
Circulating Levels of sFlt1 Splice Variants as Predictive Markers for the Development of Preeclampsia

Abstract
:1. Introduction
2. Results
2.1. Generation of Total Soluble Fms-Like Tyrosine Kinase 1 (sFlt1) and Isoform-Specific Monoclonal Antibodies
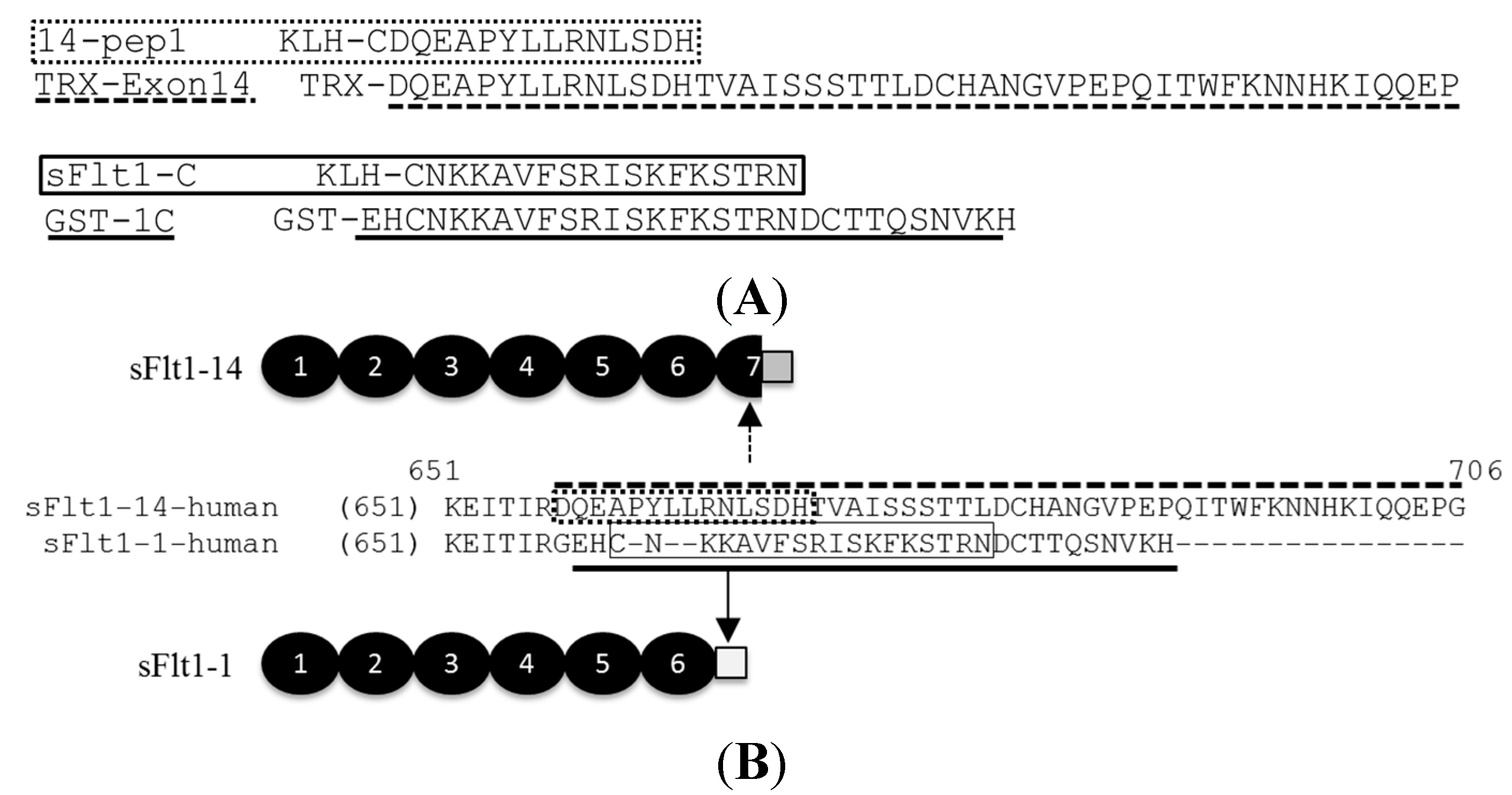
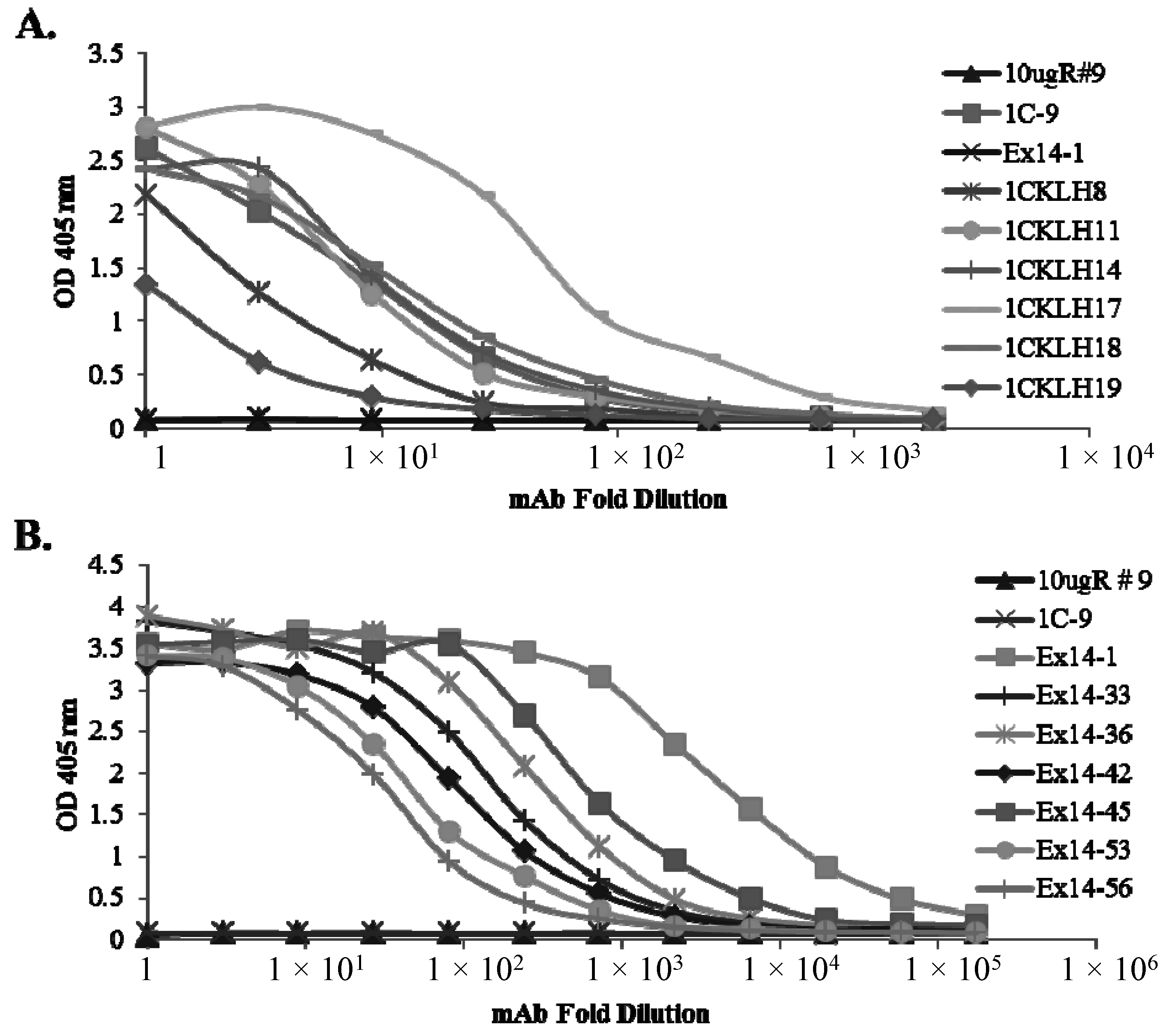
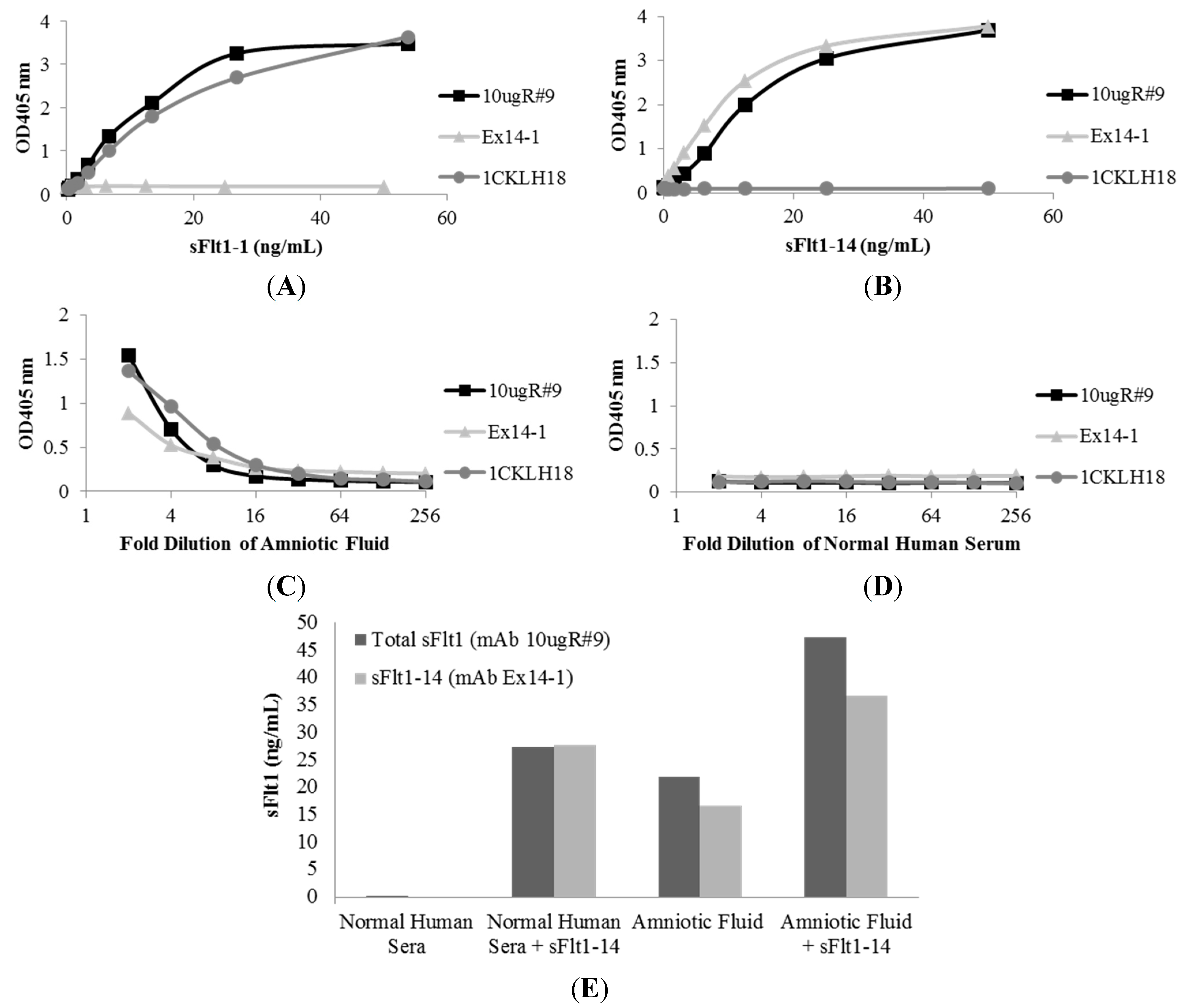
2.2. Characterization of sFlt1-Specific Monoclonal Antibodies by ELISA
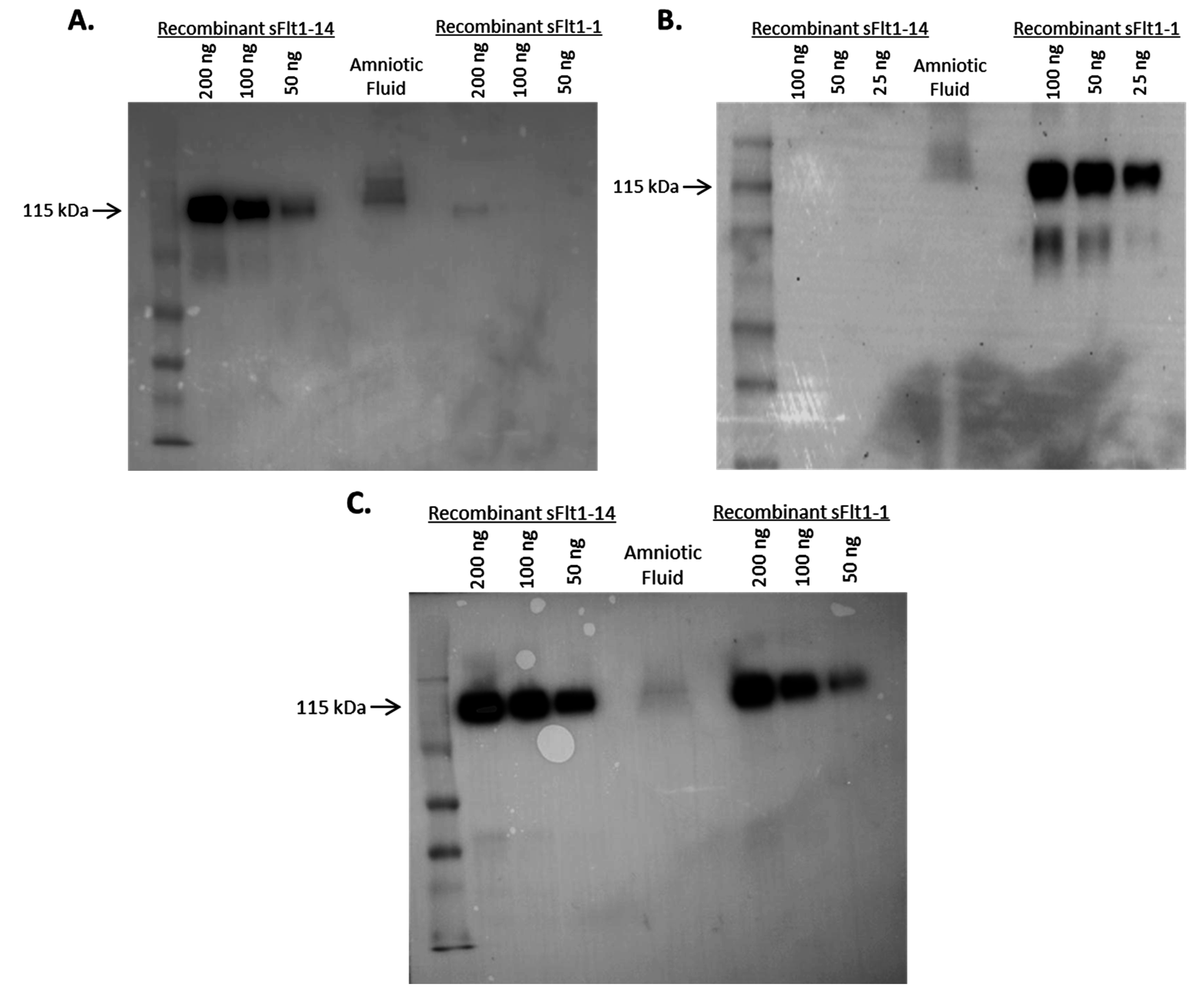
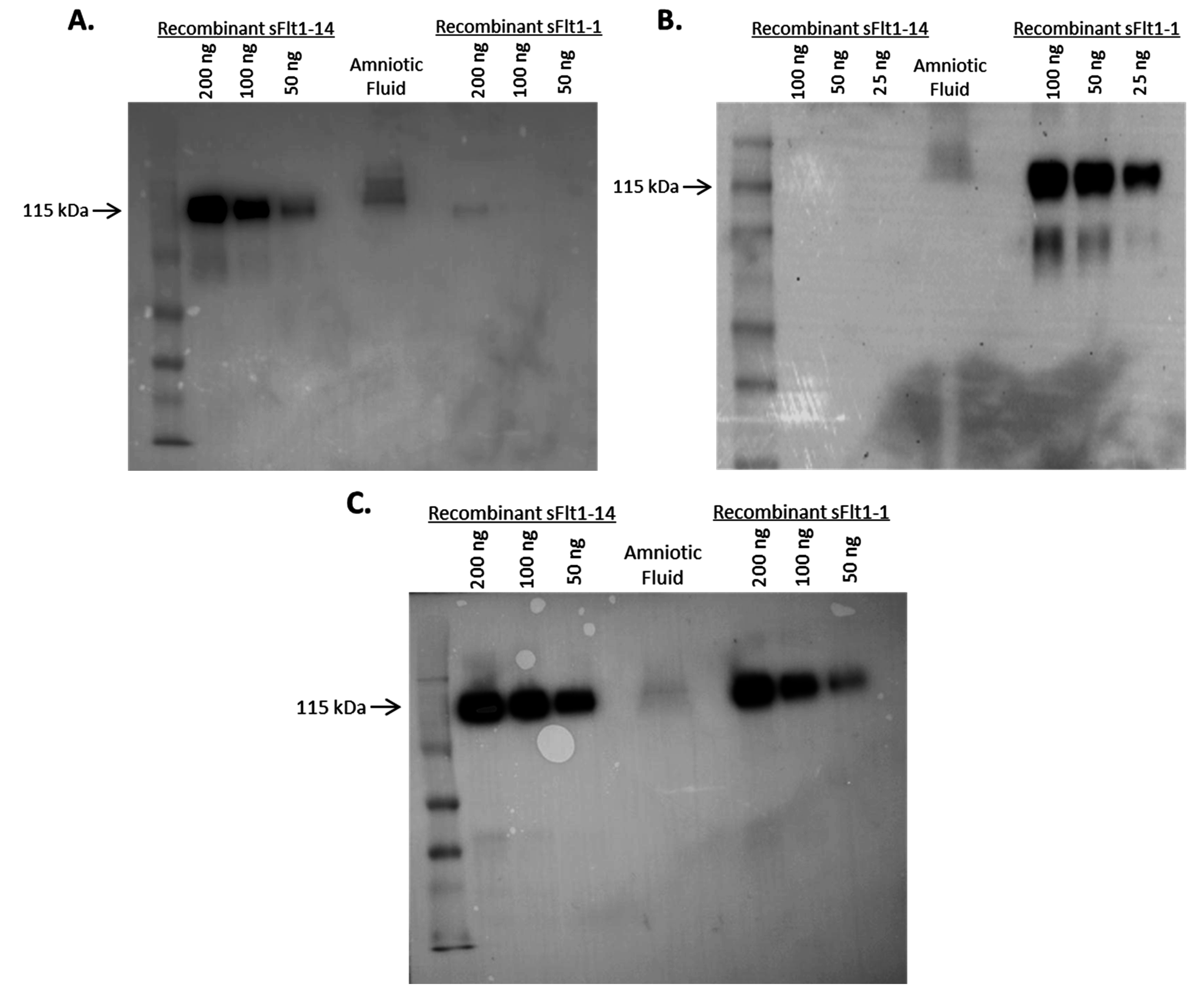
2.3. Specificity of Anti-sFlt1 Monoclonal Antibodies by Western Blot
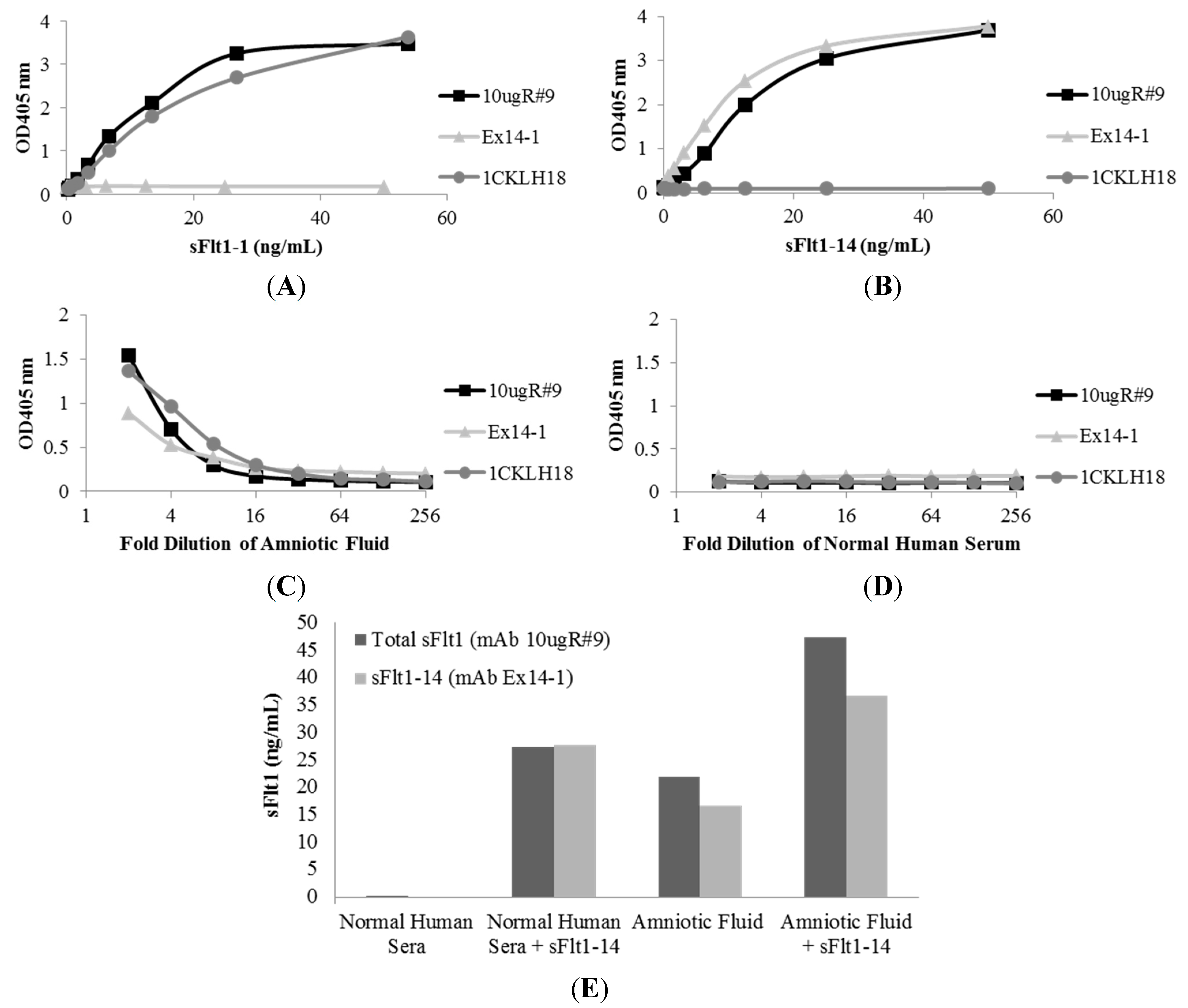
2.4. Affinity of Anti-sFlt1 Monoclonal Antibodies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Demographic Characteristics | Control Subjects (n = 137) * | Subjects with Preeclampsia (n = 15) * |
|---|---|---|
| Maternal age (years) | 31.3 (5.1) | 32.8 (5.6) |
| Gravity | 2.4 (1.5) | 2.1 (1.3) |
| Essential Nulliparity | 43 (31.4) | 7 (46.7) |
| Body mass index (kg/m2): | ||
| Underweight (BMI < 18.5) | 1 (0.7) | 0 (0.0) |
| Normal weight (BMI 18.5–24.9) | 23 (16.8) | 1 (6.7) |
| Overweight (BMI 25.0–29.9) | 43 (31.4) | 6 (40.0) |
| Obese (BMI ≥ 30.0) | 70 (51.1) | 8 (53.3) |
| Systolic BP at enrollment (mmHg) | 117.9 (14.4) | 123.1 (10.7) |
| Diastolic BP at enrollment (mmHg) | 71.7 (10.2) | 73.4 (15.9) |
| Anti-hypertensive medications at enrollment | 19 (13.9) | 5 (33.3) (exact p-value = 0.0639) |
| Smoking status: | ||
| Current smoker | 12 (8.8) | 0 (0.0) |
| Lives with smoker | 18 (13.1) | 3 (20.0) |
| Chronic hypertension † | 23 (16.8) | 6 (40.0) |
| Pregestational diabetes (DM1 or DM2) † | 16 (11.7) | 5 (33.3) |
