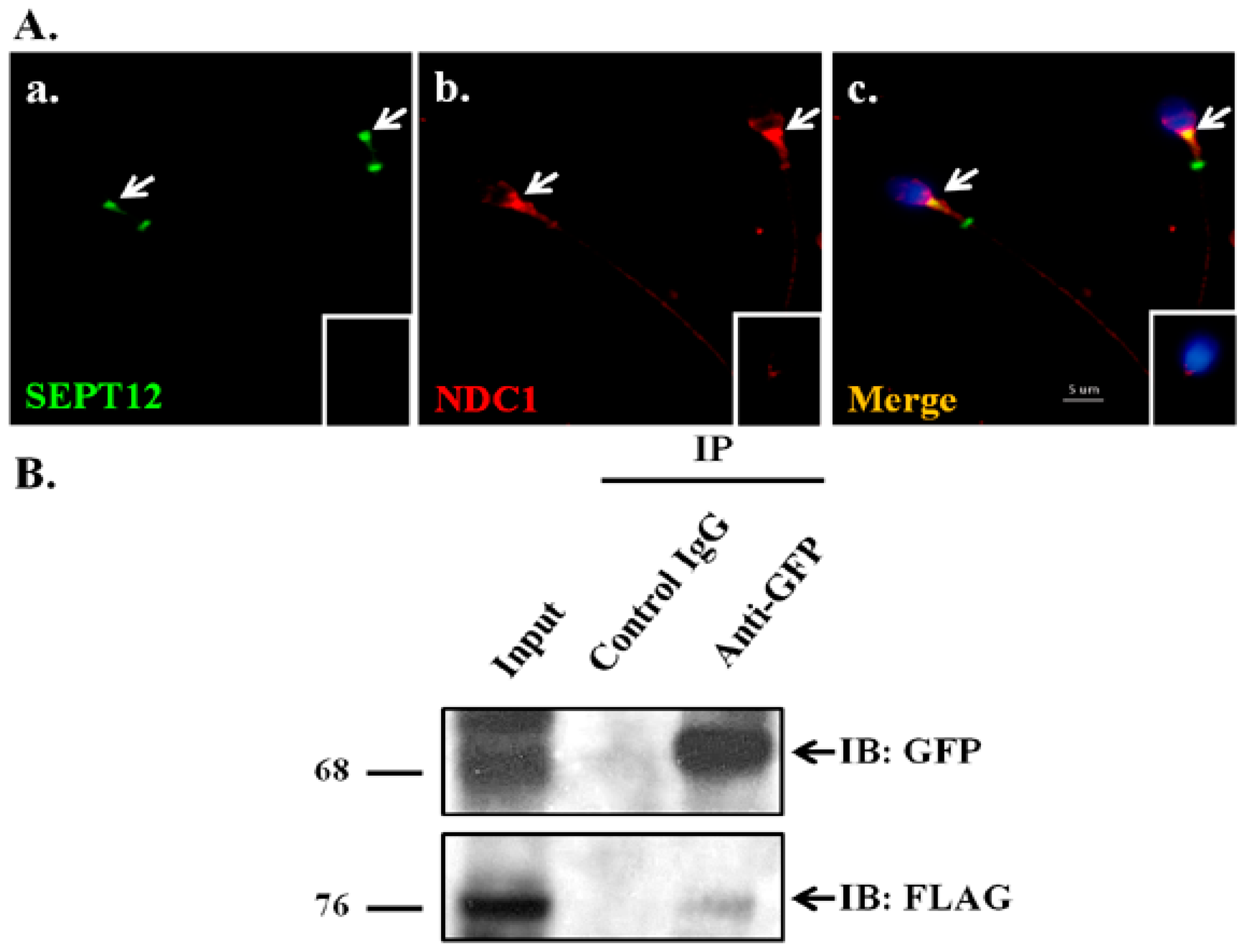
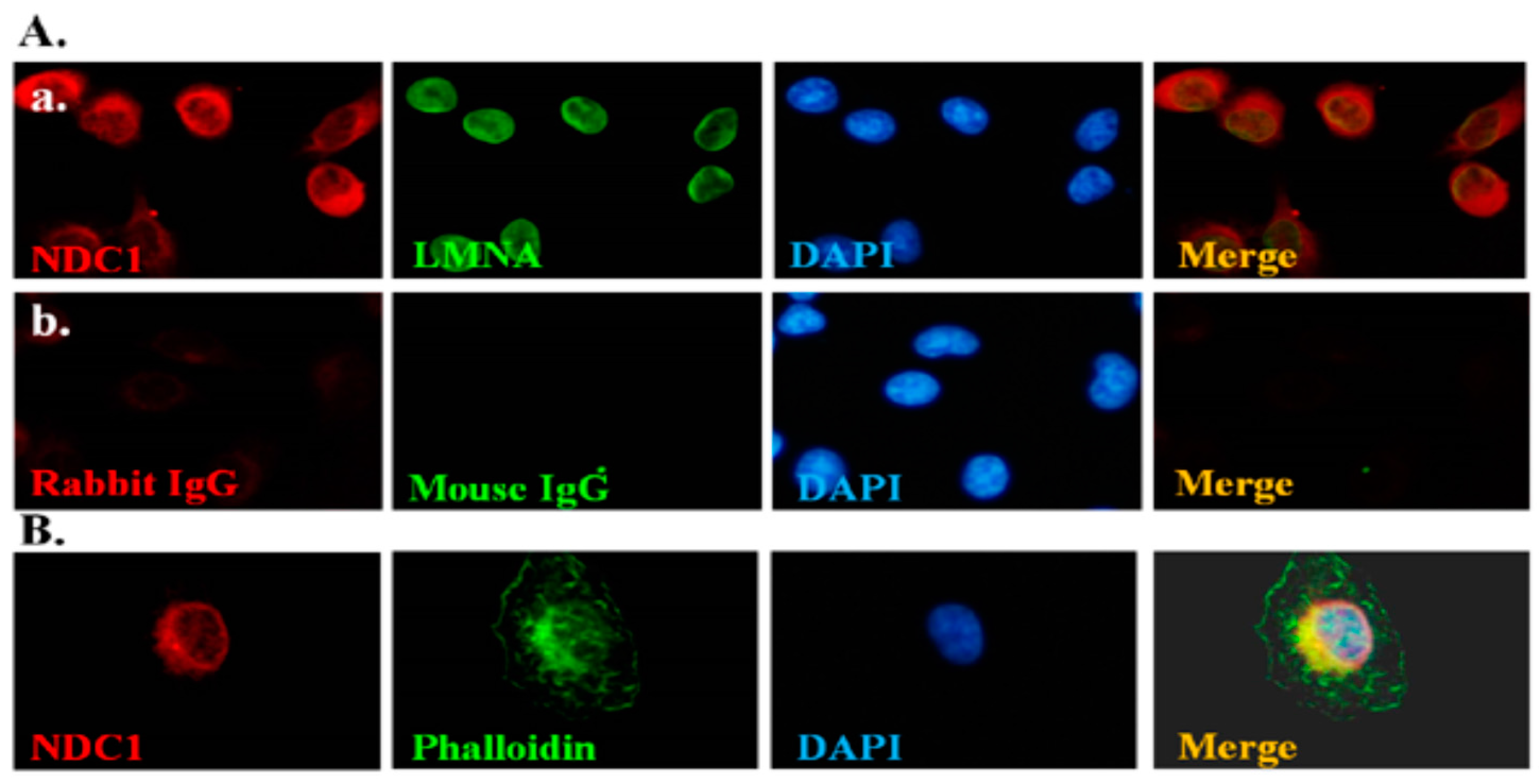
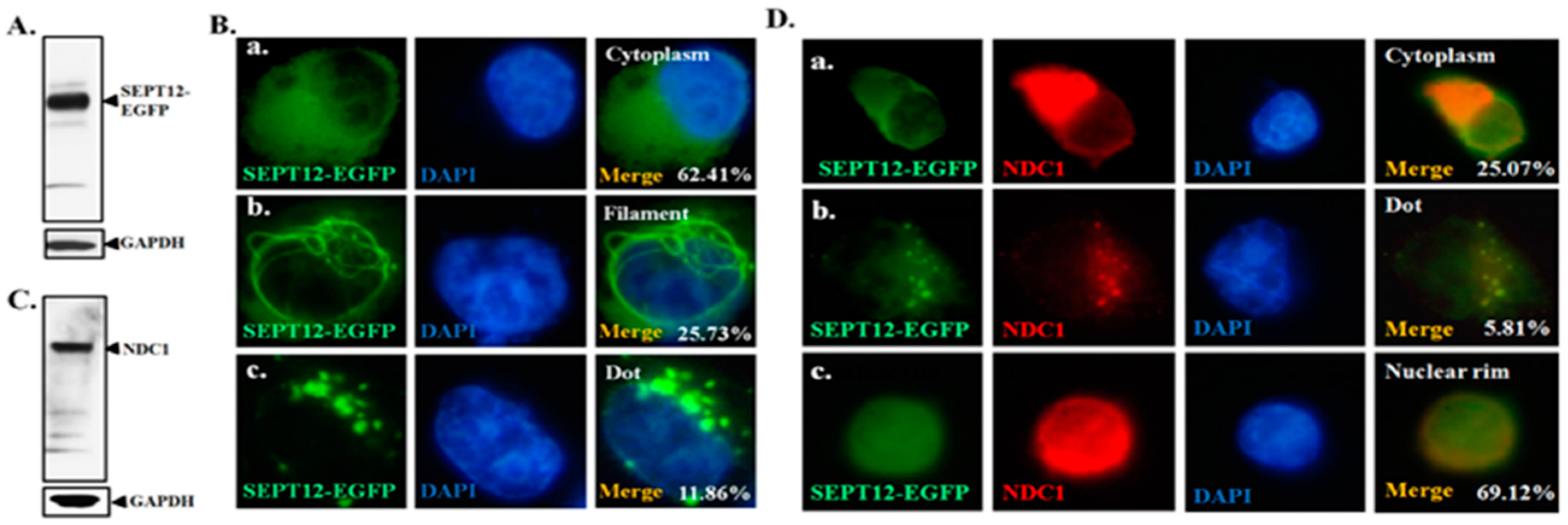
2.1. NDC1 as a SEPT12 Interactor
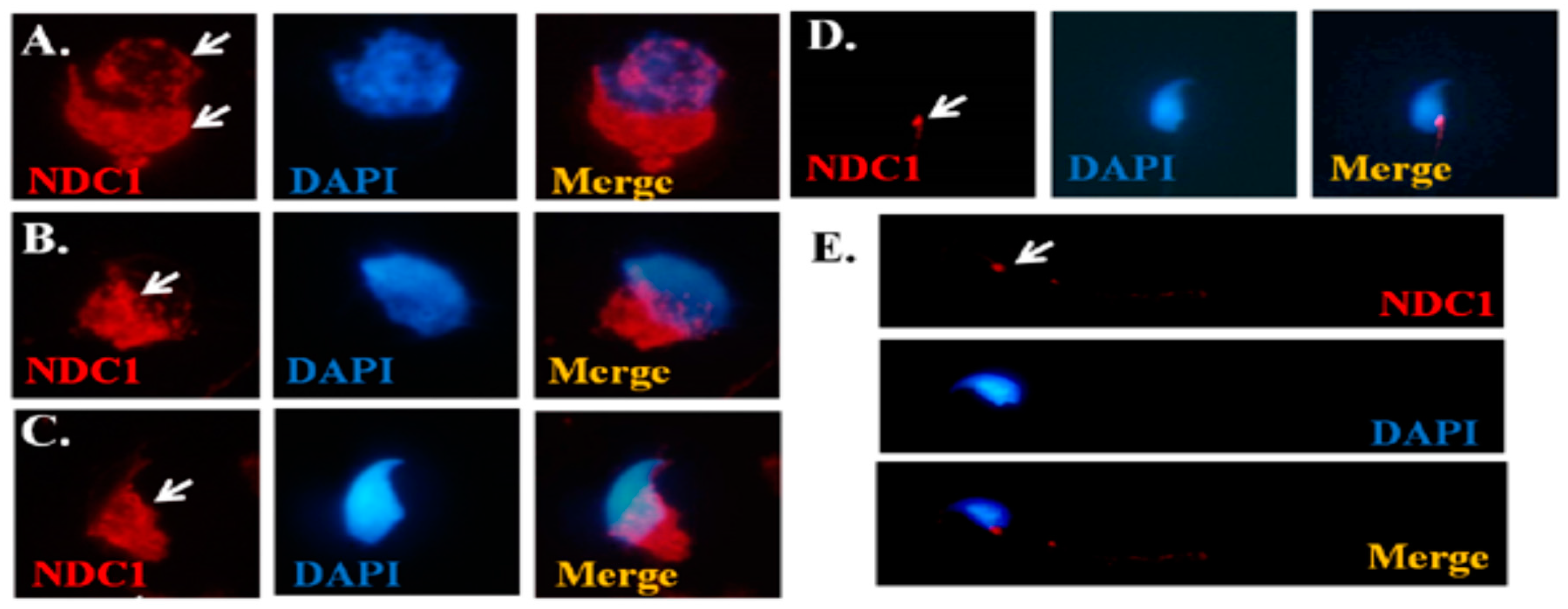
2.2. Dynamic Expression of NDC1 during Murine Spermiogenesis
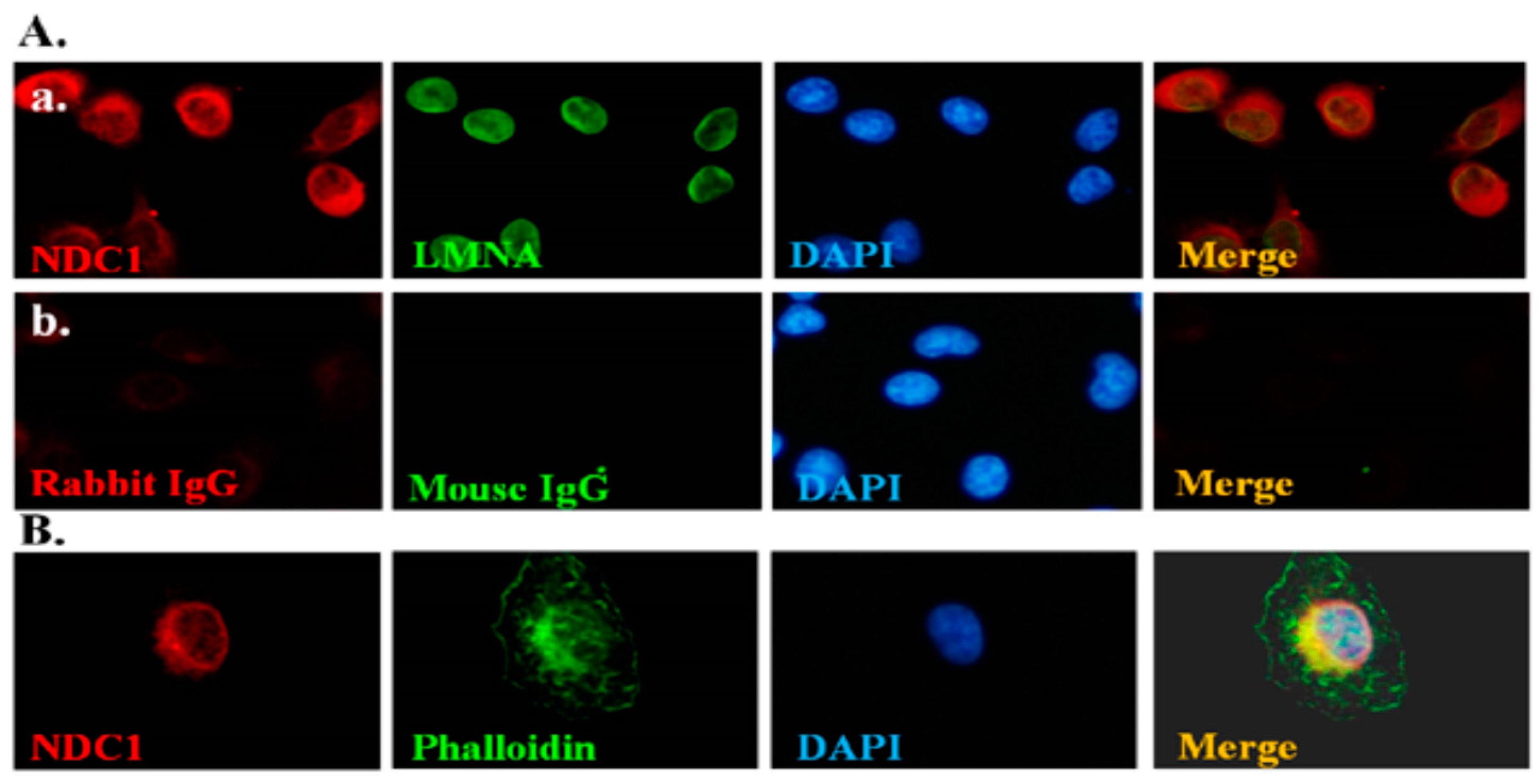
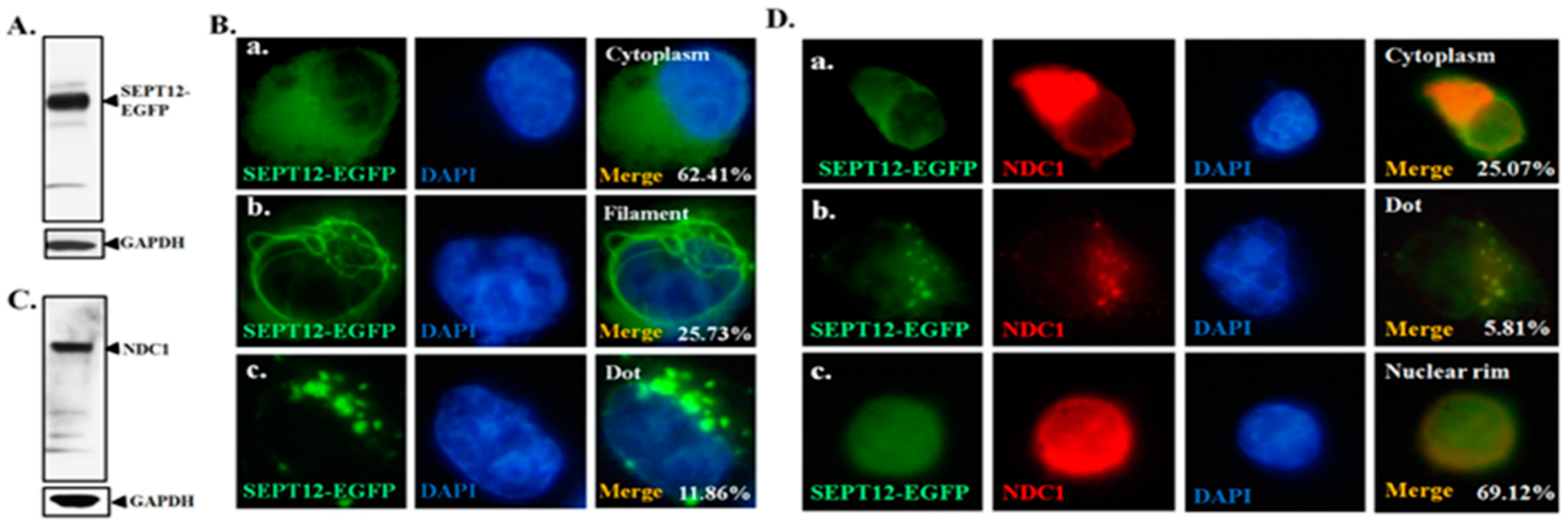
2.3. Effect of NDC1 on SEPT12 Localization and Filamental Structure in a Cell Model
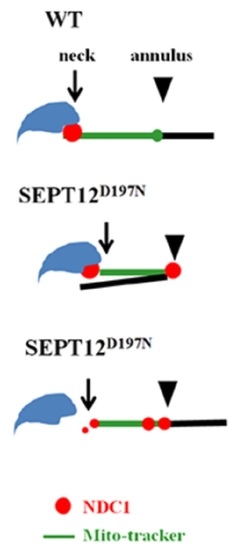
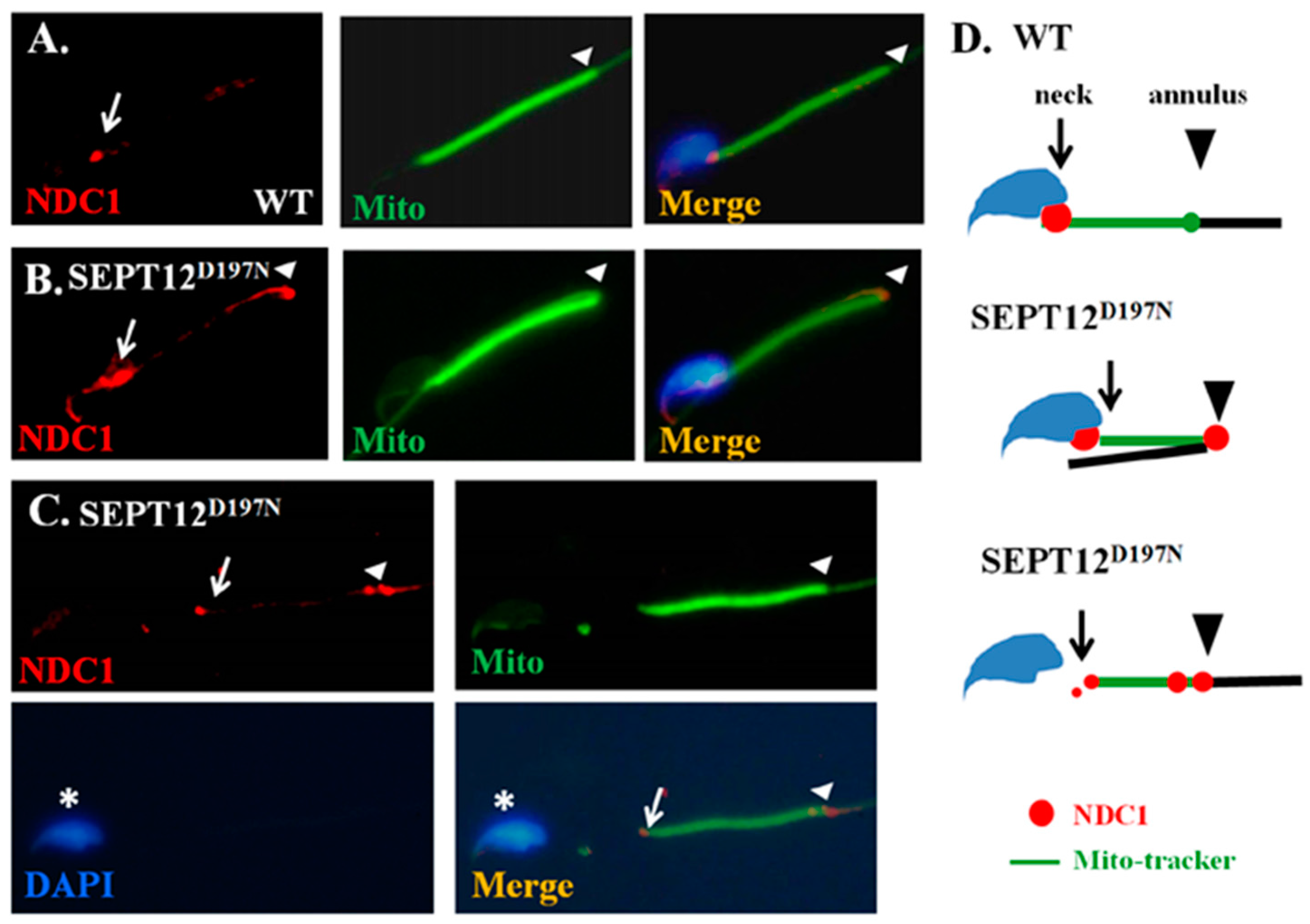
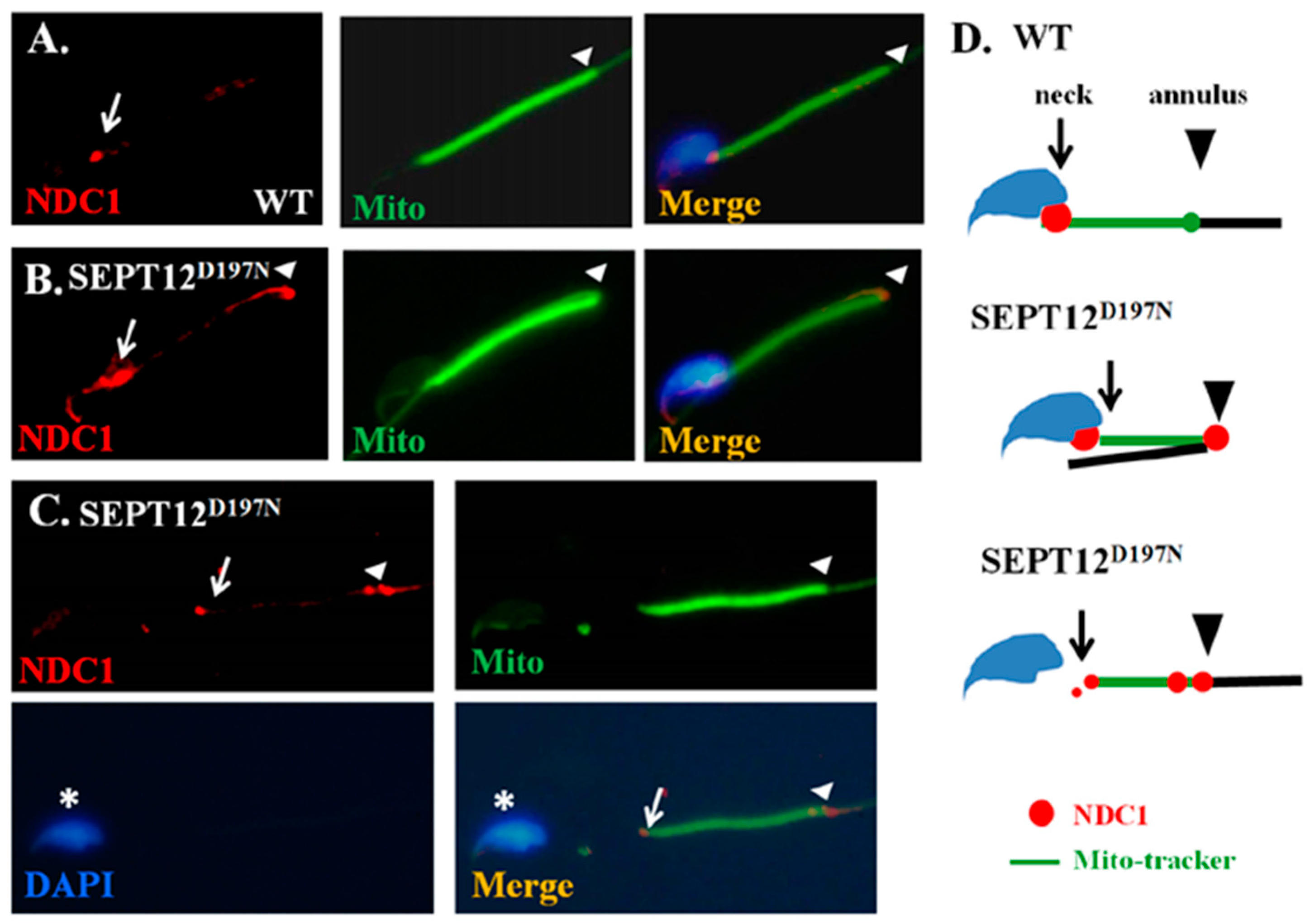
2.4. Effect of Mutated SEPT12 on NDC1 Localization In Vivo
3. Discussion
3.1. SEPT12 Interaction with Nuclear and Nuclear-Related Proteins
3.2. Male Reproductive Roles of SEPT12 and NDC1
3.3. Effect of Mutated SEPT12 on Cellular Localization of NDC1
4. Materials and Methods
4.1. Immunofluorescence Assay
4.2. Cloning, Transfection, and Coimmunoprecipitation Assay
4.3. Separation of the Murine Testicular Germ Cell Populations and Sperm Preparation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO, World Health Organization. Examination of Human Semen and Semen-Cervical Mucus Interaction; World Health Organization: Geneva, Switzerland, 1992. [Google Scholar]
