
Identification and Evaluation of 21 Novel Microsatellite Markers from the Autumnal Moth (Epirrita autumnata) (Lepidoptera: Geometridae)

 ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
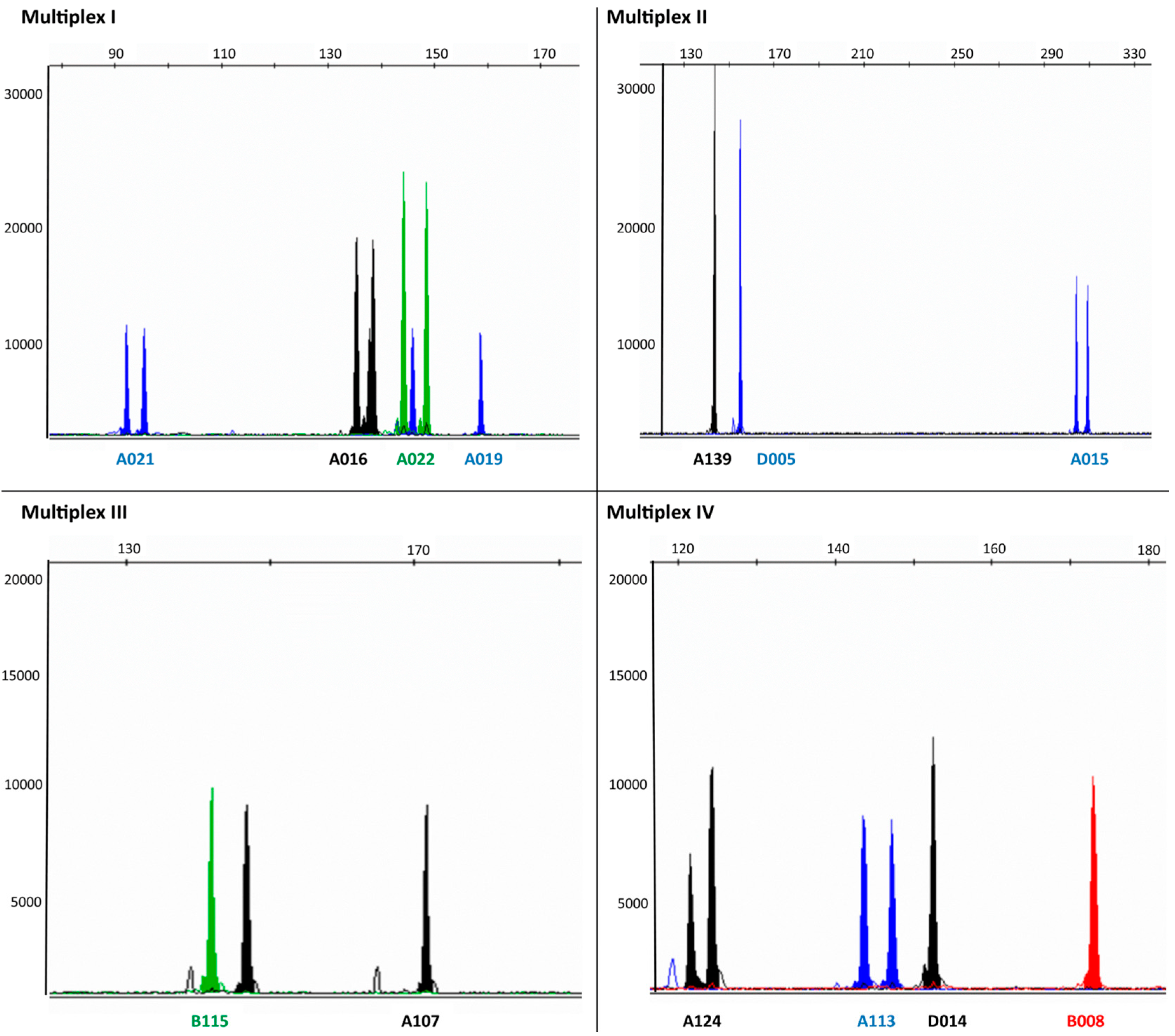

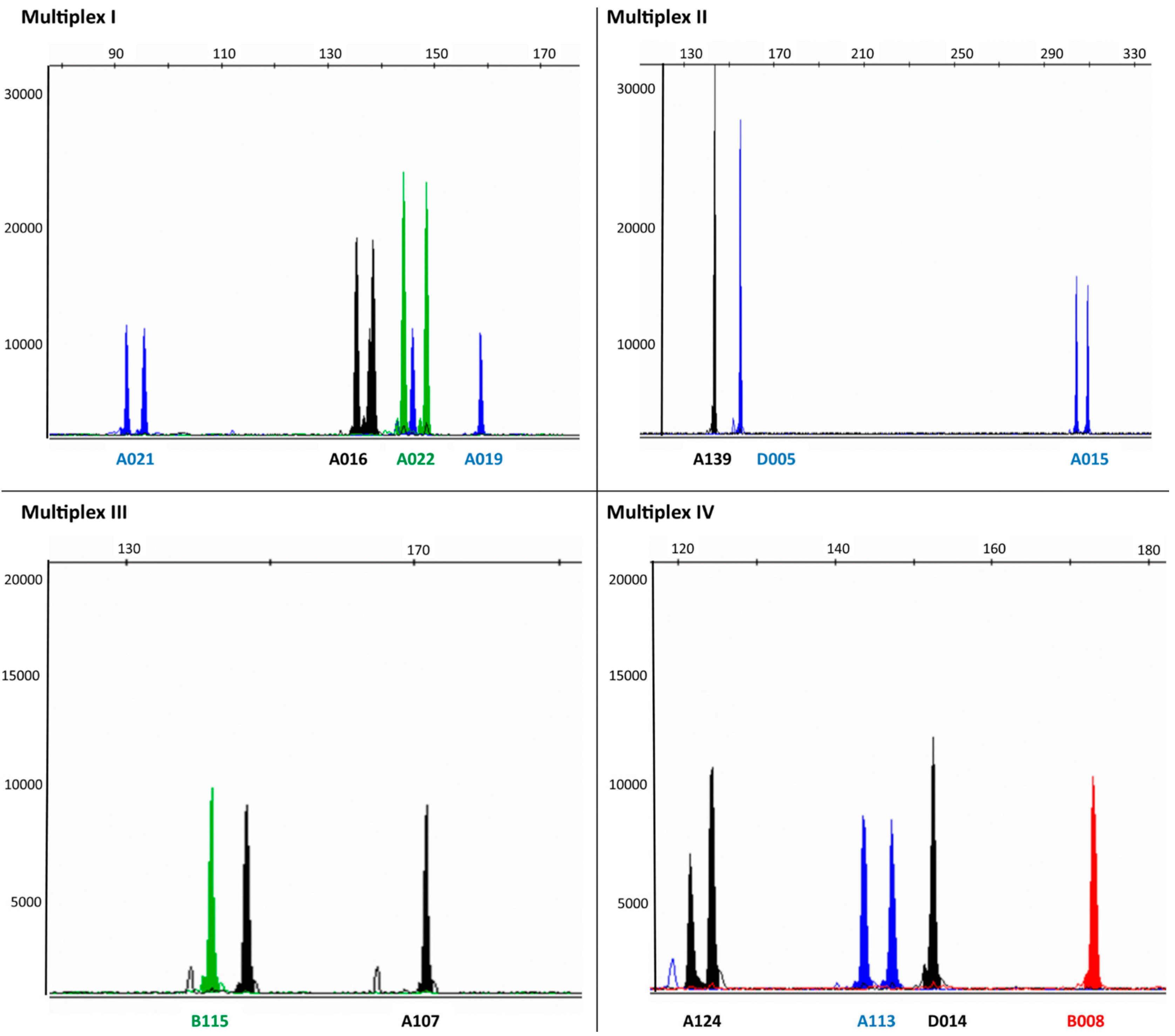
{kind=link}
{kind=link}
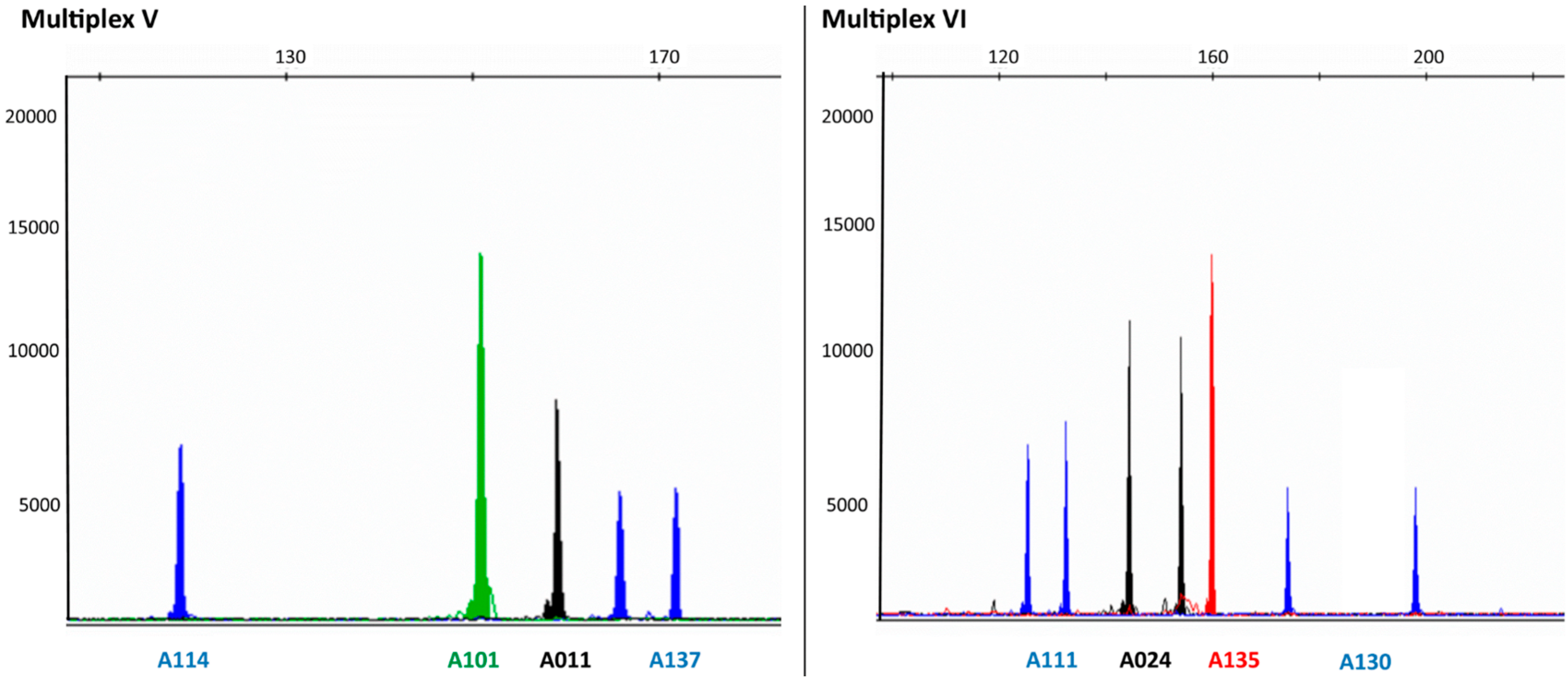
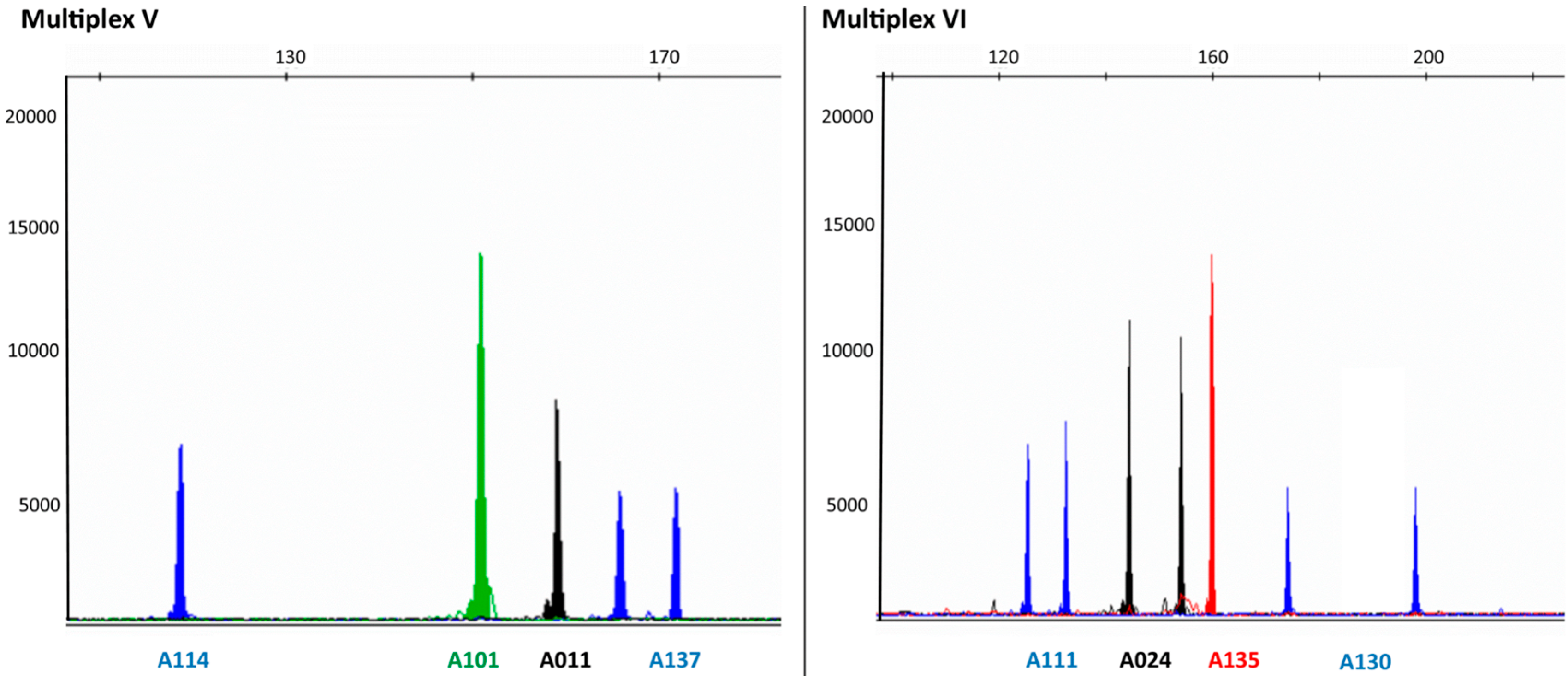{kind=link}
| Multiplex Panel | Locus | Primer Sequences (5ʹ–3ʹ) a | Repeat Motif | Size Range (bp) b | Primer Conc., Dye | GenBank Accession Number |
|---|---|---|---|---|---|---|
| I | A021 | F: CCTAAGAGGGAGGCCCATGT | TGA | 86–95 | 2 µM, FAM | KT428619 |
| R: CAGCTTGGTTCGTTAGCAAGG | ||||||
| A019 | F: GCGTTGGCGCATCTGTAAAT | CAT | 146–173 | 3 µM, FAM | KT428620 | |
| R: CGCCACAGAGGTCGTCAAA | ||||||
| A022 | F: CTGCGTGCTAAAACCTACGGA | CAT | 141–147 | 1 µM, VIC | KT428621 | |
| R: CAGCAGTGGACTTCTTCTGGC | ||||||
| A016 | F: AGACCTACACCTGAGTGCATCTTAGTT | CAT | 135–141 | 3 µM, NED | KT428622 | |
| R: CCATCCCAGGTGTGGTGATT | ||||||
| II | D005 | F: CGGTGGTTGCTATGGGTGTT | CTT | 142–163 | 2 µM, FAM | KT428623 |
| R: TTGCATTCTATGTGGGAGGCT | ||||||
| A015 | F: AATTGTATGCCACCGCTGCT | CAT | 301–325 | 2 µM, FAM | KT428624 | |
| R: TCCGTCTGCCAAGTGTA | ||||||
| A139 | F: ACCTGCGATTACCAATCCGA | GAT | 140–149 | 1 µM, NED | KT428625 | |
| R: TTCCGTGGTTCTTCTTCATCAAC | ||||||
| III | B115 | F: TTATAGGTGTCGGTTAAACACTTTAAAAAC | ACT | 141–148 | 2 µM, VIC | KT428626 |
| R: GGTTAAGGCTGCGCTAAAGCT | ||||||
| A107 | F: TAGGGCCAGCAGTGGACTTC | ATG | 147–201 | 2 µM, NED | KT428627 | |
| R: GTTTCTTTGGATGTCCCTTGGCCTTTA | ||||||
| IV | A124 | F: TGAATCGGTGCTCCAATAGGA | TAA | 115–124 | 3 µM, NED | KT428628 |
| R: GTCTCTGTTGACCCCAGGGA | ||||||
| A113 | F: AGACCTCGTCCAACAGTGGG | GAT | 144–165 | 1 µM, FAM | KT428629 | |
| R: AACATTGGACGATCTTATCGCC | ||||||
| D014 | F: TCGTTTTCATCTATTATTAGTTTAGGATTCA | CAAT | 140–156 | 2 µM, NED | KT428630 | |
| R: TTGTTGCACGCCTTAAATGG | ||||||
| B008 | F: TGCATTGTAATAGGACCTTCATATTTTT | ACAG | 173–178 | 5 µM, PET | KT428631 | |