| History of prior preeclampsia | 30 (21.9) | 1 (6.7) |
| Obese (BMI ≥ 30) nullipara | 28 (20.4) | 4 (26.7) |
| Any severe preeclampsia | 0 (0.0) | 12 (80.0) |
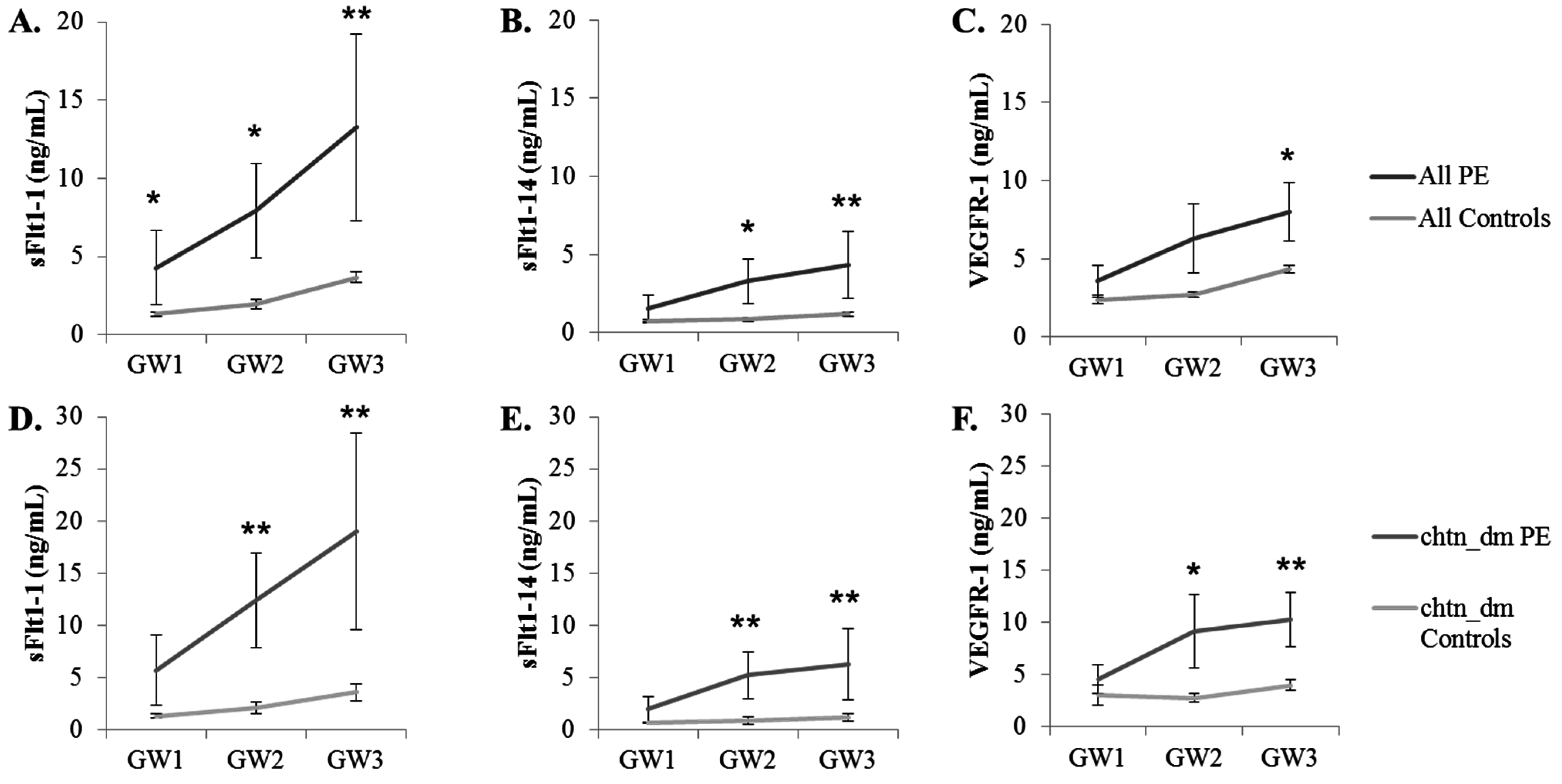
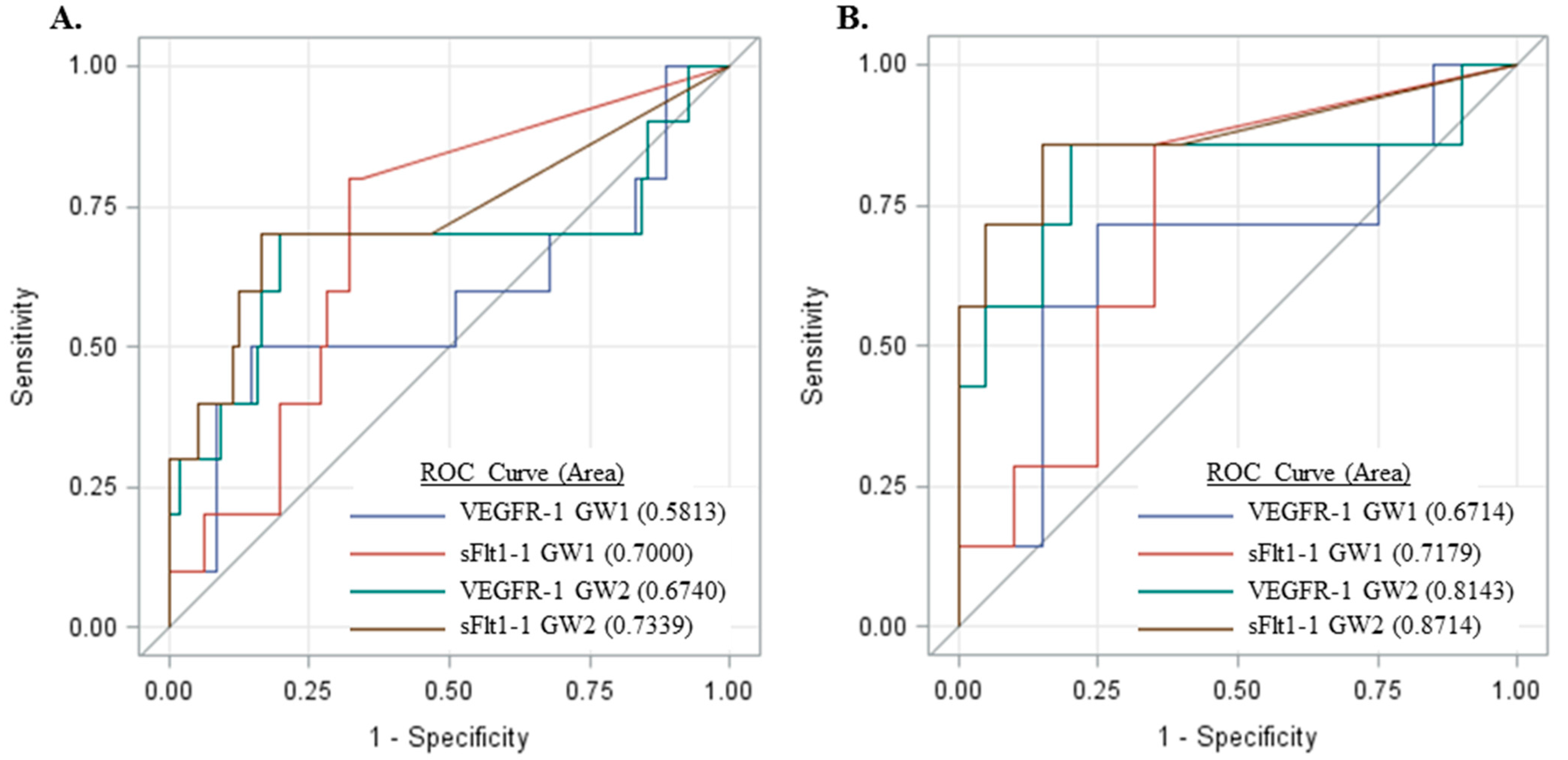
2.5. Measurement of sFlt1 Variants in the Sera of Pregnant Women

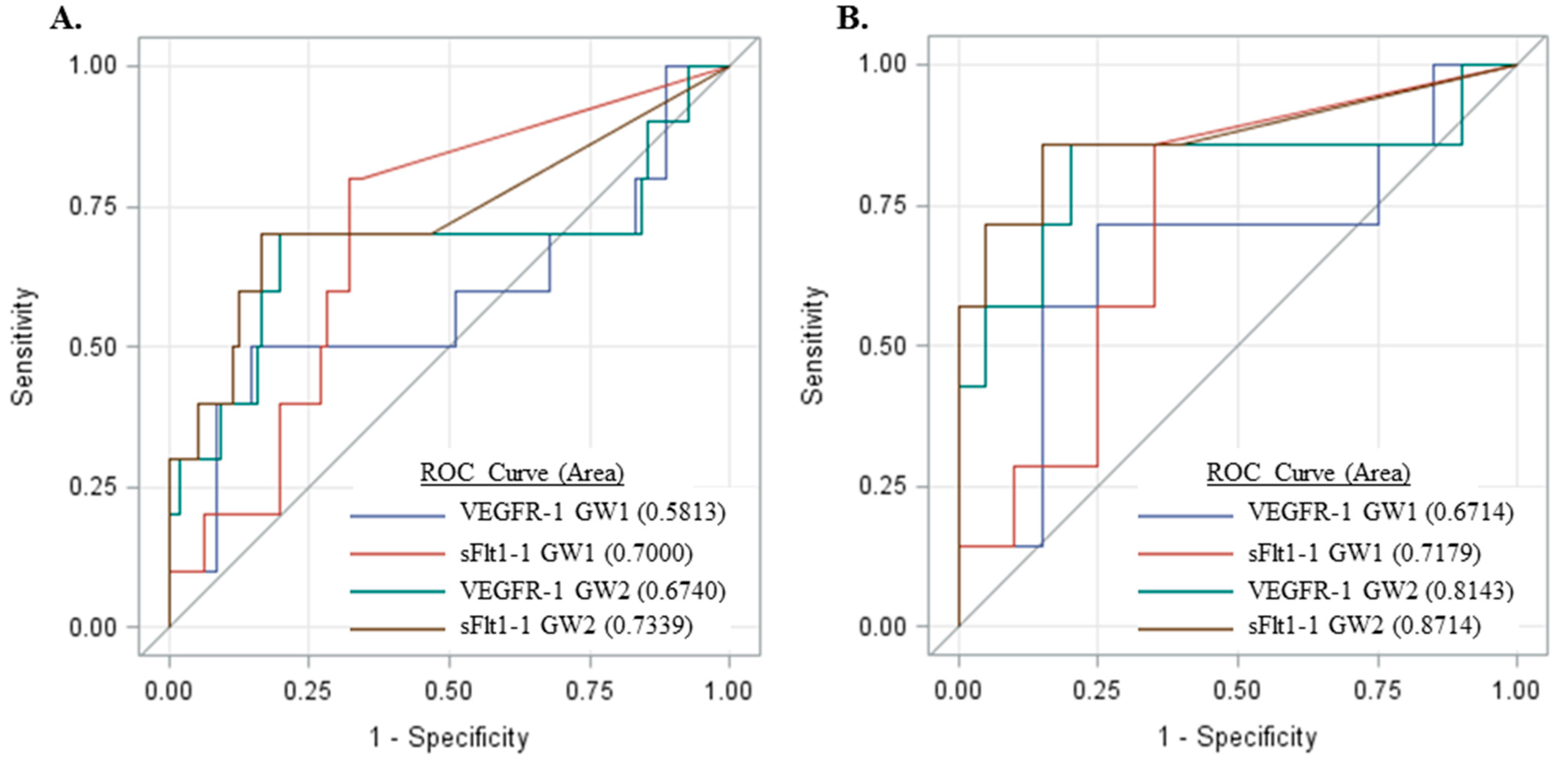
3. Discussion
4. Experimental Section
4.1. Chinese Hamster Ovary (CHO) Cell Expression and Purification of Full-Length sFlt1 Isoforms
4.2. Bacterial Expression of sFlt1-1 and sFlt1-14 C-Terminal Peptides
4.3. Generation of Monoclonal Antibodies
4.4. Screening and Characterization of mAbs
4.5. Western Blots
4.6. Quantitative Capture ELISA
4.7. Subjects and Serum Samples for Measurement of sFlt1 Isoforms
4.8. Statistics
Acknowledgments
Author Contributions
Abbreviations
| total sFlt1, VEGFR-1 | Soluble fms-like tyrosine kinase 1 |
| sFlt1-1 | soluble fms-like tyrosine kinase 1 isoform v1 |
| sFlt1-14 | soluble fms-like tyrosine kinase 1 isoform v2 |
Conflicts of Interest
References
- Abalos, E.; Cuesta, C.; Grosso, A.L.; Chou, D.; Say, L. Global and regional estimates of preeclampsia and eclampsia: A systematic review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Jaffe, I.Z.; Karumanchi, S.A. Preeclampsia and cardiovascular disease. Cardiovasc. Res. 2014, 101, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Steegers, E.A.; von Dadelszen, P.; Duvekot, J.J.; Pijnenborg, R. Preeclampsia. Lancet 2010, 376, 631–644. [Google Scholar] [CrossRef]
- Duley, L. The global impact of preeclampsia and eclampsia. Semin. Perinatol. 2009, 33, 130–137. [Google Scholar] [CrossRef] [PubMed]