- Palermo, G.; Joris, H.; Devroey, P.; van Steirteghem, A.C. Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet 1992, 340, 17–18. [Google Scholar] [CrossRef]
- Javed, M.; Esfandiari, N.; Casper, R.F. Failed fertilization after clinical intracytoplasmic sperm injection. Reprod. Biomed. Online 2010, 20, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Cossart, P. Septins: The fourth component of the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2012, 13, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.A.; Russell, S.E. The pathobiology of the septin gene family. J. Pathol. 2004, 204, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Kuo, Y.C.; Chiang, H.S.; Kuo, P.L. The role of the septin family in spermiogenesis. Spermatogenesis 2011, 1, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O.; et al. Cortical organization by the septin cytoskeleton is essential for structural and mechanical integrity of mammalian spermatozoa. Dev. Cell 2005, 8, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Kissel, H.; Georgescu, M.M.; Larisch, S.; Manova, K.; Hunnicutt, G.R.; Steller, H. The Sept4 septin locus is required for sperm terminal differentiation in mice. Dev. Cell 2005, 8, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Lhuillier, P.; Rode, B.; Escalier, D.; Lores, P.; Dirami, T.; Bienvenu, T.; Gacon, G.; Dulioust, E.; Toure, A. Absence of annulus in human asthenozoospermia: Case report. Hum. Reprod. 2009, 24, 1296–1303. [Google Scholar] [CrossRef] [PubMed]
- Sugino, Y.; Ichioka, K.; Soda, T.; Ihara, M.; Kinoshita, M.; Ogawa, O.; Nishiyama, H. Septins as diagnostic markers for a subset of human asthenozoospermia. J. Urol. 2008, 180, 2706–2709. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Wang, Y.Y.; Yu, I.S.; Lin, Y.W.; Wang, Y.H.; Wu, C.M.; Pan, H.A.; Chao, S.C.; Yen, P.H.; et al. The expression level of septin12 is critical for spermiogenesis. Am. J. Pathol. 2009, 174, 1857–1868. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Teng, Y.N.; Hsieh, T.Y.; Lin, Y.S.; Kuo, P.L. Identification of ten novel genes involved in human spermatogenesis by microarray analysis of testicular tissue. Fertil. Steril. 2006, 86, 1650–1658. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Chou, C.K.; Hung, Y.C.; Yu, I.S.; Pan, H.A.; Lin, S.W.; Kuo, P.L. SEPT12 deficiency causes sperm nucleus damage and developmental arrest of preimplantation embryos. Fertil. Steril. 