
Detection of miR-33 Expression and the Verification of Its Target Genes in the Fatty Liver of Geese

Abstract
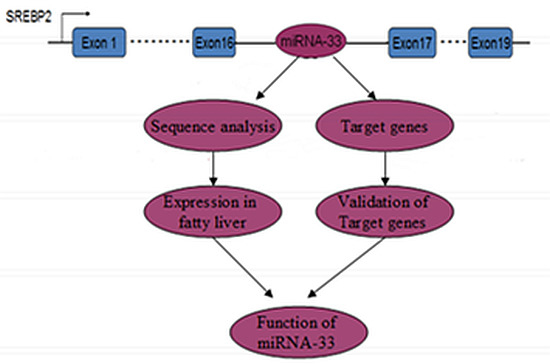
:
1. Introduction
2. Results
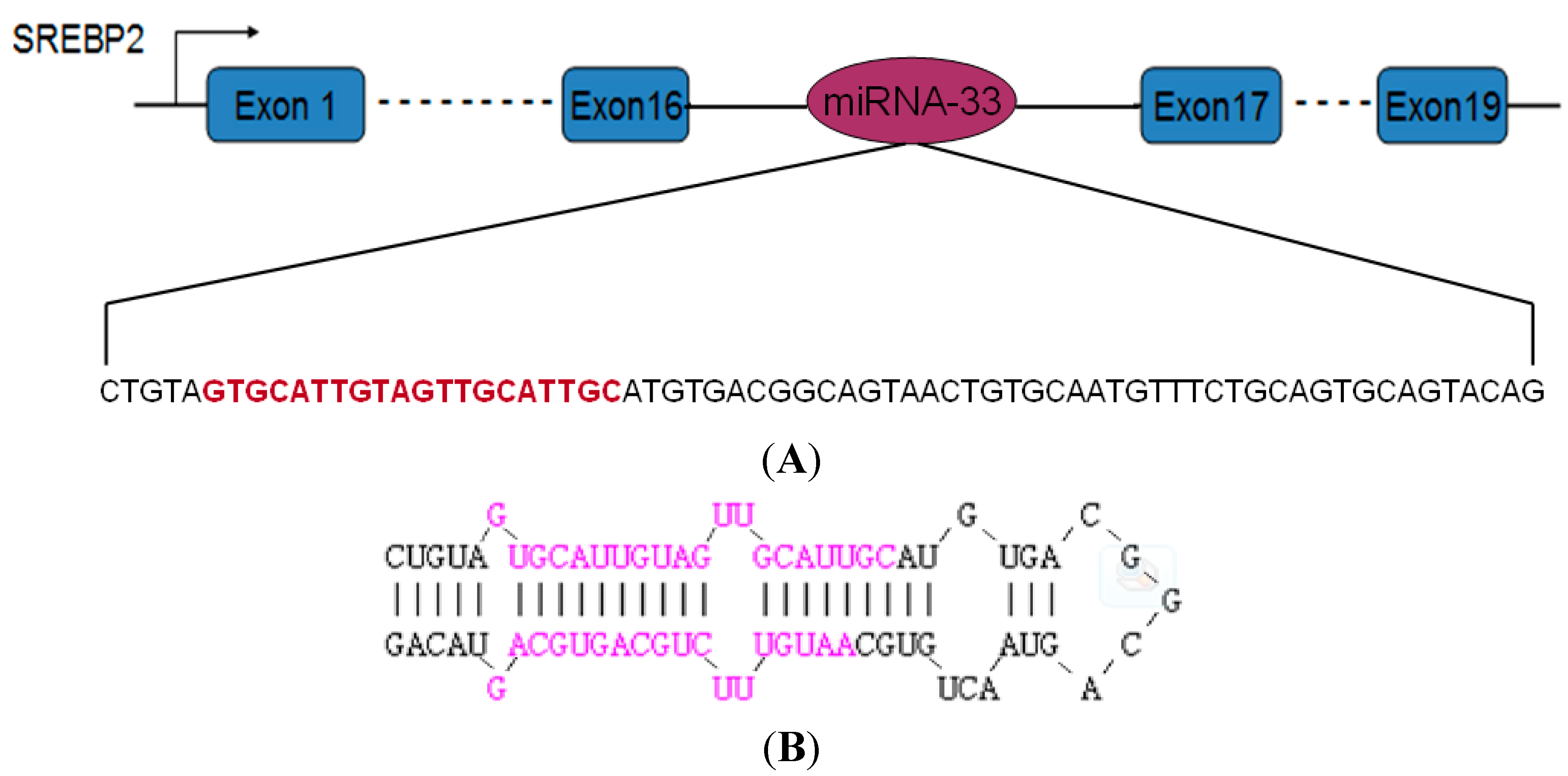
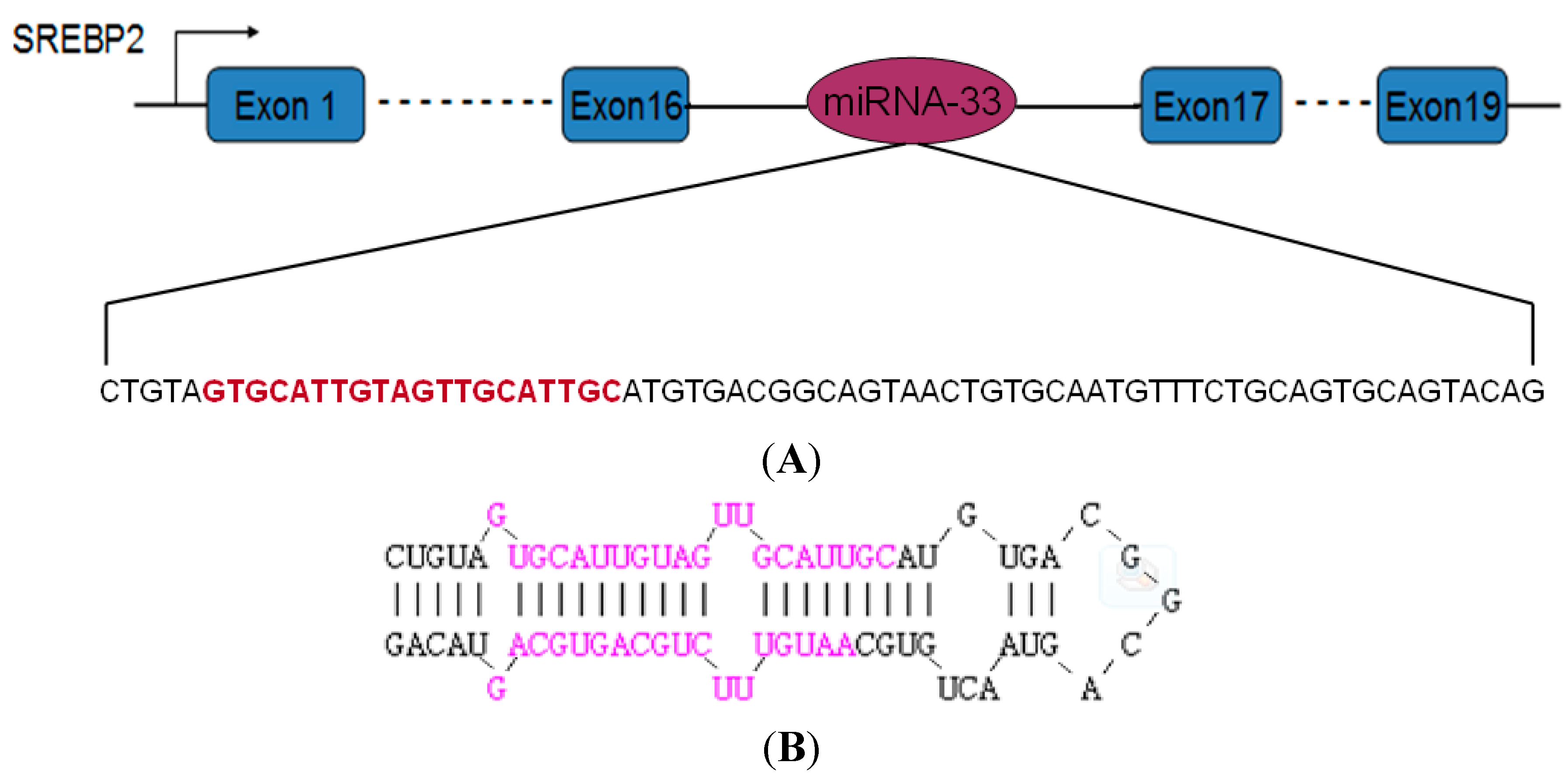
2.1. Precursor Sequence of the miRNA-33 of Landes Geese
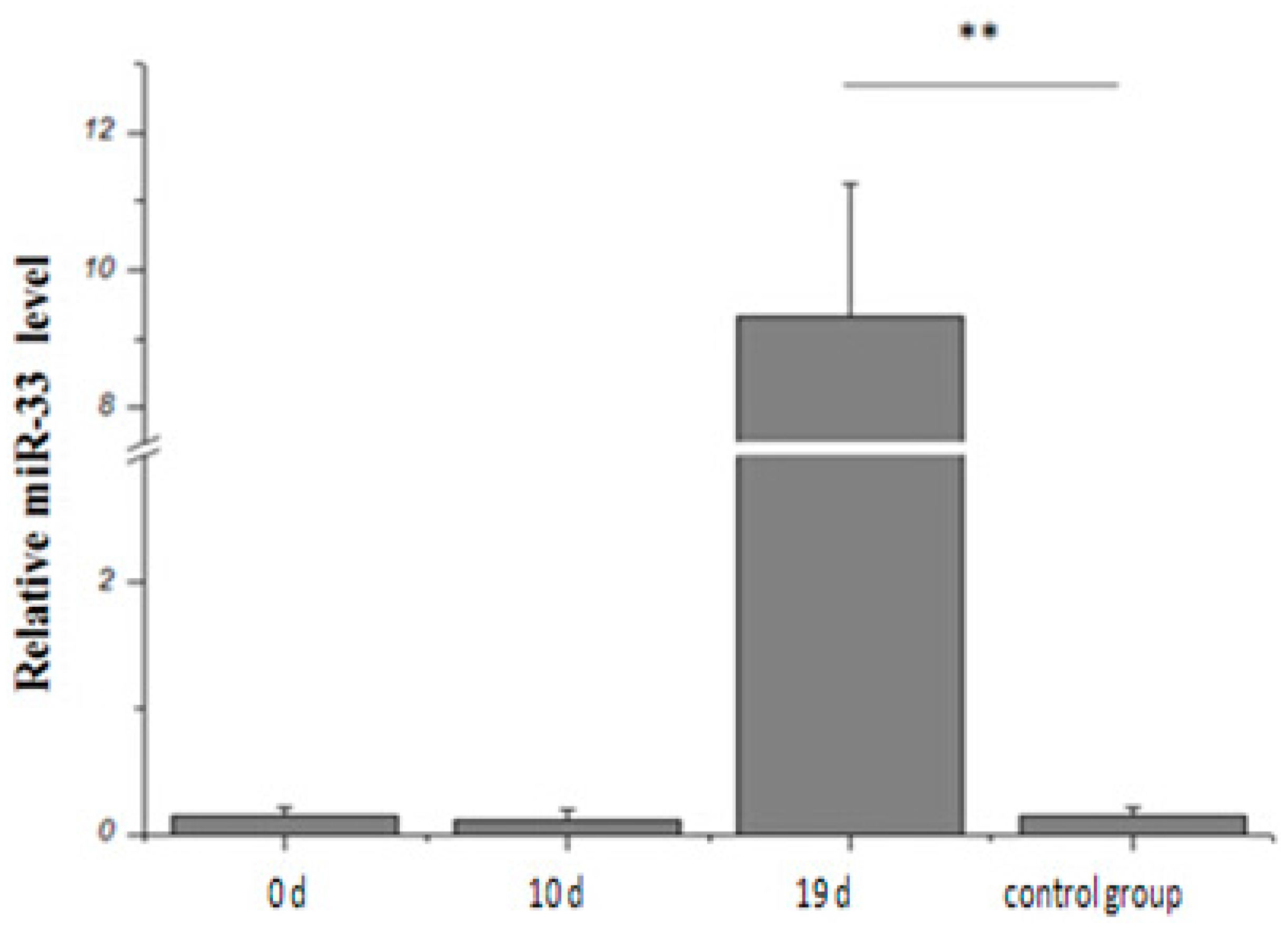
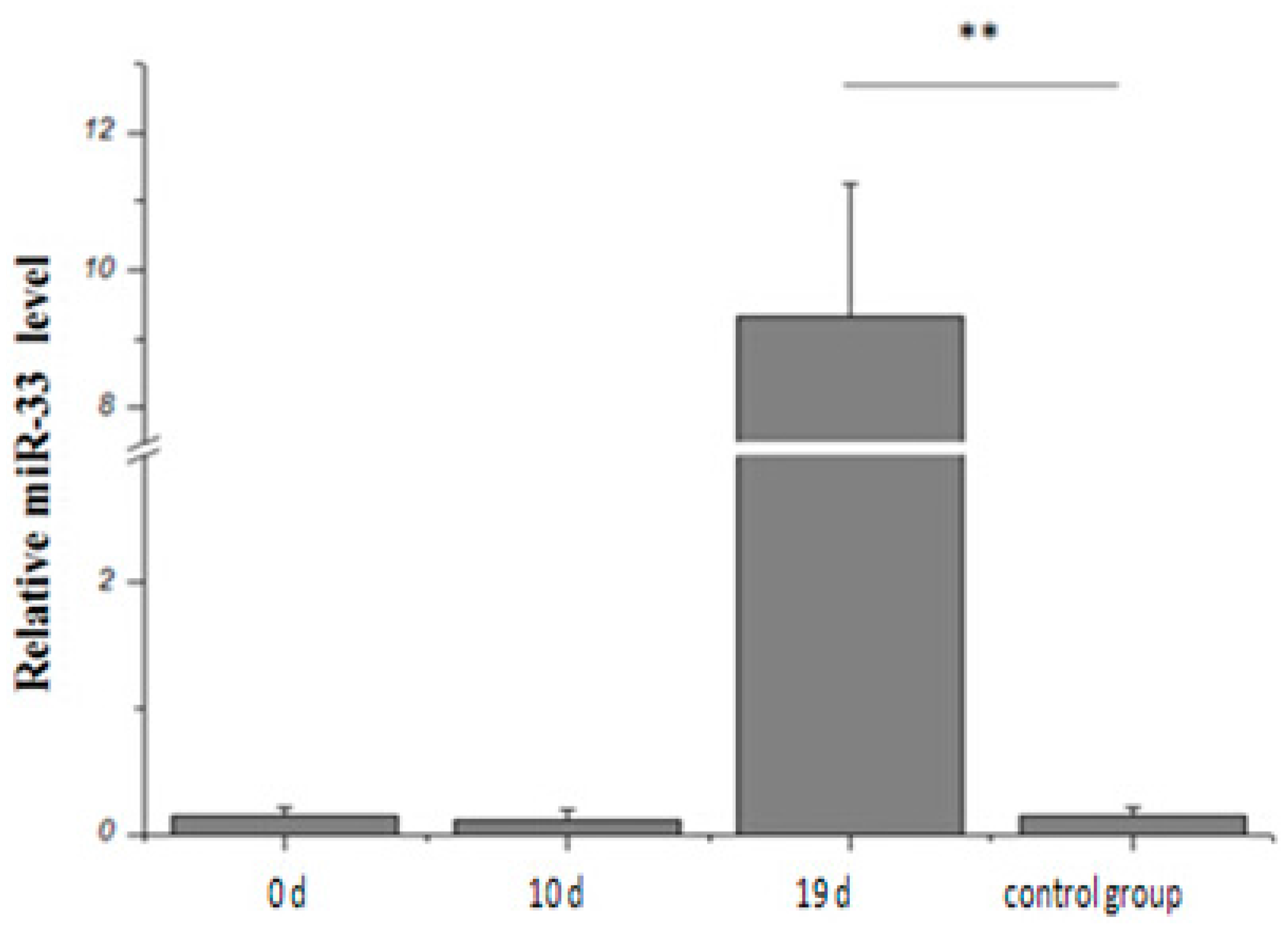
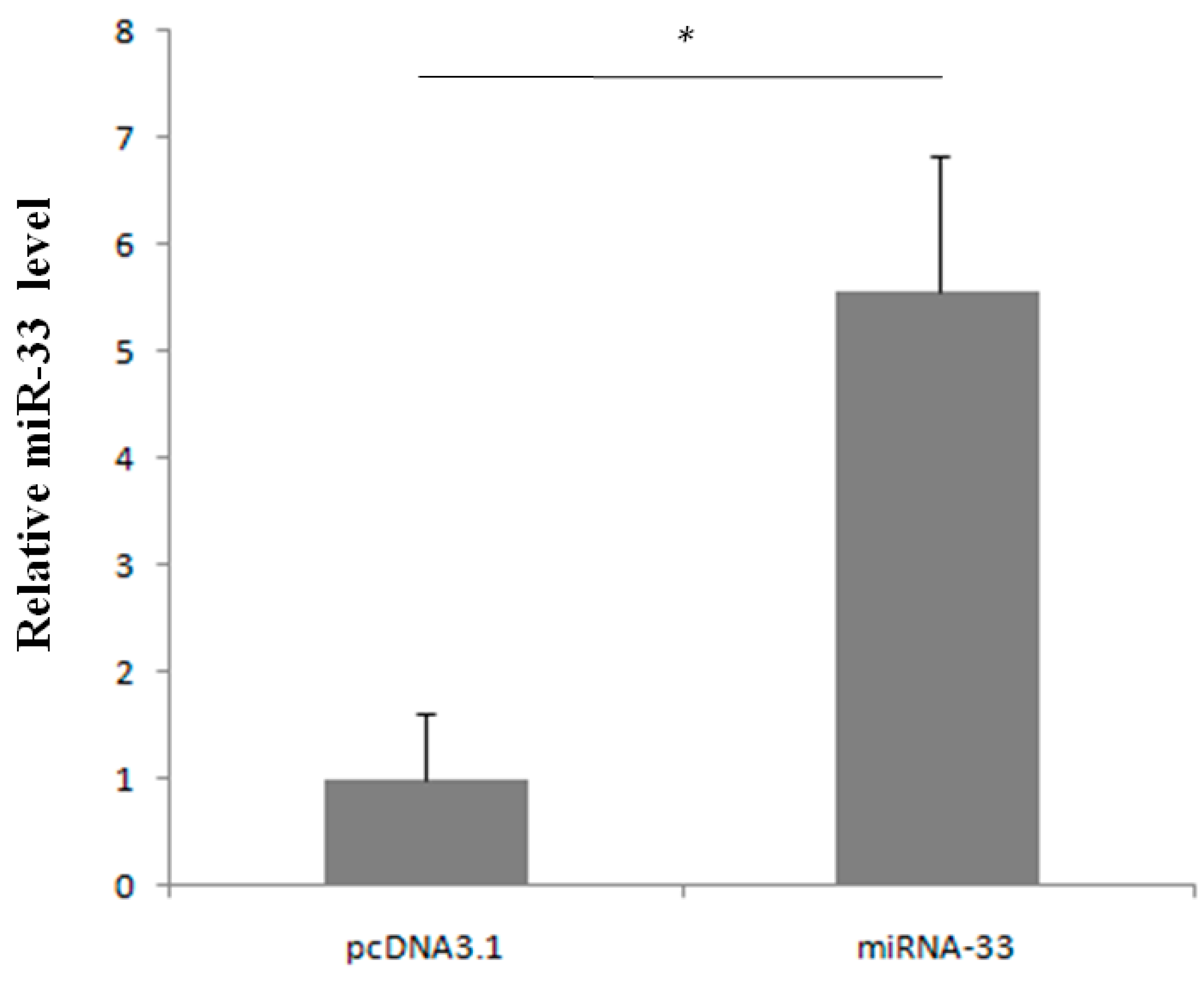
2.2. Expression Rule of miR-33 in Goose Fatty Liver
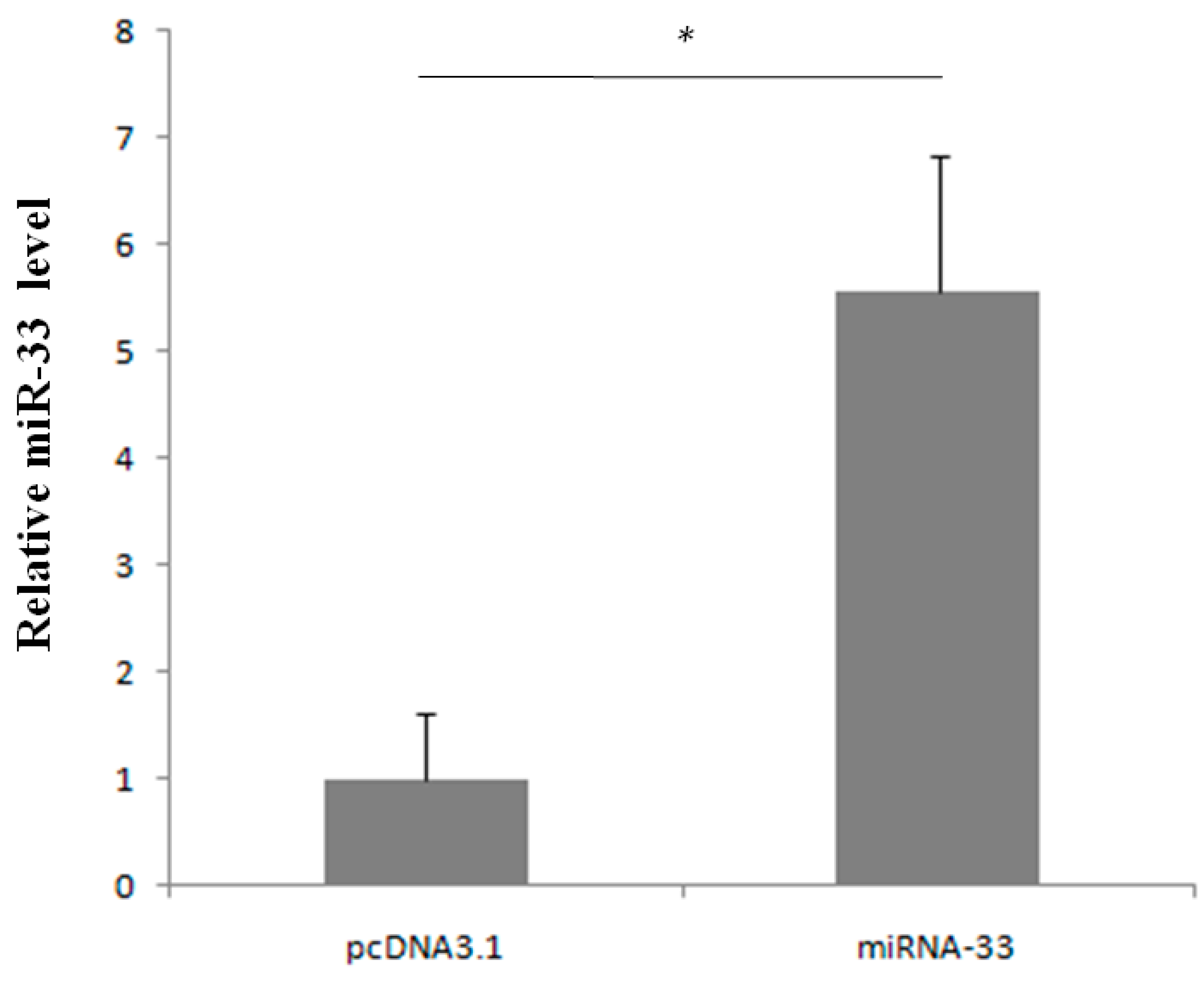
2.3. Prediction of miR-33 Target Genes
2.4. Amplification of the Target Sequence and Vector Construction


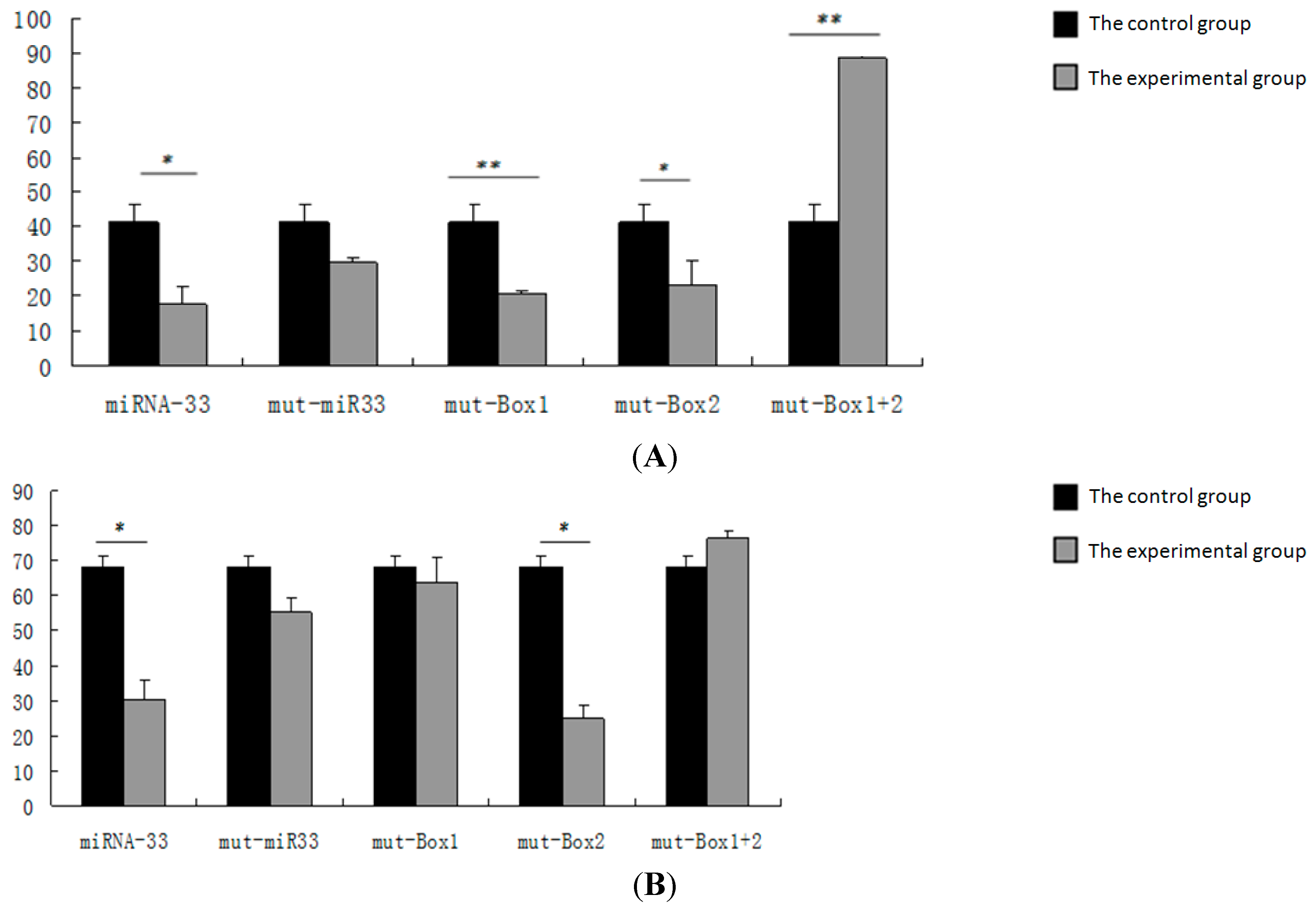
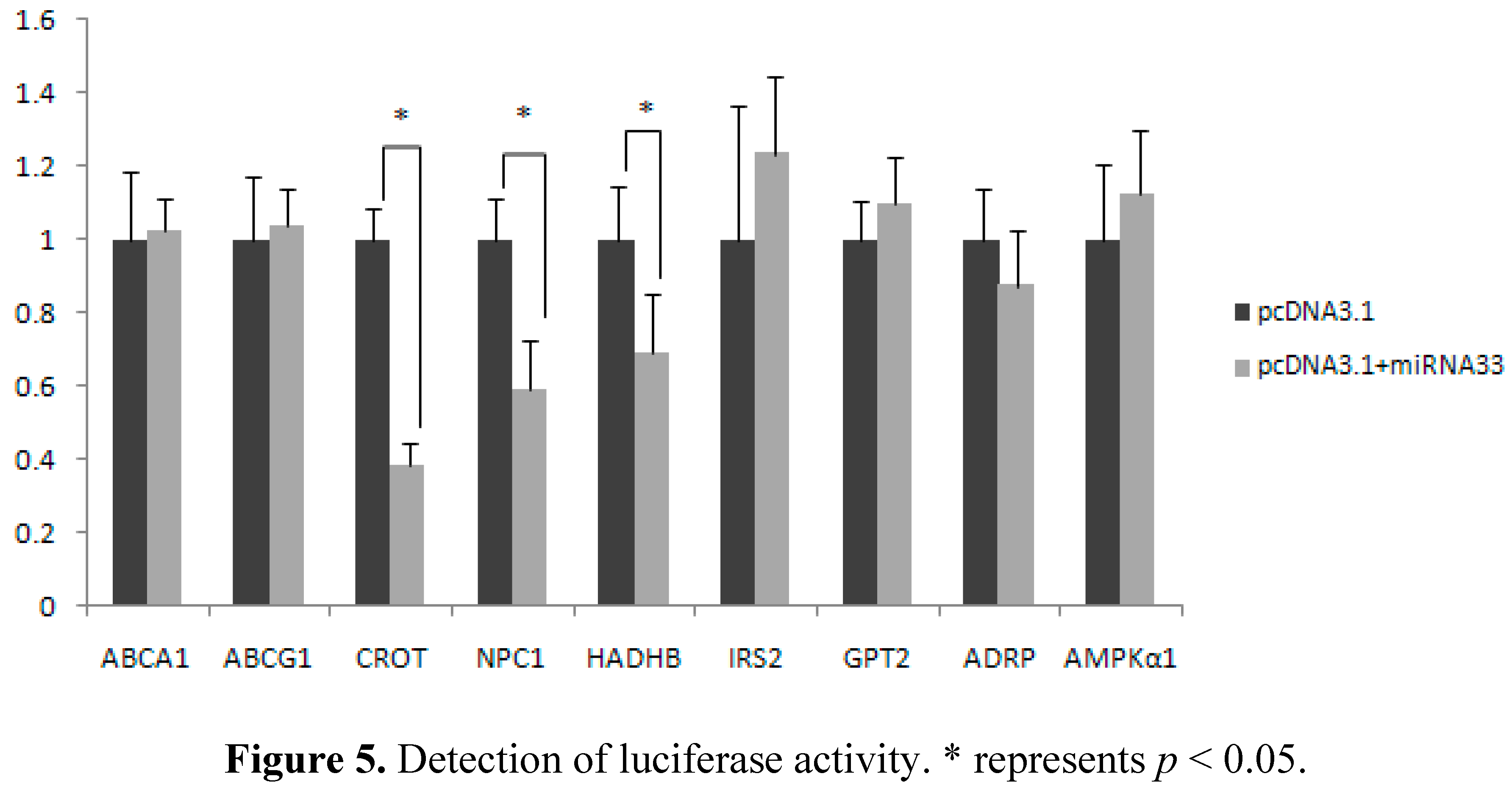
2.5. Verification of miR-33 Target Gene

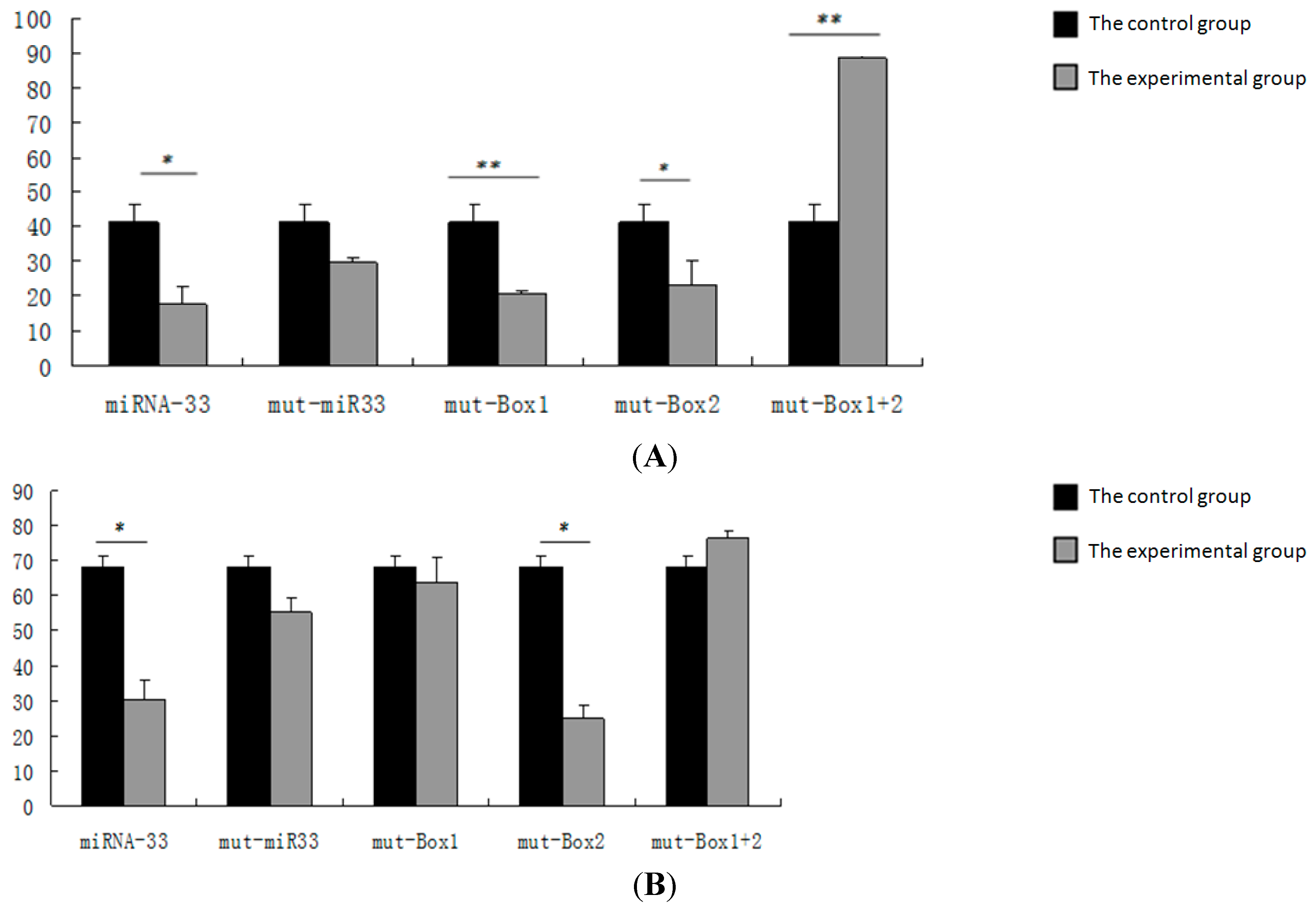
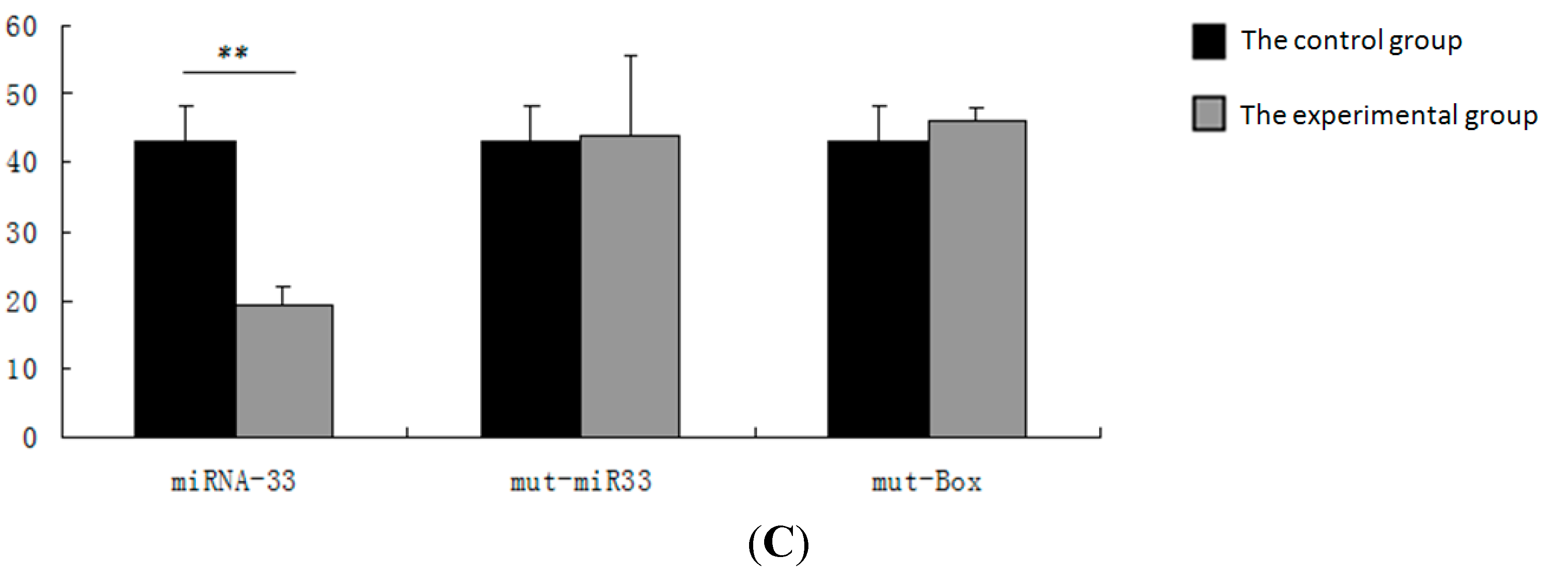
2.6. Verification of the miR-33 Target Site

3. Discussion
3.1. Analysis of miRNA-33 Expression in the Fatty Liver of Geese
3.2. Prediction of the miRNA-33 Target Gene in Landes Geese
3.3. Verification of the miRNA-33 Target Gene
3.4. Verification of the miRNA-33 Target Site
4. Experimental Section
4.1. Experimental Animals and Breeding Management
4.2. Prediction of Target Genes