- MacKay, A.P.; Berg, C.J.; Liu, X.; Duran, C.; Hoyert, D.L. Changes in pregnancy mortality ascertainment: United states, 1999–2005. Obstet. Gynecol. 2011, 118, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.M.; Caritis, S.; Hauth, J. What we have learned about preeclampsia. Semin. Perinatol. 2003, 27, 239–246. [Google Scholar] [CrossRef]
- Powe, C.E.; Thadhani, R. Diabetes and the kidney in pregnancy. Semin. Nephrol. 2011, 31, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.; Craft, N. Preeclampsia. BMJ 2012, 345, e4437. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.M.; Caritis, S.; Hauth, J.; Lindheimer, M.; VanDorsten, J.P.; MacPherson, C.; Klebanoff, M.; Landon, M.; Miodovnik, M.; Paul, R.; et al. Risks of preeclampsia and adverse neonatal outcomes among women with pregestational diabetes mellitus. Am. J. Obstet. Gynecol. 2000, 182, 364–369. [Google Scholar] [CrossRef]
- Cerdeira, A.S.; Karumanchi, S.A. Angiogenic factors in preeclampsia and related disorders. Cold Spring Harb. Perspect. Med. 2012, 2, a006585. [Google Scholar] [CrossRef] [PubMed]
- Verlohren, S.; Stepan, H.; Dechend, R. Angiogenic growth factors in the diagnosis and prediction of preeclampsia. Clin. Sci. 2012, 122, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Rana, S.; Karumanchi, S.A. Preeclampsia: The role of angiogenic factors in its pathogenesis. Physiology 2009, 24, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Ahmed, A. Elevated placental soluble vascular endothelial growth factor receptor-1 inhibits angiogenesis in preeclampsia. Circ. Res. 2004, 95, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Karumanchi, S.A.; Bdolah, Y. Hypoxia and sFlt-1 in preeclampsia: The “Chicken-and-egg” Question. Endocrinology 2004, 145, 4835–4837. