
Resistance of Lung Cancer Cells Grown as Multicellular Tumour Spheroids to Zinc Sulfophthalocyanine Photosensitization



Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
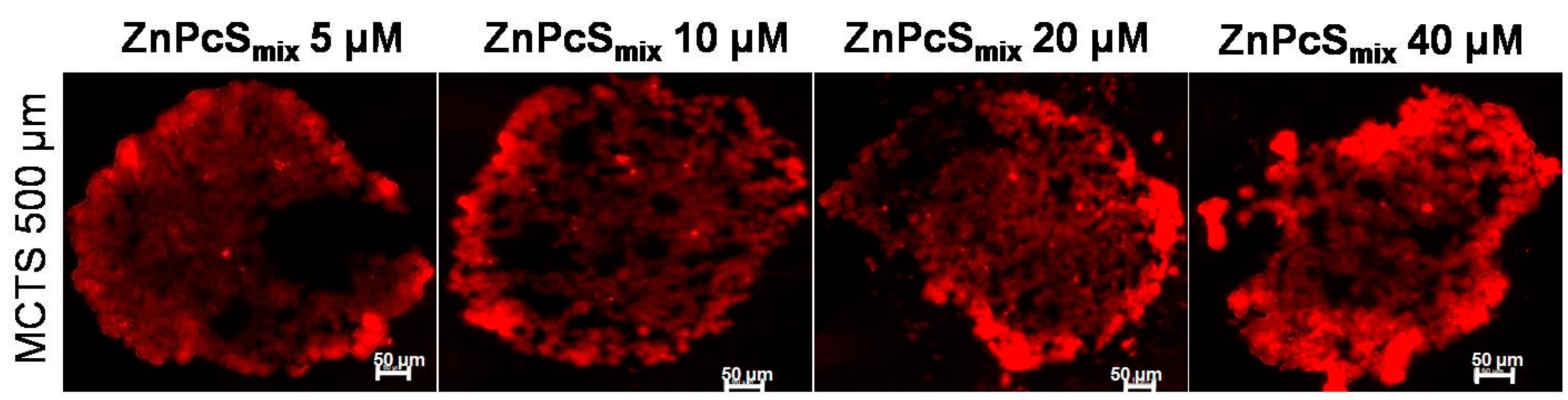
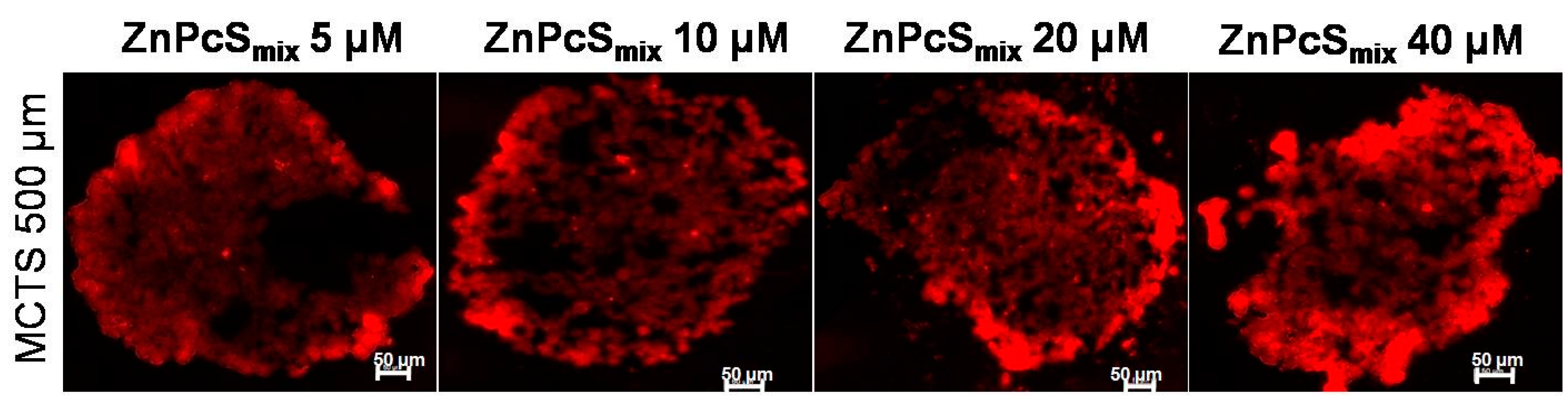
2.1.1. Zinc Sulfophthalocyanine (ZnPcSmix) Distribution in Multicellular Tumor Spheroids (MCTSs)
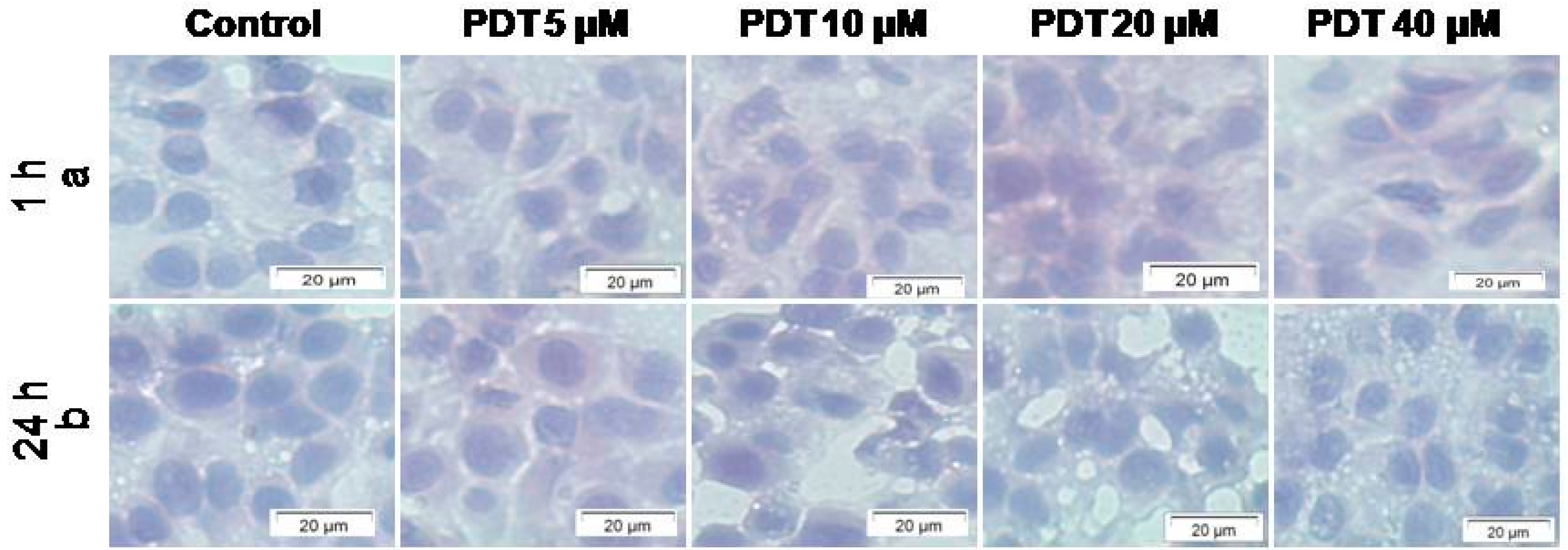
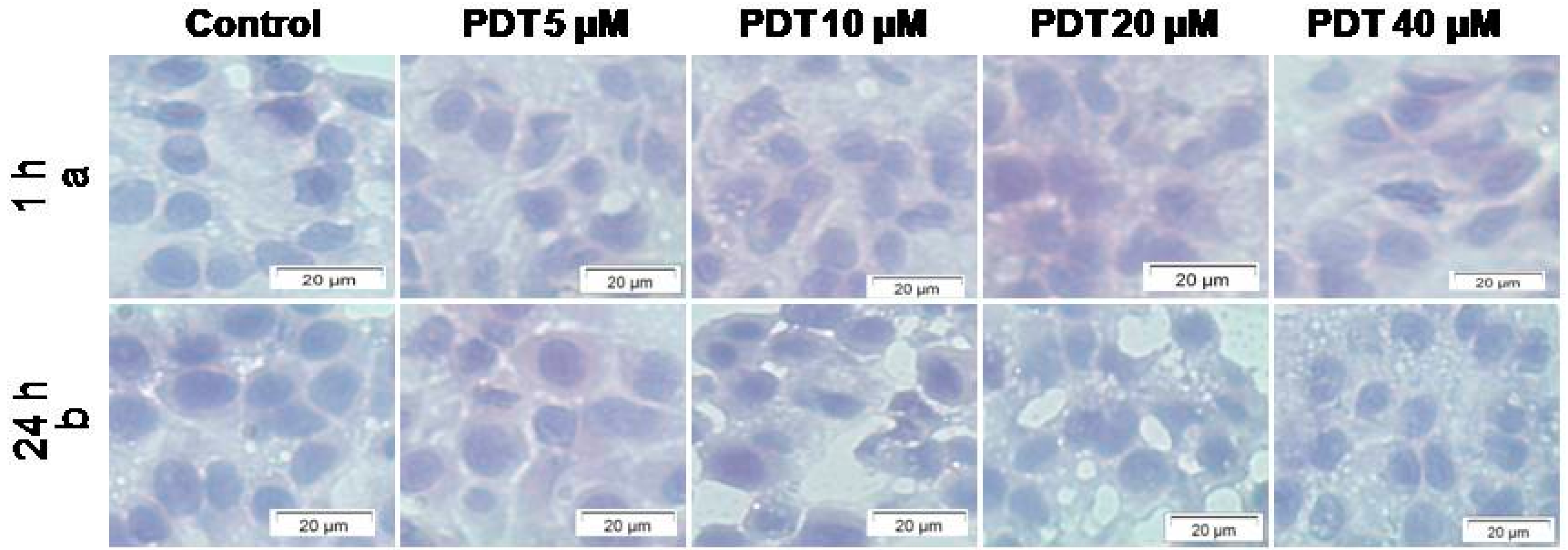
2.1.2. Cellular Morphology
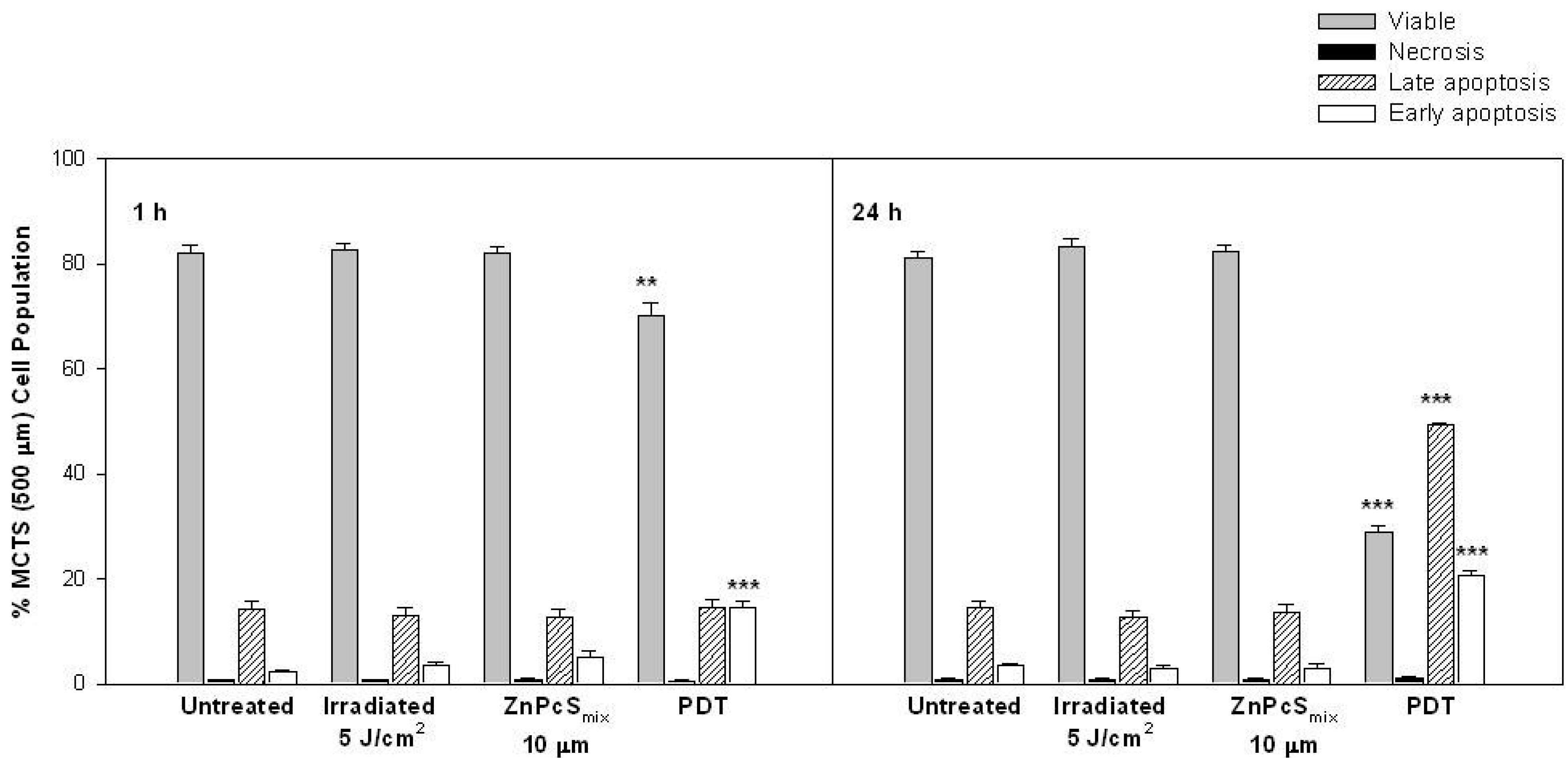
2.1.3. Cellular Viability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Function | Fluence | 0 µM | 5 µM | 10 µM | 20 µM | 40 µM | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Monolayer | MCTS | Monolayer | MCTS | Monolayer | MCTS | Monolayer | MCTS | Monolayer | MCTS | ||
| Viability | 0 J/cm2 | 88.35 a ± 3.50 b | 85.5 ± 2.22 | 97.25 ± 0.25 | 84.00 ± 1.29 | 96.75 ± 0.48 | 89.75 ± 2.63 | 98.5 ± 0.288 | 87.00 ± 2.55 | 96.75 ± 1.11 | 85.00 ± 3.29 |
| 5 J/cm2 | 91.75 a ± 2.06 b | 89.25 ± 2.17 | 69.75 *** ± 1.65 | 55.5 *** ± 3.59 | 45.75 *** ± 3.61 | 33.5 *** ± 1.55 | 39.00 *** ± 9.05 | 25.00 *** ± 2.68 | 37.25 *** ± 4.385 | 21.25 *** ± 1.31 | |