4.3. Primers of Carrier Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application | Primers (5′→3′) | Length of the Product (bp) |
|---|---|---|
| ABCA1 target site | F cgagctcGCCAATTTCAGCCAAGAAGTGA | 70 |
| R cccaagcttCTTTGGGAGTAACCTATCCCCAG | ||
| ABCG1 target site | F cgagctcAGGAAGAAGAAATAGAAGGGAA | 267 |
| R cccaagcttACAGAAAACCACAAAGATGAAA | ||
| NPC1 target site | F cgagctcCTGGACTGCTCAACCACTGAC | 211 |
| R cccaagcttGCCTCTCCCATTGGAATGTA | ||
| NPC1 target site | F AGAGACAAAAATTGCATCAACCTGCATTTA | 211 |
| R GCAATTTTTGTCTCTATTTTTAGGGGGG | ||
| CROT target site | F cgagctcATTTGCAACAGCAATGCAAG | 197 |
| R cccaagcttAGTGCTCCACTGGCAAAAAC | ||
| CROT target site 1 mutation | F ATCTCCCAAGTATGTTTGCGCTGTTGAGGCA | 197 |
| R GCAAACATACTTGGGAGATATGGTGTTG | ||
| CROT target site 2 mutation | F CCCAAGCTTAGTGCTCCACTGGCAAAAAC | 197 |
| RCGAGCTCATTTGCAACAGCAGCGCAAGTAGTA | ||
| HADHB target site | F cgagctcATGGGGGGACTGCTGAAGGAGT | 256 |
| R cccaagcttGAGATTAGTGTGGTTACGACGA | ||
| HADHB target site 1 mutation | F TGTTTTCATTAGTGCGCTGAAATGGCATTGCC | 256 |
| R GCACTAATGAAAACATACATACAGTCCT | ||
| HADHB target site 2 mutation | F TGCATTGAAATGGCGCTGCCAGGCACAGGA | 256 |
| R TCCTGTGCCTGGCAGCGCCATTTCAATGCA | ||
| ADRP target site | F cgagctcGGCTGCTGACTTGGTAGGAG | 415 |