2011, 95, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.C.; Lin, Y.H.; Chen, H.I.; Wang, Y.Y.; Chiou, Y.W.; Lin, H.H.; Pan, H.A.; Wu, C.M.; Su, S.M.; Hsu, C.C.; et al. SEPT12 mutations cause male infertility with defective sperm annulus. Hum. Mutat. 2012, 33, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Wang, Y.Y.; Chen, H.I.; Kuo, Y.C.; Chiou, Y.W.; Lin, H.H.; Wu, C.M.; Hsu, C.C.; Chiang, H.S.; Kuo, P.L. SEPTIN12 genetic variants confer susceptibility to teratozoospermia. PLoS ONE 2012, 7, e34011. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.L.; Chiang, H.S.; Wang, Y.Y.; Kuo, Y.C.; Chen, M.F.; Yu, I.S.; Teng, Y.N.; Lin, S.W.; Lin, Y.H. SEPT12-microtubule complexes are required for sperm head and tail formation. Int. J. Mol. Sci. 2013, 14, 22102–22116. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Kuo, P.L.; Wang, Y.Y.; Wu, Y.Y.; Chen, M.F.; Lin, D.Y.; Lai, T.H.; Chiang, H.S.; Lin, Y.H. SEPT12/SPAG4/LAMINB1 complexes are required for maintaining the integrity of the nuclear envelope in postmeiotic male germ cells. PLoS ONE 2015, 10, e0120722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lei, K.; Yuan, X.; Wu, X.; Zhuang, Y.; Xu, T.; Xu, R.; Han, M. SUN1/2 and Syne/Nesprin-1/2 complexes connect centrosome to the nucleus during