| Proliferation | 0 J/cm2 | 279,325 ± 1328.11 | 23,629 ± 700.93 | 313,316 ± 5179.75 | 25,332 ± 2609.47 | 319,120 ± 11,247.98 | 28,879 ± 890.52 | 289,718 ± 14,702.04 | 27,987 ± 1133.90 | 277,319 ± 21,847.83 | 25,691 ± 1478.26 |
| 5 J/cm2 | 285,533 ± 8139.17 | 21,129 ± 431.37 | 19,450 *** ± 3414.49 | 832 *** ± 99.22 | 175,827 *** ± 1592.56 | 2796 *** ± 393.48 | 140,123 *** ± 3883.81 | 1179 *** ± 90.88 | 5809 *** ± 364.67 | 420 *** ± 66.94 | |
| Cytotoxicity | 0 J/cm2 | 0.343 ± 0.02 | 0.249 ± 1.18 | 0.339 ± 9.71 | 0.25 ± 2.43 | 0.33 ± 0.01 | 0.26 ± 0.01 | 0.36 ± 8.31 | 0.23 ± 3.3 | 0.37 ± 0.01 | 0.232 ± 0.11 |
| 5 J/cm2 | 0.288 ± 0.02 | 0.249 ± 1.18 | 1.803 *** ± 0.14 | 0.876 *** ± 0.07 | 2.146 *** ± 0.17 | 0.851 *** ± 0.09 | 1.720 *** ± 0.12 | 0.882 *** ± 0.07 | 2.185 *** ± 0.34 | 0.569 *** ± 0.02 | |
2.1.4. Cellular Proliferation
2.1.5. Cellular Cytotoxicity
2.1.6. Cell Death Analysis

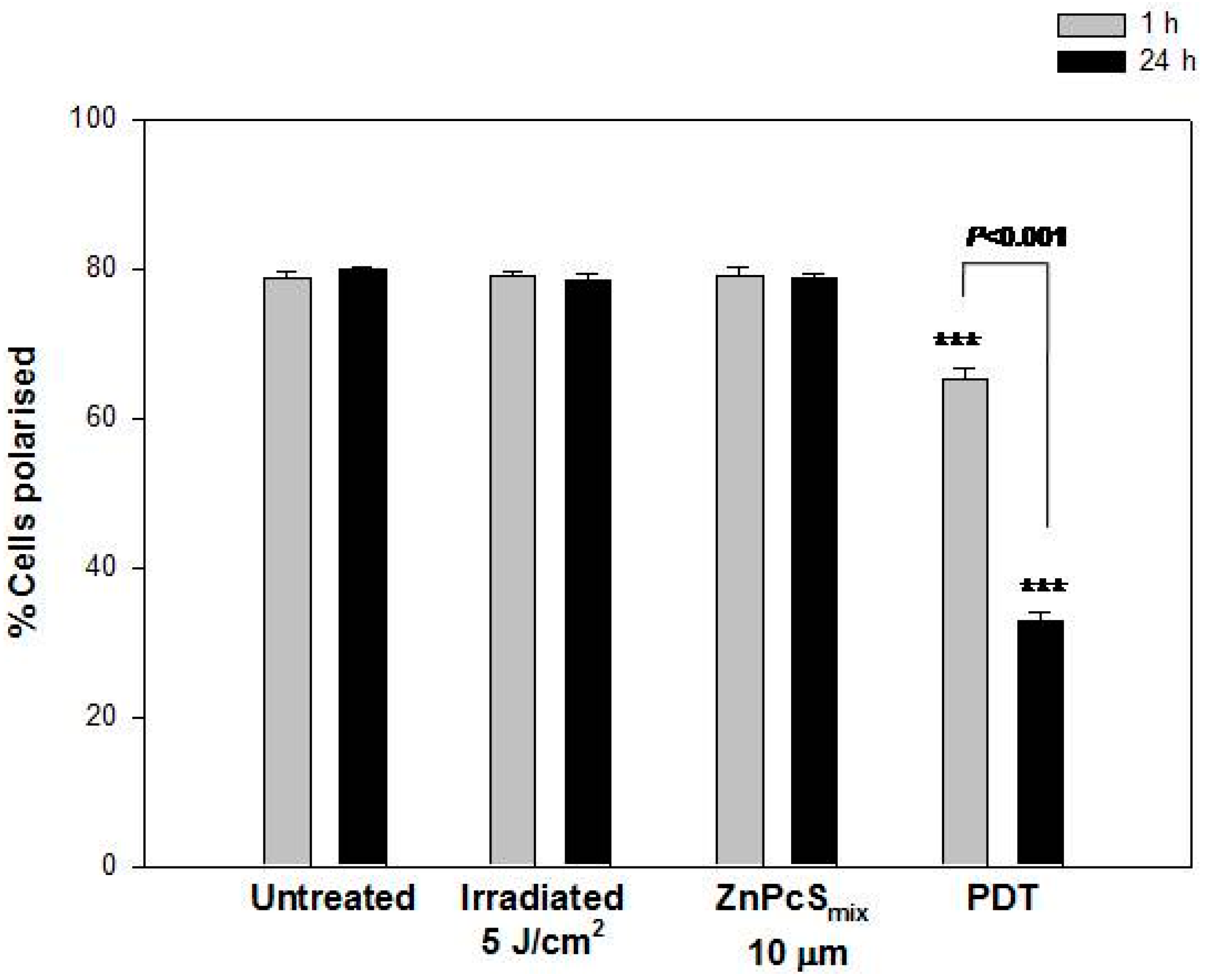
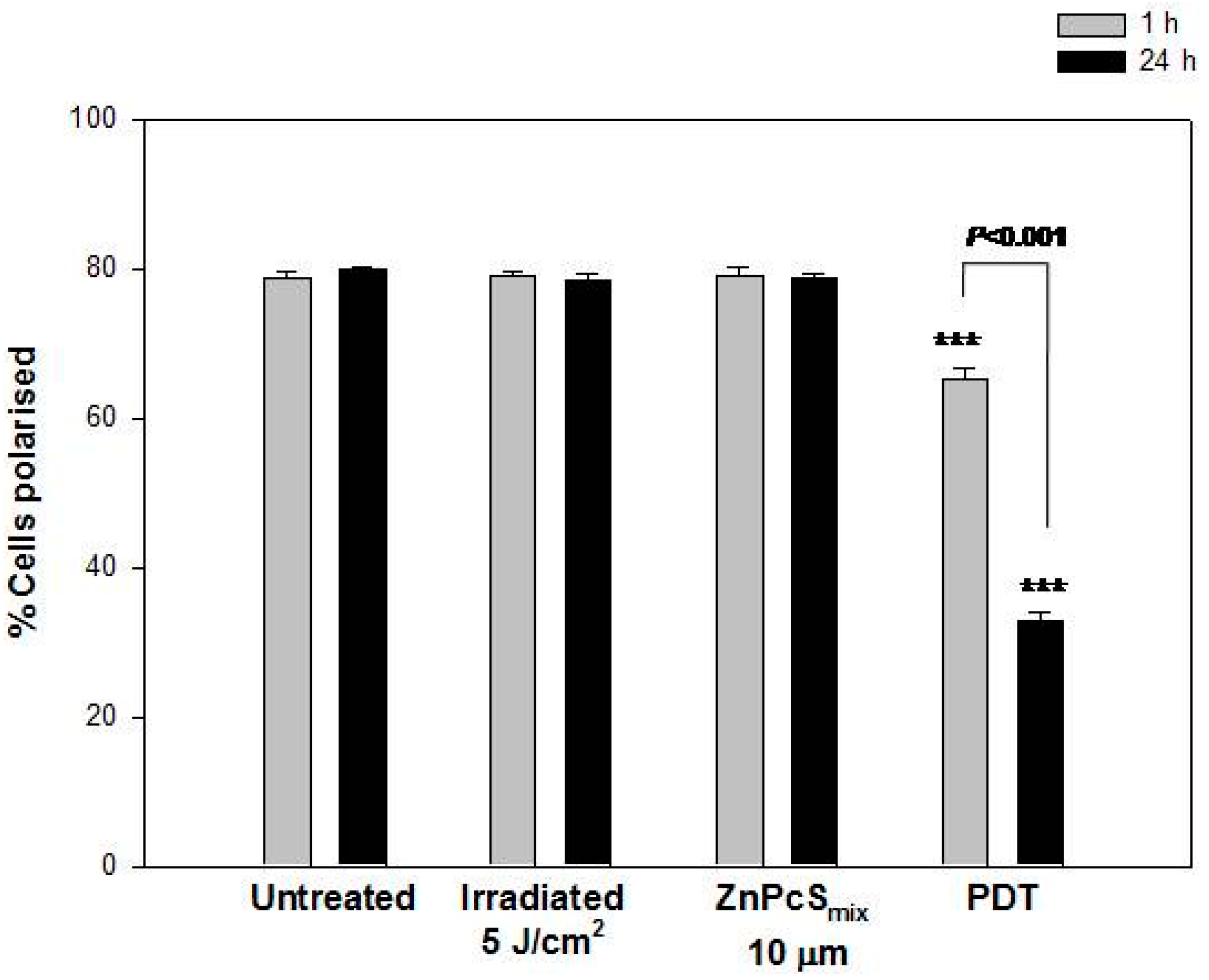
2.1.7. Mitochondrial Membrane Potential
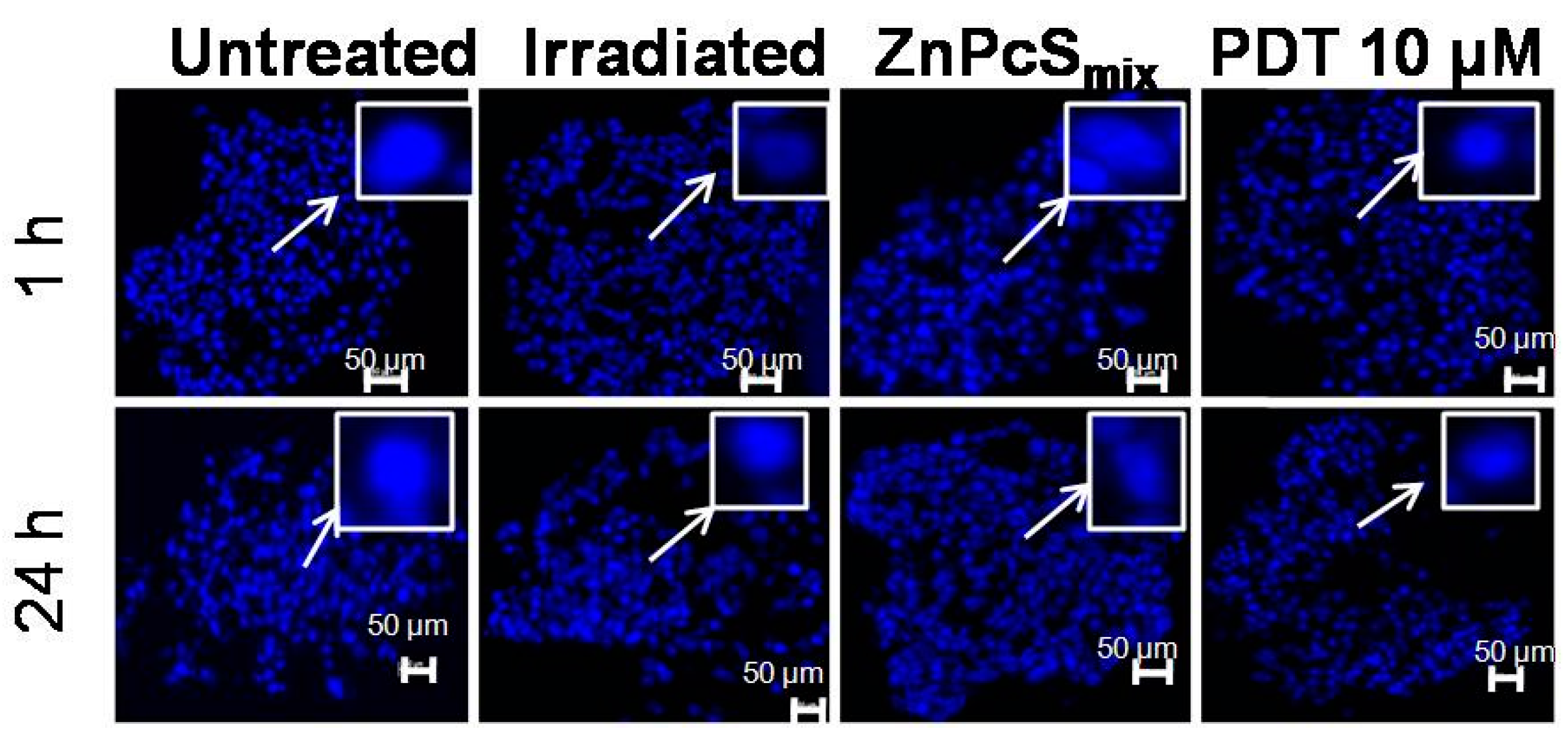
2.1.8. Nuclear Morphology
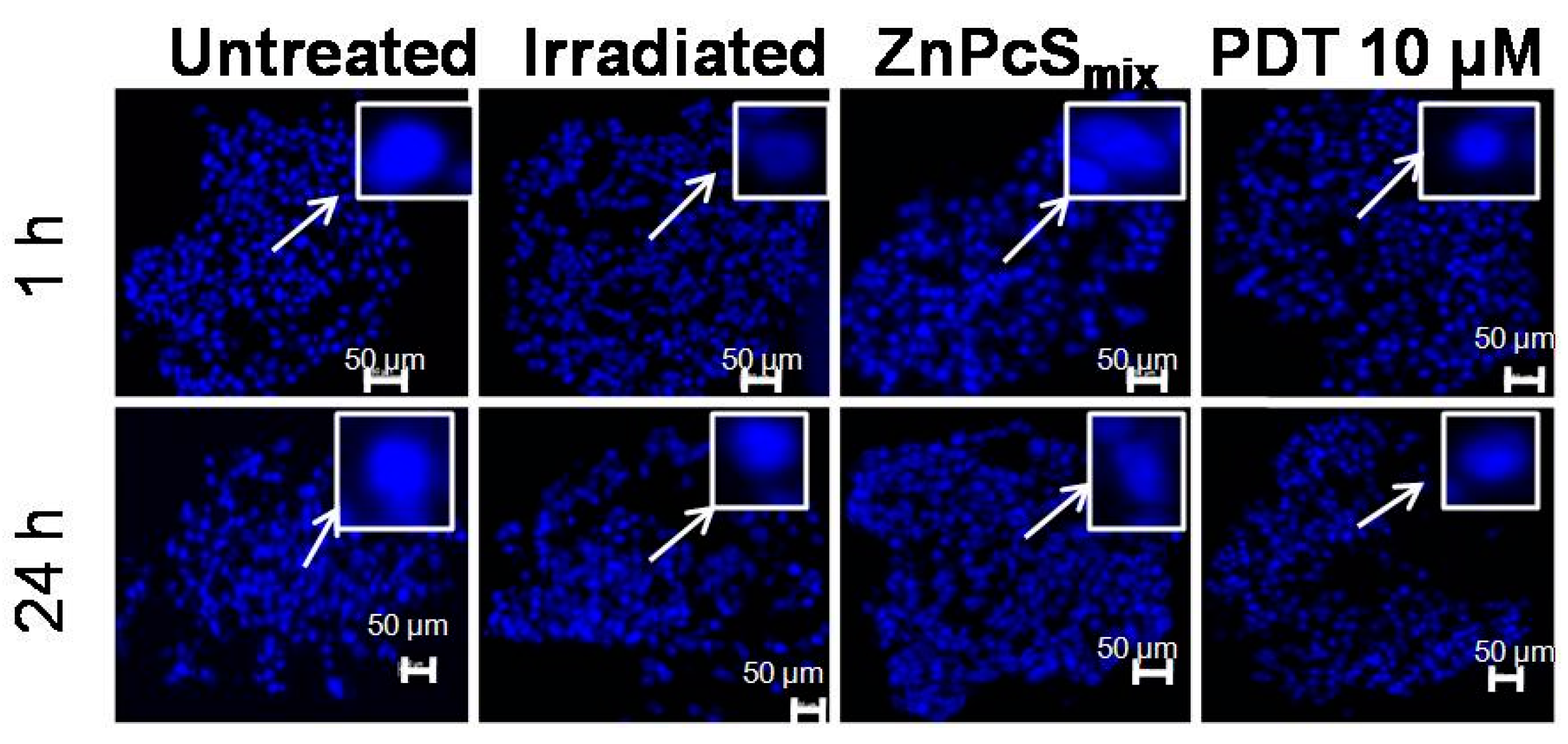
2.2. Discussion
3. Experimental Section
3.1. MCTS Cell Culture
3.2. ZnPcSmix Distribution in MCTSs
3.3. Photodynamic Treatment of MCTS
| Laser Type | Diode |
|---|---|
| Wavelength | 680 nm |
| Spot size | 9.1 cm2 |