| R cgacgcgtCACAACCAGGCATTGCTCTA | ||
| IRS2 target site | F cgagctcGCCCAACTCATGTCCTGTCA | 358 |
| R cccaagcttAGTTCAGTAAGGCTGGCGAC | ||
| GPT2 target site | F cgagctcACAGCAGACAGGGAACACTT | 223 |
| R cgacgcgtATCTGCAAGTCGAAAGCCAG | ||
| AMPKα1 target site | F cgagctcAACAAAGGCGCTGAAAAAACTA | 321 |
| R cccaagcttCTGAATAAAGGGGGAAGGAACA | ||
| miR-33 overexpression | F ggaattcCCTAAAGCTGGAGCCTTCCT | 203 |
| R ccgctcgagCGGCTCGCTATTTTAGTTGC | ||
| miR-33 point mutation 1 | F GTGCATTGTAGTTGCGCTGCATGTGACGGCA | 203 |
| R GCAACTACAATGCACTACAGCTGCCACC | ||
| miR-33 point mutation 2 | F AGTGCATTGTAGTTGCGCAACATGTGACGG | 203 |
| R GCGCAACTACAATGCACTACAGCTGCCA |
4.4. miRNA-33 Real-Time Reverse Transcription Polymerase Chain Reaction
4.5. Target in Vitro Assay
4.5.1. Vector Construction
4.5.2. Point Mutation
4.5.3. Dual-Luciferase Reporter Assay
4.6. CHO Culture and Transfection
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Doench, J.G.; Sharp, P.A. Specificity of microRNA target selection in translational repression. Genes Dev. 2004, 18, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar] [CrossRef]
- Horton, J.D. Sterol regulatory element-binding proteins: Transcriptional activators of lipid synthesis. Biochem. Soc. Trans. 2002, 30, 1091. [Google Scholar] [CrossRef] [PubMed]