| R: TTATAGGATCACTTTGTTGTCCGTCT | ||||||
| V | A114 | F: TGTCGAGCTCTACAAAAACTGCA | CAT | 118 | 2 µM, FAM | KT428632 |
| R: AATTGGGCCTCAGGTTTCTGT | ||||||
| A101 | F: GAAGCCGCGCTGTTTCTTAA | GTT | 151 | 1 µM, VIC | KT428633 | |
| R: GAGAGGTCGTCGAAACACCCT | ||||||
| A011 | F: CTAGACCGGAGGCAAACCAA | TGA | 153–159 | 0.5 µM, NED | KT428634 | |
| R: CAAAATGACGGTTTGAGCGA | ||||||
| A137 | F: GATCCAGGATCTGAAGCGGA | CAT | 153–174 | 1 µM, FAM | KT428635 | |
| R: AAGACCGTTCGTCATGGCAT | ||||||
| VI | A111 | F: GGCGGAGGTCTTTTCTAGCAG | ATG | 122–137 | 2 µM, FAM | KT428636 |
| R: AACAAGTTTGGGTTGCAAAAGTTT | ||||||
| A130 | F: AACACACTCGAGGGTCCCAA | CAT | 173–227 | 3 µM, FAM | KT428637 | |
| R: GTTCTAGGGCCAGCAGTGGA | ||||||
| A135 | F: TCCTCCAACTCTTTCCGTGG | CAT | 160–187 | 1 µM, PET | KT428638 | |
| R: TTATGGGTGAGGCTTCGTCC | ||||||
| A024 | F: TCGTCTGTAGATATCAACTGCTGGA | CAT | 145–199 | 1 µM, NED | KT428639 | |
| R: GTGGACGTAAGCAGGCTGGT |
| Locus | Allele/Genotype a | Mean b | S.D. (bp) c |
|---|---|---|---|
| A021 allele A | 92 | 92.24 | 0.07 |
| A021 allele B | 95 | 95.49 | 0.1 |
| A019 allele A | 135 | 135.36 | 0.08 |
| A019 allele B | 138 | 138.47 | 0.08 |
| A022 allele A | 145 | 145.92 | 0.08 |
| A022 allele B | 188 | 188.56 | 0.1 |
| A016 allele A | 144 | 144.20 | 0.1 |
| A016 allele B | 147 | 148.62 | 0.09 |
| D005 allele A | 148 | 148.43 | 0.07 |
| D005 allele B | 154 | 154.99 | 0.03 |
| A015 allele A | 301 | 301.08 | 0.07 |
| A015 allele B | 310 | 309.28 | 0.07 |
| A139 allele A | 143 | 143.53 | 0.07 |
| B115 allele A | 141 | 141.70 | 0.06 |
| A107 allele A | 147 | 146.45 | 0.09 |
| A107 allele B | 201 | 201.98 | 0.05 |
| A124 allele A | 121 | 121.44 | 0.06 |
| A124 allele B | 124 | 124.24 | 0.07 |
| A113 allele A | 147 | 147.06 | 0.08 |
| D014 allele A | 152 | 152.47 | 0.07 |
| B008 allele A | 181 | 181.21 | 0.06 |
| A114 allele A | 118 | 118.48 | 0.06 |
| A101 allele A | 151 | 150.68 | 0.07 |
| A011 allele A | 159 | 158.89 | 0.08 |
| A137 allele A | 165 | 165.68 | 0.09 |