| Power density | 4.87 mW/cm2 |
| Energy density (Fluence) | 5 J/cm2 |
| Output power | 44.2 mW |
| Output mode | Continuous |
| Duration of exposure | 17 min 7 s |
3.4. Cell Morphology
3.5. Cellular Viability
3.6. Cellular Proliferation
3.7. Cellular Cytotoxicity
3.8. Cell Death Analysis
3.9. Mitochondrial Membrane Potential
3.10. Nuclear Morphology
3.11. Statistics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- American Cancer Society. Cancer Facts and Figures 2011. American Cancer Society, 2011. Available online: http://www.cancer.org/Research/CancerFactsFigures/CancerFactsFigures/cancer-facts-figures-2011 (accessed on 28 January 2015).
- World Health Organisation. IARC Launches the Definitive Cancer Statistics Resource GLOBOCAN 2008. Press Release 201. 2010. Available online: http://www.iarc.fr/en/media-centre/pr/2010/pdfs/pr201_E.pdf (accessed on 19 January 2015).
- Wang, L.; Huang, Z.; Lin, H.; Li, Z.; Hetzel, F.; Liu, B. Effect of photofrin-mediated photocytotoxicity on a panel of human pancreatic cancer cells. Photodiagn. Photodyn. Ther. 2013, 10, 244–251. [Google Scholar] [CrossRef]