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Luttun, A.; Carmeliet, P. Soluble VEGF receptor Flt1: The elusive preeclampsia factor discovered? J. Clin. Investig. 2003, 111, 600–602. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Min, J.Y.; Merchan, J.; Lim, K.H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.T.; Stefanini, M.O.; Mac Gabhann, F.; Kontos, C.D.; Annex, B.H.; Popel, A.S. A systems biology perspective on sVEGFR1: Its biological function, pathogenic role and therapeutic use. J. Cell Mol. Med. 2010, 14, 528–552. [Google Scholar] [CrossRef] [PubMed]
- Kendall, R.L.; Thomas, K.A. Inhibition of vascular endothelial cell growth factor activity by an endogenously encoded soluble receptor. Proc. Natl. Acad. Sci. USA 1993, 90, 10705–10709. [Google Scholar] [CrossRef] [PubMed]
- Heydarian, M.; McCaffrey, T.; Florea, L.; Yang, Z.; Ross, M.M.; Zhou, W.; Maynard, S.E. Novel splice variants of sFlt1 are upregulated in preeclampsia. Placenta 2009, 30, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.P.; Andrews, J.I.; Liu, K.Z. Intronic polyadenylation signal sequences and alternate splicing generate human soluble Flt1 variants and regulate the abundance of soluble Flt1 in the placenta. FASEB J. 2007, 21, 3885–3895. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.L.; Palmer, K.R.; Nilsson, U.; Gao, Y.; Saglam, B.; Lappas, M.; Tong, S. Placental expression of a novel primate-specific splice variant of sFlt-1 is upregulated in pregnancies complicated by severe early onset preeclampsia. BJOG 2011, 118, 1268–1271. [Google Scholar] [CrossRef] [PubMed]