| A137 allele B | 171 | 171.55 | 0.08 |
| A111 allele A | 125 | 125.36 | 0.07 |
| A111 allele B | 131 | 131.64 | 0.07 |
| A130 allele A | 173 | 174.16 | 0.07 |
| A130 allele B | 227 | 227.31 | 0.08 |
| A135 allele A | 160 | 159.92 | 0.08 |
| A024 allele A | 154 | 154.10 | 0.06 |
| A024 allele B | 172 | 172.09 | 0.09 |
| Locus | Allele/Genotype a | Number Repeats | Repeat Structure |
|---|---|---|---|
| A021 allele A | 92 | 6 R | (TGA)6 |
| A021 allele B | 95 | 7 R | (TGA)7 |
| A019 allele A | 146 | – | (CAT)n(AAT)(CAT)n * |
| A022 allele A | 141 * | 5 R | (CAT)5 |
| A022 allele B | 147 * | 6 R | (CAT)6 |
| A016 allele A | 138 | 6 R | (CAT)6 |
| D005 allele A | 148 | 7 R | (CTT)7 |
| D005 allele B | 154 | 9 R | (CTT)9 |
| A015 allele A | 301 | 5 R | (CAT)5 |
| A015 allele B | 322 | 6 R | (CAT)6 |
| A139 allele A | 140 | 7 R | (GAT)7 |
| A139 allele B | 143 | 8 R | (GAT)8 |
| B115 allele A | 141 | 7 R | (ACT)1(TT)(ACT)1(ACA)(ACT)5 |
| B115 allele B | 144 | 8 R | (ACT)1(TT)(ACT)1(ACA)(ACT)6 |
| A107 allele A | 147 | – | – |
| A107 allele B | 201 | 14 R | (ATG)6(AGGCTG)(ATG)3(ACG)(ATG)3(CTG)(ATG)2 |
| A124 allele A | 121 | 7 R | (TAA)7 |
| A124 allele B | 124 | 8 R | (TAA)8 |
| A113 allele A | 147 | 9 R | (GAT)9 |
| A113 allele B | 165 | 15 R | (GAT)15 |
| D014 allele A | 140 | – | (CAAT)n(CAAC)(CAT)n * |
| D014 allele B | 142 | – | – |
| B008 allele A | 173 | 5 R | (ACAG)2(ACAT)(ACAG)3 |
| B008 allele B | 181 | – | – |
| A114 allele A | 118 | 6 R | (CAT)6 |
| A101 allele A | 151 | 8 R | (GTT)3(GT)(GTT)2(T)(GTT)1(GTA)(GTT)2 |
| A011 allele A | 159 | 6 R | (TGA)3(TGT)(TGA)3 |
| A137 allele A | 165 | 7 R | (CAT)7 |
| A137 allele B | 171 | 9R | (CAT)9 |
| A111 allele A | 125 | 10 R | (ATG)10 |
| A111 allele B | 134 | 7 R | (ATG)7 |
| A130 allele A | 173 | 6 R | (CAT)6 |
| A130 allele B | 227 | 19 R | (CAT)7(CAGCCT)(CAT)6(CAGCCTCAC)(CAT)6 |
| A135 allele A | 160 | 10 R | (CAT)10 |
| A024 allele A | 145 | 6 R | (CAT)6 |
| A024 allele B | 187 | 17 R | CTGAT(CAT)2(CACA)(CAT)15 |