- Manoto, S.L.; Houreld, N.N.; Abrahamse, H. Phototoxic effect of photodynamic therapy on lung cancer cells grown as a monolayer and three dimensional multicellular spheroids. Lasers Surg. Med. 2013, 45, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Manoto, S.L.; Abrahamse, H. Effect of a newly synthesized Zn sulfophthalocyanine derivative on cell morphology, viability, proliferation, and cytotoxicity in a human lung cancer cell line (A549). Lasers Med. Sci. 2011, 26, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Jori, G. Tumour photosensitizers: Approaches to enhance the selectivity and efficiency of photodynamic therapy. Photochem. Photobiol. B 1996, 36, 87–93. [Google Scholar] [CrossRef]
- Friedrich, J.; Ebner, R.; Kunz-Schughart, L.A. Experimental anti-tumour therapy in 3-D: Spheroids-old hat or new challenge? Int. J. Radic. Biol. 2007, 83, 849–871. [Google Scholar] [CrossRef]
- Sutherland, R.M.; Inch, W.R.; McCredie, J.A.; Kruuv, J. A multicomponent radiation survival curve using an in vivo tumour model. Int. J. Radic. Biol. 1970, 18, 491–495. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Menne, H.; Dittfeld, C.; West, J.; Mueller-Klieser, W.; Kunz-Schughart, L.A. Multicellular tumour spheroids: An underestimated tool is catching up again. J. Biotechnol. 2010, 148, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Dubessy, C.; Merlin, J.L.; Marchal, C.; Guillemin, F. Spheroids in radiobiology and photodynamic therapy. Crit. Rev. Oncol. Hematol. 