- Jebbink, J.; Keijser, R.; Veenboer, G.; van der Post, J.; Ris-Stalpers, C.; Afink, G. Expression of placental FLT1 transcript variants relates to both gestational hypertensive disease and fetal growth. Hypertension 2011, 58, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Rajakumar, A.; Powers, R.W.; Hubel, C.A.; Shibata, E.; von Versen-Hoynck, F.; Plymire, D.; Jeyabalan, A. Novel soluble Flt-1 isoforms in plasma and cultured placental explants from normotensive pregnant and preeclamptic women. Placenta 2009, 30, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sela, S.; Itin, A.; Natanson-Yaron, S.; Greenfield, C.; Goldman-Wohl, D.; Yagel, S.; Keshet, E. A novel human-specific soluble vascular endothelial growth factor receptor 1: Cell-type-specific splicing and implications to vascular endothelial growth factor homeostasis and preeclampsia. Circ. Res. 2008, 102, 1566–1574. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.J.; Lam, C.; Qian, C.; Yu, K.F.; Maynard, S.E.; Sachs, B.P.; Sibai, B.M.; Epstein, F.H.; Romero, R.; Thadhani, R.; et al. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N. Engl. J. Med. 2006, 355, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Moore Simas, T.A.; Crawford, S.L.; Solitro, M.J.; Frost, S.C.; Meyer, B.A.; Maynard, S.E. Angiogenic factors for the prediction of preeclampsia in high-risk women. Am. J. Obstet. Gynecol. 2007, 197, 244.e1–244.e8. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Karumanchi, S.A.; Levine, R.J.; Venkatesha, S.; Rauh-Hain, J.A.; Tamez, H.; Thadhani, R. Sequential changes in antiangiogenic factors in early pregnancy and risk of developing preeclampsia. Hypertension 2007, 50, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Leanos-Miranda, A.; Campos-Galicia, I.; Isordia-Salas, I.; Rivera-Leanos, R.; Romero-Arauz, J.F.; Ayala-Mendez, J.A.; Ulloa-Aguirre, A. Changes in circulating concentrations of soluble fms-like tyrosine kinase-1 and placental growth factor measured by automated electrochemiluminescence immunoassays methods are predictors of preeclampsia. J. Hypertens. 2012, 30, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, S.K.; Larkin, J.; Sammel, M.D.; Appleby, D.; Bastek, J.; Andrela, C.M.; Ofori, E.; Elovitz, M.A. The use of angiogenic factors in discriminating preeclampsia: Are they ready for prime time? J. Matern. Fetal Neonatal. Med. 2010, 23, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.E.; Crawford, S.L.; Bathgate, S.; Yan, J.; Robidoux, L.; Moore, M.; Moore Simas, T.A. Gestational angiogenic biomarker patterns in high risk preeclampsia groups. Am. J. Obstet. Gynecol. 2013, 209, 53.e1–53.e9. [Google Scholar] [CrossRef] [PubMed]
- Moore Simas, T.A.; Crawford, S.L.; Bathgate, S.; Yan, J.; Robidoux, L.; Moore, M.; Maynard, S.E. Angiogenic biomarkers for prediction of early preeclampsia onset in high-risk women. J. Matern. Fetal Neonatal. Med. 2014, 27, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Verlohren, S.; Galindo, A.; Schlembach, D.; Zeisler, H.; Herraiz, I.; Moertl, M.G.; Pape, J.; Dudenhausen, J.W.; Denk, B.; Stepan, H. An automated method for the determination of the sFlt-1/PIGF ratio in the assessment of preeclampsia. Am. J. Obstet. Gynecol. 2010, 202, 161.e1–161.e11. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Powe, C.E.; Salahuddin, S.; Verlohren, S.; Perschel, F.H.; Levine, R.J.; Lim, K.H.; Wenger, J.B.; Thadhani, R.; Karumanchi, S.A. Angiogenic factors and the risk of adverse outcomes in women with suspected preeclampsia. Circulation 2012, 125, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Verlohren, S.; Herraiz, I.; Lapaire, O.; Schlembach, D.; Moertl, M.; Zeisler, H.; Calda, P.; Holzgreve, W.; Galindo, A.; Engels, T.; et al. The sFlt-1/PLGF ratio in different types of hypertensive pregnancy disorders and its prognostic potential in preeclamptic patients. Am. J. Obstet. Gynecol. 2012, 206, 58.e1–58.e8. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Jiang, W.G.; Grant, M.B.; Boulton, M. Pigment epithelium-derived factor inhibits angiogenesis via regulated intracellular proteolysis of vascular endothelial growth factor receptor 1. J. Biol. Chem. 2006, 281, 3604–3613. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N.; Golde, T.E.; Meyer, R.D. Identification of ligand-induced proteolytic cleavage and ectodomain shedding of VEGFR-1/FLT1 in leukemic cancer cells. Cancer Res. 2009, 69, 2607–2614. [Google Scholar] [CrossRef] [PubMed]
- Surmon, L.; Bobek, G.; Makris, A.; Chiu, C.L.; Lind, C.A.; Lind, J.M.; Hennessy, A. Variability in mrna expression of fms-like tyrosine kinase-1 variants in normal and preeclamptic placenta. BMC Res. Notes 2014, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Kuklina, E.V.; Ayala, C.; Callaghan, W.M. Hypertensive disorders and severe obstetric morbidity in the United States. Obstet. Gynecol. 2009, 113, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Buhimschi, C.S.; Norwitz, E.R.; Funai, E.; Richman, S.; Guller, S.; Lockwood, C.J.; Buhimschi, I.A. Urinary angiogenic factors cluster hypertensive disorders and identify women with severe preeclampsia. Am. J. Obstet. Gynecol. 2005, 192, 734–741. [Google Scholar] [CrossRef] [PubMed]
