
Effects of Aflatoxin B1 on T-Cell Subsets and mRNA Expression of Cytokines in the Intestine of Broilers


Abstract
:1. Introduction
2. Results
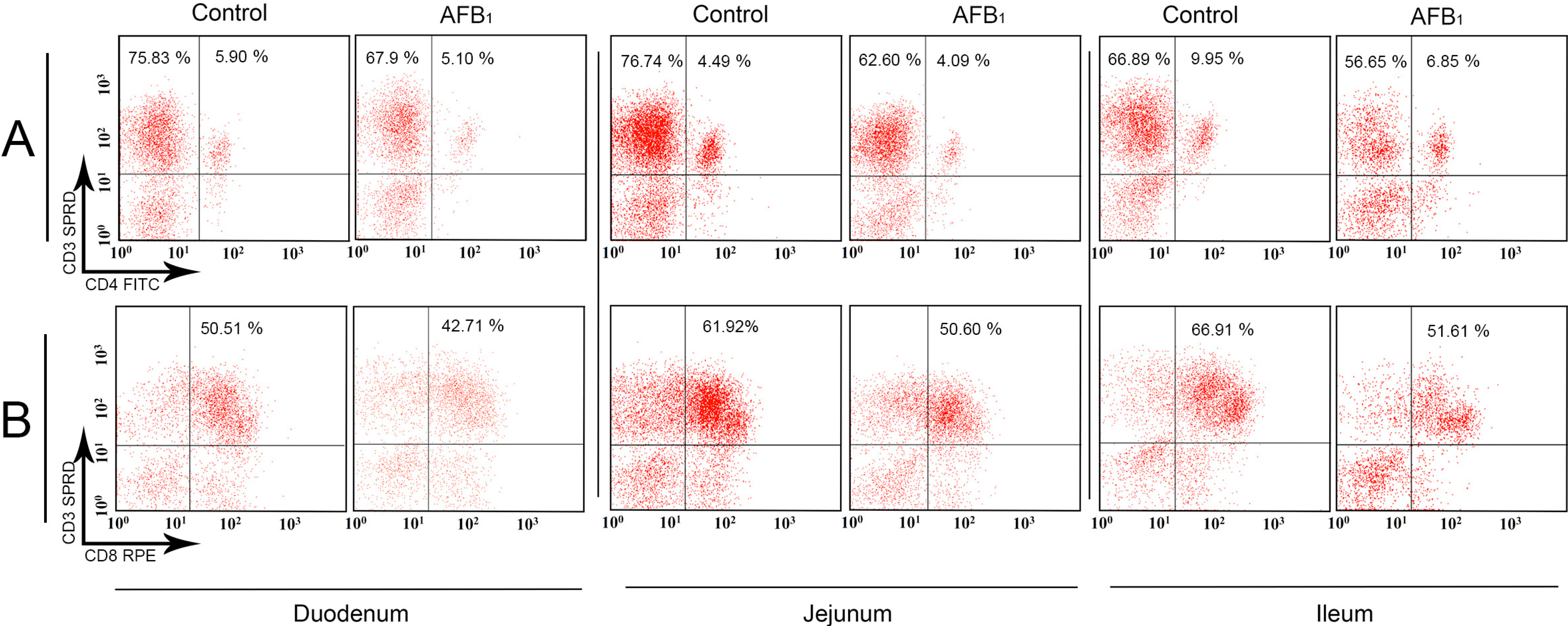
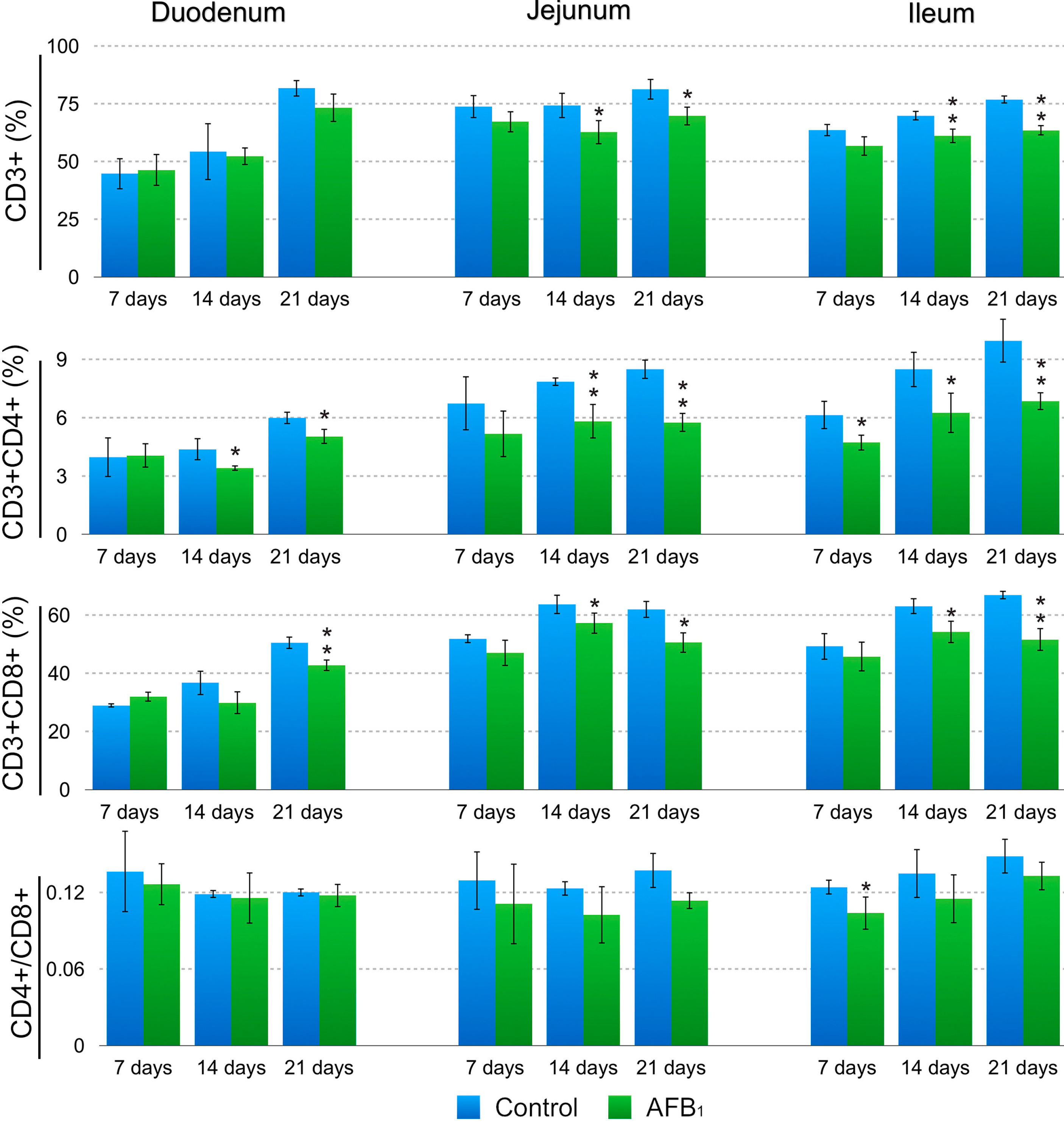