neurogenesis and neuronal migration in mice. Neuron 2009, 64, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Chatel, G.; Fahrenkrog, B. Nucleoporins: Leaving the nuclear pore complex for a successful mitosis. Cell Signal. 2011, 23, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Strambio-De-Castillia, C.; Niepel, M.; Rout, M.P. The nuclear pore complex: Bridging nuclear transport and gene regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Antonin, W.; Franz, C.; Haselmann, U.; Antony, C.; Mattaj, I.W. The integral membrane nucleoporin pom121 functionally links nuclear pore complex assembly and nuclear envelope formation. Mol. Cell 2005, 17, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Mansfeld, J.; Guttinger, S.; Hawryluk-Gara, L.A.; Pante, N.; Mall, M.; Galy, V.; Haselmann, U.; Muhlhausser, P.; Wozniak, R.W.; Mattaj, I.W.; et al. The conserved transmembrane nucleoporin NDC1 is required for nuclear pore complex assembly in vertebrate cells. Mol. Cell 2006, 22, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Galy, V.; Antonin, W.; Jaedicke, A.; Sachse, M.; Santarella, R.; Haselmann, U.; Mattaj, I. A role for gp210 in mitotic nuclear-envelope breakdown. J. Cell Sci. 2008, 121, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Handel, M.A.; Lane, P.W.; Schroeder, A.C.; Davisson, M.T. New mutation causing sterility in the mouse. Gamete Res. 1988, 21, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Noguchi, J.; Hirose, M.; Kajita, S.; Katayama, K.; Khalaj, M.; Tsuji, T.; Fairfield, H.; Byers, C.; Reinholdt, L.; et al. A mutation in the nuclear pore complex gene Tmem48 causes gametogenesis defects in skeletal fusions with sterility (sks) mice. J. Biol. Chem. 2013, 288, 31830–31841. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Wu, Y.Y. Staining Anti-NDC1 Antibody on Mouse Sperm; Graduate Institute of Biomedical and Pharmaceutical Science, Fu Jen Catholic University: New Taipei City, Taiwan, 2015. [Google Scholar]
- Kuo, Y.C.; Shen, Y.R.; Chen, H.I.; Lin, Y.H.; Wang, Y.Y.; Chen, Y.R.; Wang, C.Y.; Kuo, P.L. SEPT12 orchestrates the formation of mammalian sperm annulus by organizing core octameric complexes with other SEPT proteins. J. Cell Sci. 2015, 128, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.; Willis, W.D.; Goulding, E.H.; Jung-Ha, H.; Choi, Y.C.; Hecht, N.B.; Eddy, E.M. Haploinsufficiency of protamine-1 or -2 causes infertility in mice. Nat. Genet. 