- ACOG Practice Bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, january 2002. American college of obstetricians and gynecologists. Int. J. Gynaecol. Obstet. 2002, 77, 67–75. [Google Scholar]
- Maynard, S.E.; Moore Simas, T.A.; Solitro, M.J.; Rajan, A.; Crawford, S.; Soderland, P.; Meyer, B.A. Circulating angiogenic factors in singleton vs. multiple-gestation pregnancies. Am. J. Obstet. Gynecol. 2008, 198, 200.e1–200.e7. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souders, C.A.; Maynard, S.E.; Yan, J.; Wang, Y.; Boatright, N.K.; Sedan, J.; Balyozian, D.; Cheslock, P.S.; Molrine, D.C.; Simas, T.A.M. Circulating Levels of sFlt1 Splice Variants as Predictive Markers for the Development of Preeclampsia. Int. J. Mol. Sci. 2015, 16, 12436-12453. https://doi.org/10.3390/ijms160612436
Souders CA, Maynard SE, Yan J, Wang Y, Boatright NK, Sedan J, Balyozian D, Cheslock PS, Molrine DC, Simas TAM. Circulating Levels of sFlt1 Splice Variants as Predictive Markers for the Development of Preeclampsia. International Journal of Molecular Sciences. 2015; 16(6):12436-12453. https://doi.org/10.3390/ijms160612436
Chicago/Turabian StyleSouders, Colby A., Sharon E. Maynard, Jing Yan, Yang Wang, Naomi K. Boatright, Jessica Sedan, David Balyozian, Peter S. Cheslock, Deborah C. Molrine, and Tiffany A. Moore Simas. 2015. "Circulating Levels of sFlt1 Splice Variants as Predictive Markers for the Development of Preeclampsia" International Journal of Molecular Sciences 16, no. 6: 12436-12453. https://doi.org/10.3390/ijms160612436