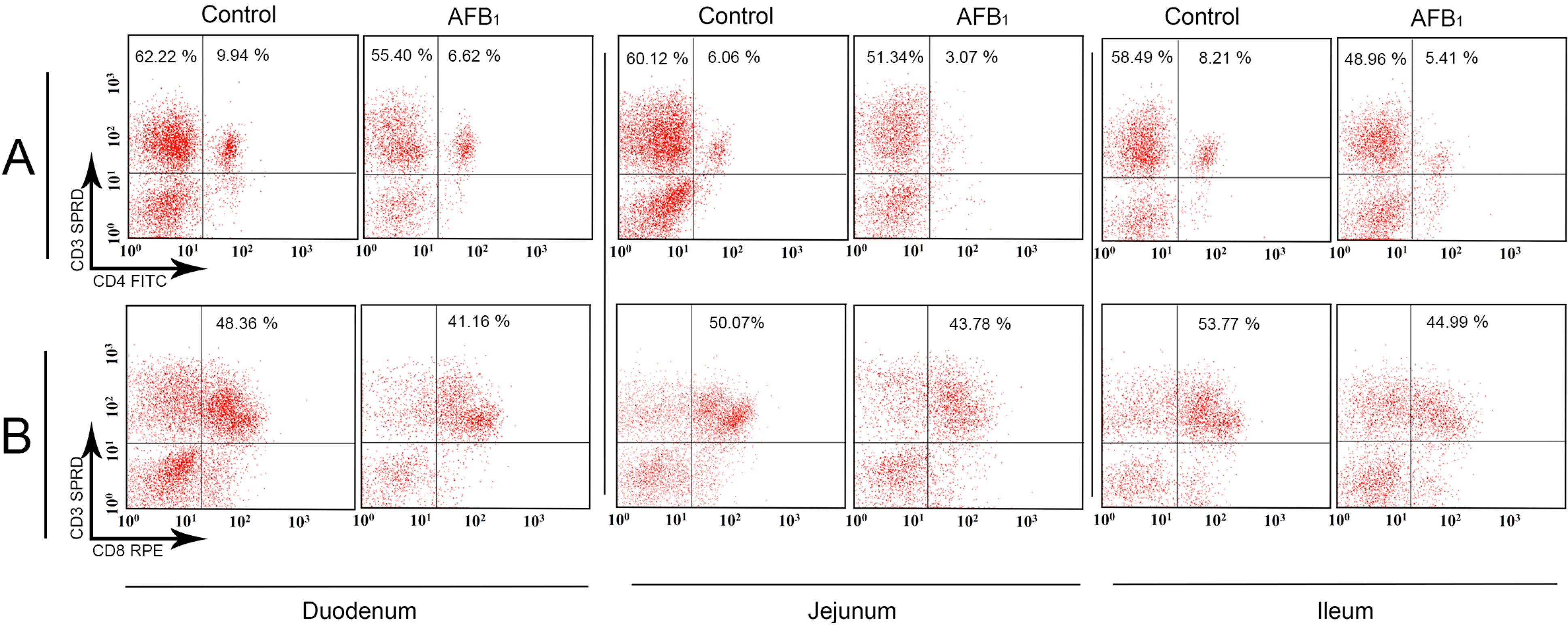
2.1. T-Cell Subsets in the Small Intestinal Intraepithelial