| Locus | NA | HO | FIS | HE | PI | FNULL | HWE p Values a |
|---|---|---|---|---|---|---|---|
| A021 | 3 | 0.438 | 0.1258 | 0.495 | 0.32 | 0.0249 | 0.2676 |
| A019 | 3 | 0.553 | −0.1423 | 0.479 | 0.32 | −0.0496 | 0.9699 |
| A022 | 3 | 0.4375 | 0.2654 | 0.5877 | 0.24 | 0.0918 | 0.0054 ** |
| A016 | 3 | 0.1667 | −0.0697 | 0.1543 | 0.72 | −0.0111 | 1.0000 |
| D005 | 7 | 0.6042 | 0.0347 | 0.6191 | 0.19 | 0.0091 | 0.6835 |
| A015 | 5 | 0.4583 | 0.3233 | 0.6680 | 0.17 | 0.1322 | 0.0001 ** |
| A139 | 3 | 0.229 | 0.1009 | 0.252 | 0.59 | 0.0178 | 0.2429 |
| B115 | 5 | 0.354 | 0.2766 | 0.483 | 0.34 | 0.0913 | 0.0420 * |
| A107 | 8 | 0.596 | 0.1796 | 0.717 | 0.12 | 0.0571 | 0.0341* |
| A124 | 4 | 0.479 | −0.0810 | 0.439 | 0.39 | −0.0225 | 0.7396 |
| A113 | 7 | 0.158 | 0.7739 | 0.682 | 0.14 | 0.3127 | 0.0000 ** |
| D014 | 6 | 0.106 | 0.4148 | 0.179 | 0.68 | 0.0625 | 0.0026 * |
| B008 | 2 | 0.08 | 0.6336 | 0.211 | 0.64 | 0.1083 | 0.0194 * |
| A114 | 1 | 0 | 0 | 0 | 1.00 | 0 | No |
| A101 | 1 | 0 | 0 | 0 | 1.00 | 0 | No |
| A011 | 2 | 0.041 | −0.0108 | 0.041 | 0.92 | −0.0009 | 1.0000 |
| A137 | 5 | 0.688 | 0.0061 | 0.645 | 0.16 | 0.0034 | 0.5214 |
| A111 | 8 | 0.404 | 0.4519 | 0.726 | 0.12 | 0.1722 | 0.0000 ** |
| A130 | 7 | 0.568 | 0.1975 | 0.698 | 0.14 | 0.055 | 0.0603 |
| A135 | 2 | 0.255 | −0.1231 | 0.225 | 0.62 | −0.0246 | 1.0000 |
| A024 | 13 | 0.614 | 0.2379 | 0.794 | 0.06 | 0.0833 | 0.0014 * |
3. Materials and Methods
3.1. Sampling and Materials
3.2. Identification of Microsatellites Markers
3.3. DNA Extraction
3.4. Development of PCR Assays
3.5. Testing for PCR Specificity, Sensitivity and Precision
3.5.1. DNA Sequencing
3.5.2. Analysis of Data
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kallio, P.; Lehtonen, J. On the plasmatic template system in Micrasterias morphogenesis. Ann. Acad. Sci. Fenn. Biol. 1973, 199, 1–6. [Google Scholar] [PubMed]
- Tenow, O.; Bylund, H. Recovery of a Betula pubescens forest in northern Sweden after severe defoliation by Epirrita autumnata. J. Veg. Sci. 2000, 11, 855–862. [Google Scholar] [CrossRef]