2000, 36, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.T.; Hughes-Fulford, M. Monolayer and spheroid culture of human liver hepatocellular carcinoma cell line cells demonstrate distinct global gene expression patterns and functional phenotypes. Tissue Eng. A 2009, 15, 559–567. [Google Scholar] [CrossRef]
- Cody, N.A.; Zietarska, M.; Filali-Mouhim, A.; Provencher, D.M.; Mes-Masson, A.M.; Tonin, P.N. Influence of monolayer, spheroid, and tumour growth conditions on chromosomes 3 gene expression in tumourigenic epithelial ovarian cancer cell lines. BMC Med. Genomics 2008, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Marchal, S.; Fadloun, A.; Maugain, E.; D’Hallewin, M.; Guillemin, F.; Bezdetnaya, L. Necrotic and apoptotic features of cell death in response to Foscan® photosensitization of HT29 monolayer and multicell spheroids. Biochem. Pharmacol. 2005, 69, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Seotsanyana-Makhosi, I.; Kresfelder, T.; Abrahamse, H.; Nyokong, T. The effect of Ge, Si and Sn phthalocyanine photosensitizers on cell proliferation and viability of human oesophageal carcinoma cells. Photochem. Photobiol. B 2006, 83, 55–62. [Google Scholar] [CrossRef]