- Osborne, T.F. Sterol regulatory element-binding proteins (SREBPs): Key regulators of nutritional homeostasis and insulin action. J. Biol. Chem. 2000, 275, 32379–32382. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Ono, K.; Horiguchi, M.; Nishi, H.; Nakamura, T.; Nagao, K.; Kinoshita, M.; Kuwabara, Y.; Marusawa, H.; Iwanaga, Y.; et al. MicroRNA-33 encoded by an intron of sterol regulatory element-binding protein 2 (Srebp2) regulates HDL in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 17321–17326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dávalos, A.; Goedeke, L.; Smibert, P.; Ramírez, C.M.; Warrier, N.P.; Andreo, U.; Cirera-Salinas, D.; Rayner, K.; Suresh, U.; Pastor-Pareja, J.C.; et al. miR-33a/b contribute to the regulation of fatty acid metabolism and insulin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 9232–9237. [Google Scholar] [CrossRef] [PubMed]
- Gerin, I.; Clerbaux, L.A.; Haumont, O.; Lanthier, N.; Das, A.K.; Burant, C.F.; Leclercq, I.A.; MacDougald, O.A.; Bommer, G.T. Expression of miR-33 from an SREBP2 intron inhibits cholesterol export and fatty acid oxidation. J. Biol. Chem. 2010, 285, 33652–33661. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.J.; Suárez, Y.; Dávalos, A.; Parathath, S.; Fitzgerald, M.L.; Tamehiro, N.; Fisher, E.A.; Moore, K.J.; Fernández-Hernando, C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science 2010, 328, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R. Cholesterol efflux pathways and other potential mechanisms involved in the athero-protective effect of high density lipoproteins. J. Intern. Med. 2008, 263, 256–273. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R.; Yvan-Charvet, L.; Terasaka, N.; Pagler, T.; Wang, N. HDL, ABC transporters, and cholesterol efflux: Implications for the treatment of atherosclerosis. Cell Metab. 2008, 7, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.C.; Hsu, S.D.; Hsu, C.S.; Lai, T.C.; Chen, S.J.; Shen, R.; Huang, Y.; Chen, H.C.; Lee, C.H.; Tsai, T.F.; et al. MicroRNA-122 plays a critical role in liver homeostasis and hepatocarcinogenesis. J. Clin. Investig. 2012, 122, 2773–2897. [Google Scholar] [CrossRef] [PubMed]
- Davail, S.; Guy, G.; André, J.M.; Hermier, D.; Hoo-Paris, R. Metabolism in two breeds of geese with moderate or large overfeeding induced liver-steatosis. Comp. Biochem. Phys. A 2000, 126, 91–99. [Google Scholar] [CrossRef]