2001, 28, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Haque, F.; Lloyd, D.J.; Smallwood, D.T.; Dent, C.L.; Shanahan, C.M.; Fry, A.M.; Trembath, R.C.; Shackleton, S. SUN1 interacts with nuclear lamin A and cytoplasmic nesprins to provide a physical connection between the nuclear lamina and the cytoskeleton. Mol. Cell. Biol. 2006, 26, 3738–3751. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Smoyer, C.J.; Slaughter, B.D.; Unruh, J.R.; Jaspersen, S.L. The SUN protein Mps3 controls Ndc1 distribution and function on the nuclear membrane. J. Cell Biol. 2014, 204, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Kierszenbaum, A.L.; Tres, L.L. The acrosome-acroplaxome-manchette complex and the shaping of the spermatid head. Arch. Histol. Cytol. 2004, 67, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Kierszenbaum, A.L.; Rivkin, E.; Tres, L.L. The actin-based motor myosin Va is a component of the acroplaxome, an acrosome-nuclear envelope junctional plate, and of manchette-associated vesicles. Cytogenet. Genome Res. 2003, 103, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.H.; Lin, Y.M.; Kuo, Y.C.; Wang, Y.Y.; Kuo, P.L. Identification and characterization of a novel Rab GTPase-activating protein in spermatids. Int. J. Androl. 