- Laranjo, M.; Serra, A.C.; Abrantes, M.; Pineiro, M.; Goncalves, A.C.; Casalta-Lopes, J.; Carvalho, L.; Sarmento-Ribeiro, A.N.; Rocha-Gonsalves, A.; Botelho, F. 2-Bromo-5-hydroxyphenylporphyrins for photodynamic therapy: Photosensitization efficiency, subcellular localization and in vivo studies. Photodiagn. Photodyn. Ther. 2012, 10, 51–61. [Google Scholar] [CrossRef]
- Kresfelder, T.L.; Cronjé, M.J.; Abrahamse, H. The effects of two metallophalocyanines on the viability and proliferation of an esophageal cancer cell line. Photomed. Laser Surg. 2009, 27, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Tynga, I.M.; Houreld, N.N.; Abrahamse, H. The primary subcellular localization of zinc phthalocyanine and its cellular impact on viability, proliferation and structure of breast cancer cells (MCF-7). Photochem. Photobiol. B 2012, 120, 171–176. [Google Scholar] [CrossRef]
- Ho, W.Y.; Yeap, S.K.; Ho, C.L.; Rahim, R.A.; Alitheen, N.B. Development of multicellular tumour spheroids (MCTS) culture from breast cancer cell and a high throughput screening method using the MTT assay. PLoS ONE 2012, 7, 1–7. [Google Scholar] [CrossRef]