- Karlsen, S.R.; Jepsen, J.U.; Odland, A.; Ims, R.A.; Elvebakk, A. Outbreaks by canopy-feeding geometrid moth cause state-dependent shifts in understorey plant communities. Oecologia 2013, 173, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, J.U.; Biuw, M.; Ims, R.A.; Kapari, L.; Schott, T.; Vindstad, O.P.L.; Hagen, S.B. Ecosystem impacts of a range expanding forest defoliator at the forest-tundra ecotone. Ecosystems 2013, 16, 561–575. [Google Scholar] [CrossRef]
- Vindstad, O.P.L.; Schultze, S.; Jepsen, J.U.; Biuw, M.; Kapari, L.; Sverdrup-Thygeson, A.; Ims, R.A. Numerical responses of saproxylic beetles to rapid increases in dead wood availability following geometrid moth outbreaks in sub-arctic mountain birch forest. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, E.; Forchhammer, M.C.; Bret-Harte, M.S.; Callaghan, T.V.; Christensen, T.R.; Elberling, B.; Fox, A.D.; Gilg, O.; Hik, D.S.; Hoye, T.T.; et al. Ecological dynamics across the arctic associated with recent climate change. Science 2009, 325, 1355–1358. [Google Scholar] [CrossRef]
- Hagen, S.B.; Jepsen, J.U.; Schott, T.; Ims, R.A. Spatially mismatched trophic dynamics: Cyclically outbreaking geometrids and their larval parasitoids. Biol. Lett. 2010, 6, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Hagen, S.B.; Jepsen, J.U.; Yoccoz, N.G.; Ims, R.A. Anisotropic patterned population synchrony in climatic gradients indicates nonlinear climatic forcing. Proc. R. Soc. B Biol. Sci. 2008, 275, 1509–1515. [Google Scholar] [CrossRef] [PubMed]
- Ims, R.A.; Yoccoz, N.G.; Hagen, S.B. Do sub-Arctic winter moth populations in coastal birch forest exhibit spatially synchronous dynamics? J. Anim. Ecol. 2004, 73, 1129–1136. [Google Scholar] [CrossRef]