- Mourot, J.; Guy, G.; Lagarrigue, S.; Peiniau, P.; Hermier, D. Role of hepatic lipogenesis in the susceptibility to fatty liver in the goose (Anser anser). Comp. Biochem. Phys. B 2000, 126, 81–87. [Google Scholar] [CrossRef]
- Adams, L.A.; Lymp, J.F.; St Sauver, J.; Sanderson, S.O.; Lindor, K.D.; Feldstein, A.; Angulo, P. The natural history of nonalcoholic fatty liver disease: A population-based cohort study. Gastroenterology 2005, 129, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Mourot, J.; Guy, G.; Peiniau, P.; Hermier, D. Effects of overfeeding on lipid synthesis, transport and storage in two breeds of geese differing in their capacity for fatty liver production. Anim. Res. 2006, 55, 427–442. [Google Scholar] [CrossRef]
- Han, C.; Wan, H.; Ma, S.; Liu, D.; He, F.; Wang, J.; Pan, Z.; Liu, H.; Li, L.; He, H.; et al. Role of mammalian sirtuin 1 (SIRT1) in lipids metabolism and cell proliferation of goose primary hepatocytes. Mol. Cell. Endocrinol. 2014, 382, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Marquart, T.J.; Allen, R.M.; Ory, D.S.; Baldán, A. miR-33 links SREBP-2 induction to repression of sterol transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 12228–12232. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Xiao, C.; Wang, R.H.; Lahusen, T.; Xu, X.; Vassilopoulos, A.; Vazquez-Ortiz, G.; Jeong, W.I.; Park, O.; Ki, S.H.; et al. Hepatic-specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis. Cell Metab. 2010, 12, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Najafi-Shoushtari, S.H.; Kristo, F.; Li, Y.; Shioda, T.; Cohen, D.E.; Gerszten, R.E.; Näär, A.M. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science 2010, 328, 1566–1569. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.J.; Sheedy, F.J.; Esau, C.C.; Hussain, F.N.; Temel, R.E.; Parathath, S.; van Gils, J.M.; Rayner, A.J.; Chang, A.N.; Suarez, Y.; et al. Antagonism of miR-33 in mice promotes reverse cholesterol transport and regression of atherosclerosis. J. Clin. Investig. 2011, 121, 2921. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.J.; Esau, C.C.; Hussain, F.N.; McDaniel, A.L.; Marshall, S.M.; van Gils, J.M.; Ray, T.D.; Sheedy, F.J.; Goedeke, L.; Liu, X.; et al. Inhibition of miR-33a/b in non-human primates raises plasma HDL and lowers VLDL triglycerides. Nature 2011, 478, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Baba, O.; Kuwabara, Y.; Chujo, Y.; Watanabe, S.; Kinoshita, M.; Horiguchi, M.; Nakamura, T.; Chonabayashi, K.; Hishizawa, M.; et al. MicroRNA-33 deficiency reduces the progression of atherosclerotic plaque in ApoE−/− Mice. J. Am. Heart Assoc. 2012, 1, e003376. [Google Scholar] [CrossRef] [PubMed]
- TargetScanHuman. Available online: http://www.targetscan.org/ (accessed on 14 February 2015).
- miRDB. Available online: http://mirdb.org/miRDB/ (accessed on 14 February 2015).
- MicroCosm Targets Version 5. Available online: http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/ (accessed on 14 February 2015).
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Y.; Jiang, S.; Zhang, Y.; Zhang, R.; Gong, D. Detection of miR-33 Expression and the Verification of Its Target Genes in the Fatty Liver of Geese. Int. J. Mol. Sci. 2015, 16, 12737-12752. https://doi.org/10.3390/ijms160612737
Zheng Y, Jiang S, Zhang Y, Zhang R, Gong D. Detection of miR-33 Expression and the Verification of Its Target Genes in the Fatty Liver of Geese. International Journal of Molecular Sciences. 2015; 16(6):12737-12752. https://doi.org/10.3390/ijms160612737
Chicago/Turabian StyleZheng, Yun, Shibei Jiang, Yihui Zhang, Rui Zhang, and Daoqing Gong. 2015. "Detection of miR-33 Expression and the Verification of Its Target Genes in the Fatty Liver of Geese" International Journal of Molecular Sciences 16, no. 6: 12737-12752. https://doi.org/10.3390/ijms160612737