Lymphocytes (IELs)
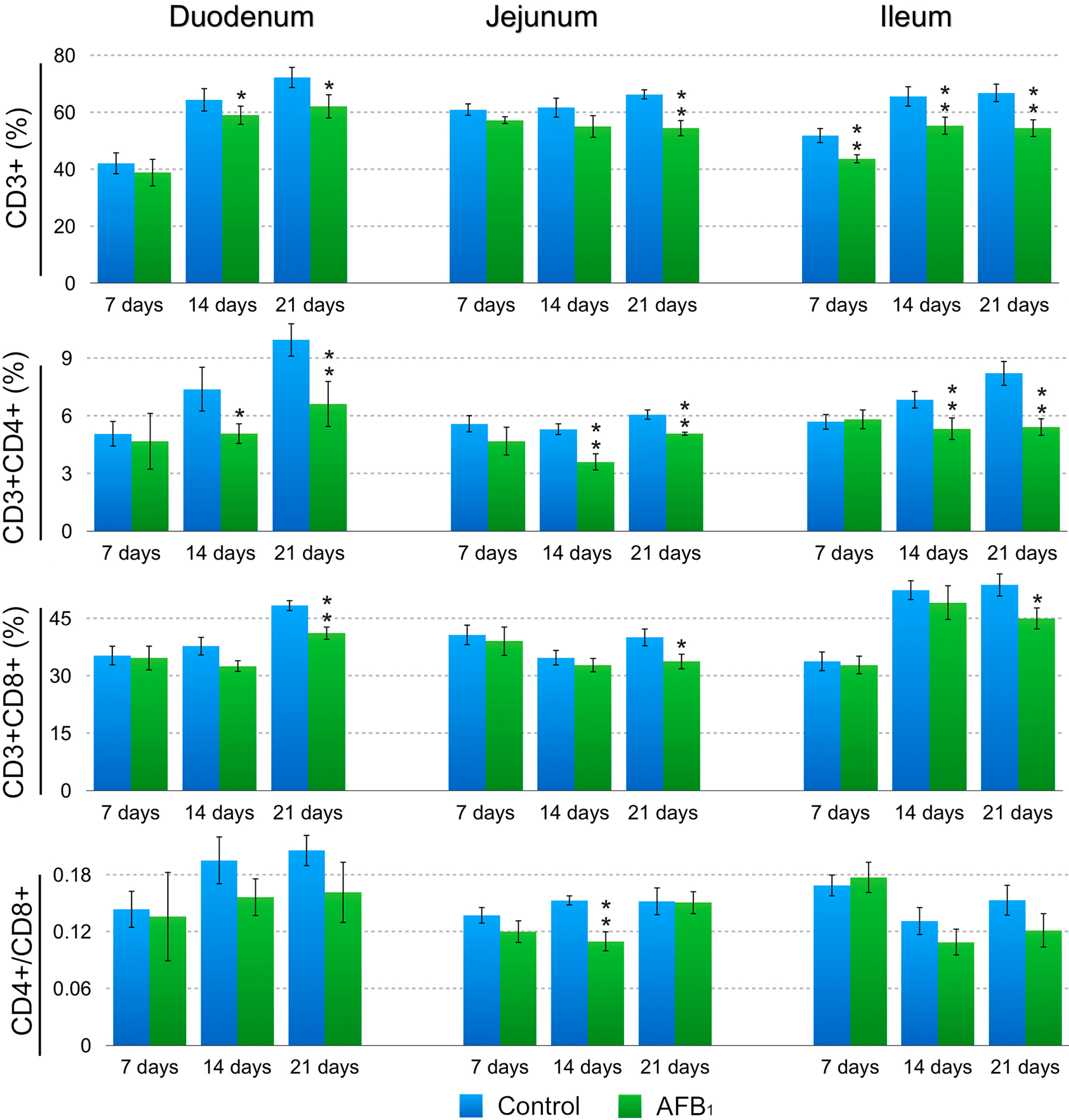
2.2. T-Cell Subsets in the Small Intestinal Lamina