- Hagen, S.B.; Jepsen, J.U.; Ims, R.A.; Yoccoz, N.G. Shifting altitudinal distribution of outbreak zones of winter moth Operophtera brumata in sub-arctic birch forest: A response to recent climate warming? Ecography 2007, 30, 299–307. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Kapari, L.; Hagen, S.B.; Schott, T.; Vindstad, O.P.L.; Nilssen, A.C.; Ims, R.A. Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Glob. Chang. Biol. 2011, 17, 2071–2083. [Google Scholar] [CrossRef]
- Jepsen, J.U.; Hagen, S.B.; Ims, R.A.; Yoccoz, N.G. Climate change and outbreaks of the geometrids Operophtera brumata and Epirrita autumnata in subarctic birch forest: Evidence of a recent outbreak range expansion. J. Anim. Ecol. 2008, 77, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Meglecz, E.; Solignac, M. Microsatellite loci for Parnassius mnemosyne (Lepidoptera). Hereditas 1998, 128, 179–180. [Google Scholar] [CrossRef]
- Meglecz, E.; Petenian, F.; Danchin, E.; Coeur d’Acier, A.; Rasplus, J.Y.; Faure, E. High similarity between flanking regions of different microsatellites detected within each of two species of Lepidoptera: Parnassius apollo and Euphydryas aurinia. Mol. Ecol. 2004, 13, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.X. Lepidopteran microsatellite DNA: Redundant but promising. Trends Ecol. Evol. 2004, 19, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Sarhan, A. Isolation and characterization of five microsatellite loci in the Glanville fritillary butterfly (Melitaea cinxia). Mol. Ecol. Notes 2006, 6, 163–164. [Google Scholar] [CrossRef]
- Hundsdoerfer, A.K.; Sanetra, M.; Corbeil, D.; Stuckas, H. Eleven hawkmoth microsatellite loci of Canary Island Hyles tithymali (Lepidoptera). Conserv. Genet. Resour. 2010, 2, 241–244. [Google Scholar] [CrossRef]
- Sinama, M.; Dubut, V.; Costedoat, C.; Gilles, A.; Junker, M.; Malausa, T.; Martin, J.-F.; Neve, G.; Pech, N.; Schmitt, T.; et al. Challenges of microsatellite development in Lepidoptera: Euphydryas aurinia (Nymphalidae) as a case study. Eur. J. Entomol. 2011, 108, 261–266. [Google Scholar] [CrossRef]