- Madsen, S.J.; Sun, C.; Tromberg, B.J.; Cristini, V.; Magalhaes, N.; Hirschberg, H. Multicell tumour spheroids in photodynamic therapy. Lasers Surg. Med. 2006, 38, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Oleinick, N.L.; Morris, R.L.; Nieminem, A.L. Photodynamic therapy-induced apoptosis. In Apoptosis, Senescence and Cancer; Gewritz, D.A., Holt, S.E., Grant, S., Eds.; Cancer Drug Discovery and Development, Humana Press, Inc.: Totowa, NJ, USA, 2007; pp. 557–577. [Google Scholar]
- Xu, D.D.; Lam, H.M.; Hoeven, R.; Xu, B.C.; Leung, A.W.N.L.; Cho, W.C.S. Photodynamic therapy induced cell death of hormone insensitive prostate cancer PC-3 cells with autophagic characteristics. Photodiagn. Photodyn. Ther. 2013, 10, 278–287. [Google Scholar] [CrossRef]
- Soriano, J.; Villanueva, A.; Stockert, C.; Magdelena, C. Regulated necrosis in Hela cells induced by ZnPc photodynamic treatment: A new nuclear morphology. Int. J. Mol. Sci. 2014, 15, 22772–22785. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.C.E. Signaling pathways in cell death and survival after photodynamic therapy. Photochem. Photobiol. B 2000, 57, 1–13. [Google Scholar] [CrossRef]