2011, 34, 358–367. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, T.-H.; Wu, Y.-Y.; Wang, Y.-Y.; Chen, M.-F.; Wang, P.; Chen, T.-M.; Wu, Y.-N.; Chiang, H.-S.; Kuo, P.-L.; Lin, Y.-H. SEPT12–NDC1 Complexes Are Required for Mammalian Spermiogenesis. Int. J. Mol. Sci. 2016, 17, 1911. https://doi.org/10.3390/ijms17111911
Lai T-H, Wu Y-Y, Wang Y-Y, Chen M-F, Wang P, Chen T-M, Wu Y-N, Chiang H-S, Kuo P-L, Lin Y-H. SEPT12–NDC1 Complexes Are Required for Mammalian Spermiogenesis. International Journal of Molecular Sciences. 2016; 17(11):1911. https://doi.org/10.3390/ijms17111911
Chicago/Turabian StyleLai, Tsung-Hsuan, Ying-Yu Wu, Ya-Yun Wang, Mei-Feng Chen, Pei Wang, Tsung-Ming Chen, Yi-No Wu, Han-Sun Chiang, Pao-Lin Kuo, and Ying-Hung Lin. 2016. "SEPT12–NDC1 Complexes Are Required for Mammalian Spermiogenesis" International Journal of Molecular Sciences 17, no. 11: 1911. https://doi.org/10.3390/ijms17111911
APA StyleLai, T.-H., Wu, Y.-Y., Wang, Y.-Y., Chen, M.-F., Wang, P., Chen, T.-M., Wu, Y.-N., Chiang, H.-S., Kuo, P.-L., & Lin, Y.-H. (2016). SEPT12–NDC1 Complexes Are Required for Mammalian Spermiogenesis. International Journal of Molecular Sciences, 17(11), 1911. https://doi.org/10.3390/ijms17111911