- Massardo, D.; Roratto, P.A.; Vargas, H.A.; Kronforst, M.R.; Moreira, G.R.P. Development of a microsatellite library for the passion flower butterfly Dione moneta Hubner (Lepidoptera: Nymphalidae: Heliconiinae). Conserv. Genet. Resour. 2012, 4, 719–724. [Google Scholar] [CrossRef]
- Weibin, J.; Jianqing, Z.; Ling, Z.; Minghai, C.; Chao, S.; Weidong, Y. Isolation and characterization of microsatellite loci in Polytremis nascens (Lepidoptera: Hesperiidae) and their cross-amplification in related species. Appl. Entomol. Zool. 2014, 49, 177–181. [Google Scholar] [CrossRef]
- Mira, O.; Martinez, J.G.; Dawson, D.A.; Tinaut, A.; Sanchez-Prieto, C. Twenty new microsatellite loci for population structure and parentage studies of Parnassius apollo nevadensis (Lepidoptera; Papilionidae). J. Insect Conserv. 2014, 18, 771–779. [Google Scholar] [CrossRef]
- Zima, J., Jr.; Lestina, D.; Jansta, P.; Petru, V.; Tropek, R. Isolation and characterisation of microsatellite markers for Mylothris jacksoni knutsoni (Lepidoptera: Pieridae), an endemic butterfly of the Gulf of Guinea Highlands. Conserv. Genet. Resour. 2014, 6, 763–764. [Google Scholar] [CrossRef]
- Behura, S.K.; Severson, D.W. Motif mismatches in microsatellites: Insights from genome-wide investigation among 20 insect species. DNA Res. 2015, 22, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Mende, M.B.; Stuckas, H.; Hundsdoerfer, A.K. Eight new microsatellite loci of the Western Palearctic Hyles euphorbiae complex (Lepidoptera, Sphingidae). Ann. Zool. Fenn. 2011, 48, 142–146. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Weetman, D.; Hutchinson, W.F. Estimation and adjustment of microsatellite null alleles in nonequilibrium populations. Mol. Ecol. Notes 2006, 6, 255–256. [Google Scholar] [CrossRef]
- Dakin, E.E.; Avise, J.C. Microsatellite null alleles in parentage analysis. Heredity 2004, 93, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, H.; Kwon, D.H.; Lee, S. Isolation and characterization of microsatellite loci from Spodoptera exigua (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2012, 47, 149–152. [Google Scholar] [CrossRef]