- Shao, J.; Xue, J.; Dai, Y.; Liu, H.; Jia, L.; Chen, N.; Huang, J. Inhibition of human hepatocellular carcinoma HepG2 by phthalocyanine photosensitizer ZnPcS2P2: ROS production, apoptosis, cell cycle arrest. Eur. J. Cancer 2012, 48, 2086–2096. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, C.; Bonora, M.; Missiroli, S.; Poletti, F.; Ramirez, G.; Morciano, G.; Morganti, C.; Pandolfi, P.P.; Mammano, F.; Pinton, P. Intravital imaging reveals p53 dependent cancer cell death induced by phototherapy via calcium signaling. Oncotarget 2014, 6, 1435–1445. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manoto, S.L.; Houreld, N.N.; Abrahamse, H. Resistance of Lung Cancer Cells Grown as Multicellular Tumour Spheroids to Zinc Sulfophthalocyanine Photosensitization. Int. J. Mol. Sci. 2015, 16, 10185-10200. https://doi.org/10.3390/ijms160510185
Manoto SL, Houreld NN, Abrahamse H. Resistance of Lung Cancer Cells Grown as Multicellular Tumour Spheroids to Zinc Sulfophthalocyanine Photosensitization. International Journal of Molecular Sciences. 2015; 16(5):10185-10200. https://doi.org/10.3390/ijms160510185
Chicago/Turabian StyleManoto, Sello Lebohang, Nicolette Nadene Houreld, and Heidi Abrahamse. 2015. "Resistance of Lung Cancer Cells Grown as Multicellular Tumour Spheroids to Zinc Sulfophthalocyanine Photosensitization" International Journal of Molecular Sciences 16, no. 5: 10185-10200. https://doi.org/10.3390/ijms160510185