- Sauné, L.; Abella, F.; Kerdelhué, C. Isolation, characterization and PCR multiplexing of 17 microsatellite loci in the pine processionary moth Thaumetopoea pityocampa (Lepidoptera, Notodontidae). Conserv. Genet. Resour. 2015, 7, 755–757. [Google Scholar] [CrossRef]
- GENETIX, version 4.05: Logiciel sous Windows pour la Génétique des Populations, Laboratoire Génome, Populations, Interactions, CNRS UMR 5171, Université de Montpellier II: Montpellier, France, 1996–2004.
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aarnes, S.G.; Fløystad, I.; Schregel, J.; Vindstad, O.P.L.; Jepsen, J.U.; Eiken, H.G.; Ims, R.A.; Hagen, S.B. Identification and Evaluation of 21 Novel Microsatellite Markers from the Autumnal Moth (Epirrita autumnata) (Lepidoptera: Geometridae). Int. J. Mol. Sci. 2015, 16, 22541-22554. https://doi.org/10.3390/ijms160922541
Aarnes SG, Fløystad I, Schregel J, Vindstad OPL, Jepsen JU, Eiken HG, Ims RA, Hagen SB. Identification and Evaluation of 21 Novel Microsatellite Markers from the Autumnal Moth (Epirrita autumnata) (Lepidoptera: Geometridae). International Journal of Molecular Sciences. 2015; 16(9):22541-22554. https://doi.org/10.3390/ijms160922541
Chicago/Turabian StyleAarnes, Siv Grethe, Ida Fløystad, Julia Schregel, Ole Petter Laksforsmo Vindstad, Jane Uhd Jepsen, Hans Geir Eiken, Rolf A. Ims, and Snorre B. Hagen. 2015. "Identification and Evaluation of 21 Novel Microsatellite Markers from the Autumnal Moth (Epirrita autumnata) (Lepidoptera: Geometridae)" International Journal of Molecular Sciences 16, no. 9: 22541-22554. https://doi.org/10.3390/ijms160922541
APA StyleAarnes, S. G., Fløystad, I., Schregel, J., Vindstad, O. P. L., Jepsen, J. U., Eiken, H. G., Ims, R. A., & Hagen, S. B. (2015). Identification and Evaluation of 21 Novel Microsatellite Markers from the Autumnal Moth (Epirrita autumnata) (Lepidoptera: Geometridae). International Journal of Molecular Sciences, 16(9), 22541-22554. https://doi.org/10.3390/ijms160922541