Propria Lymphocytes (LPLs)
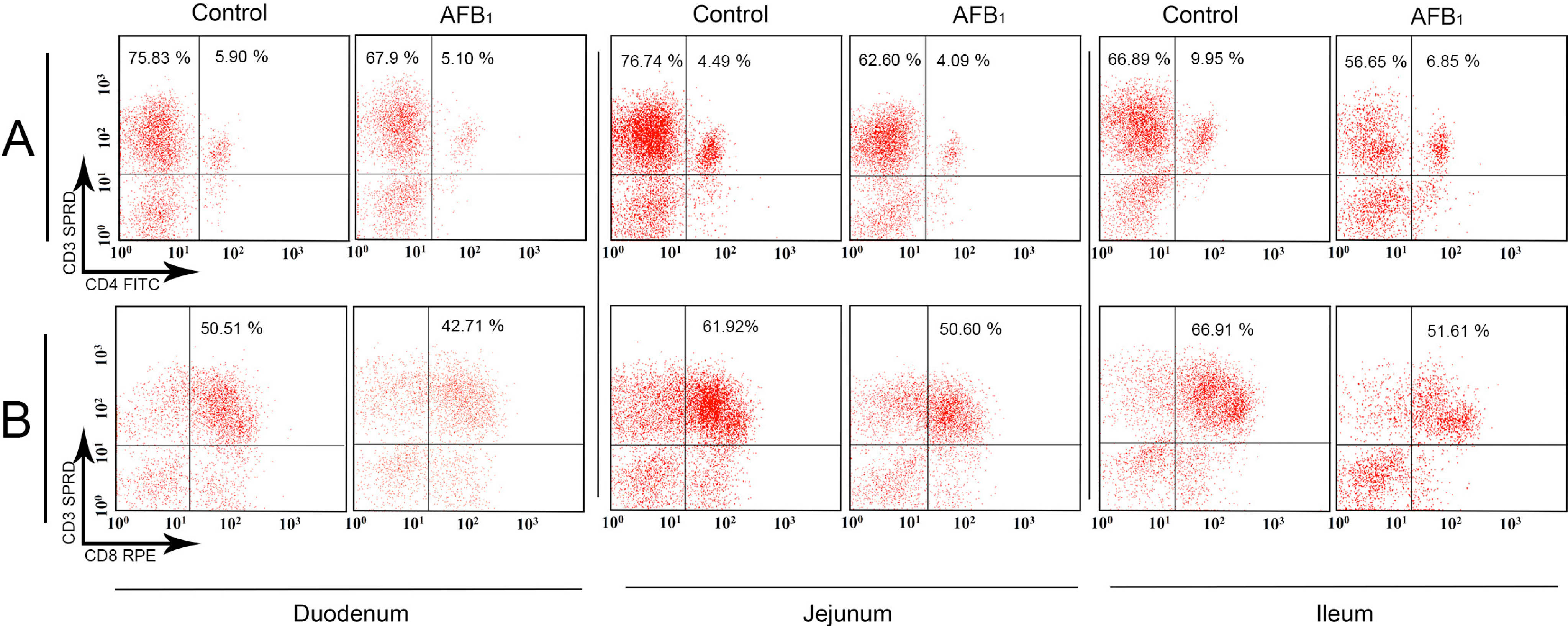

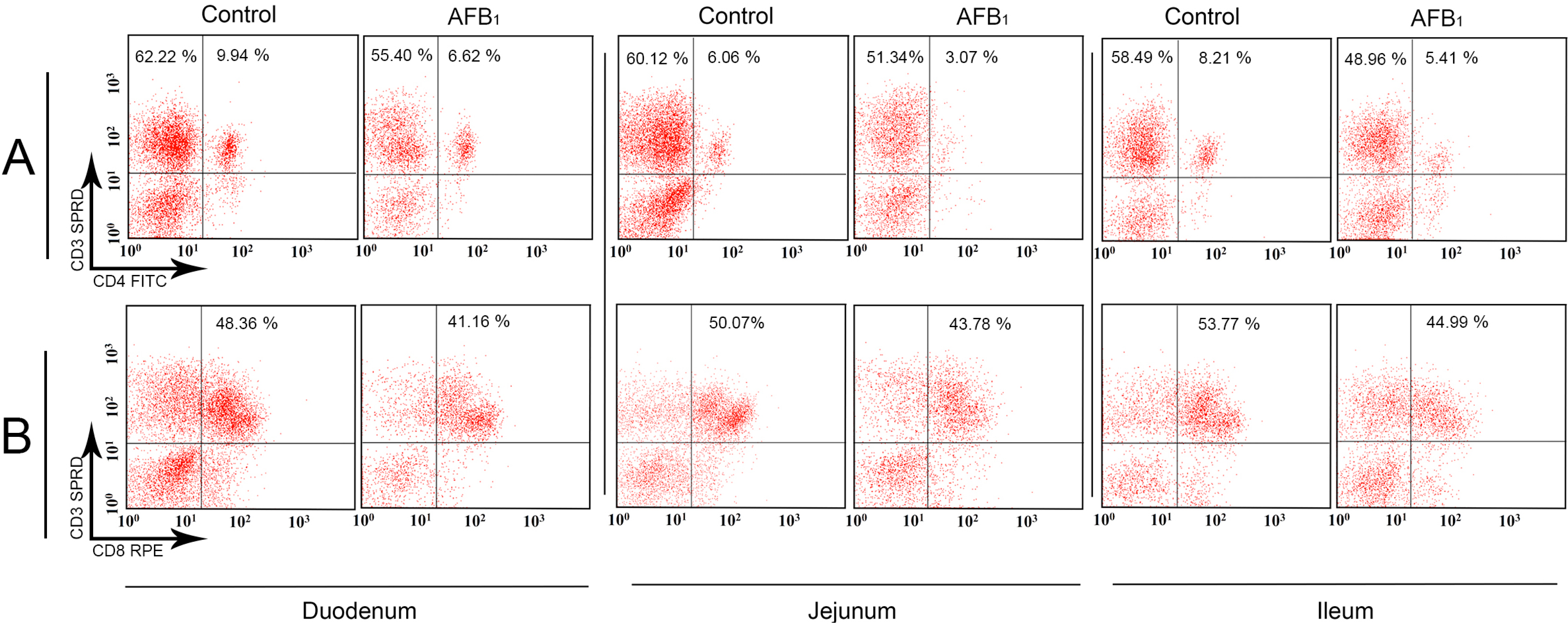

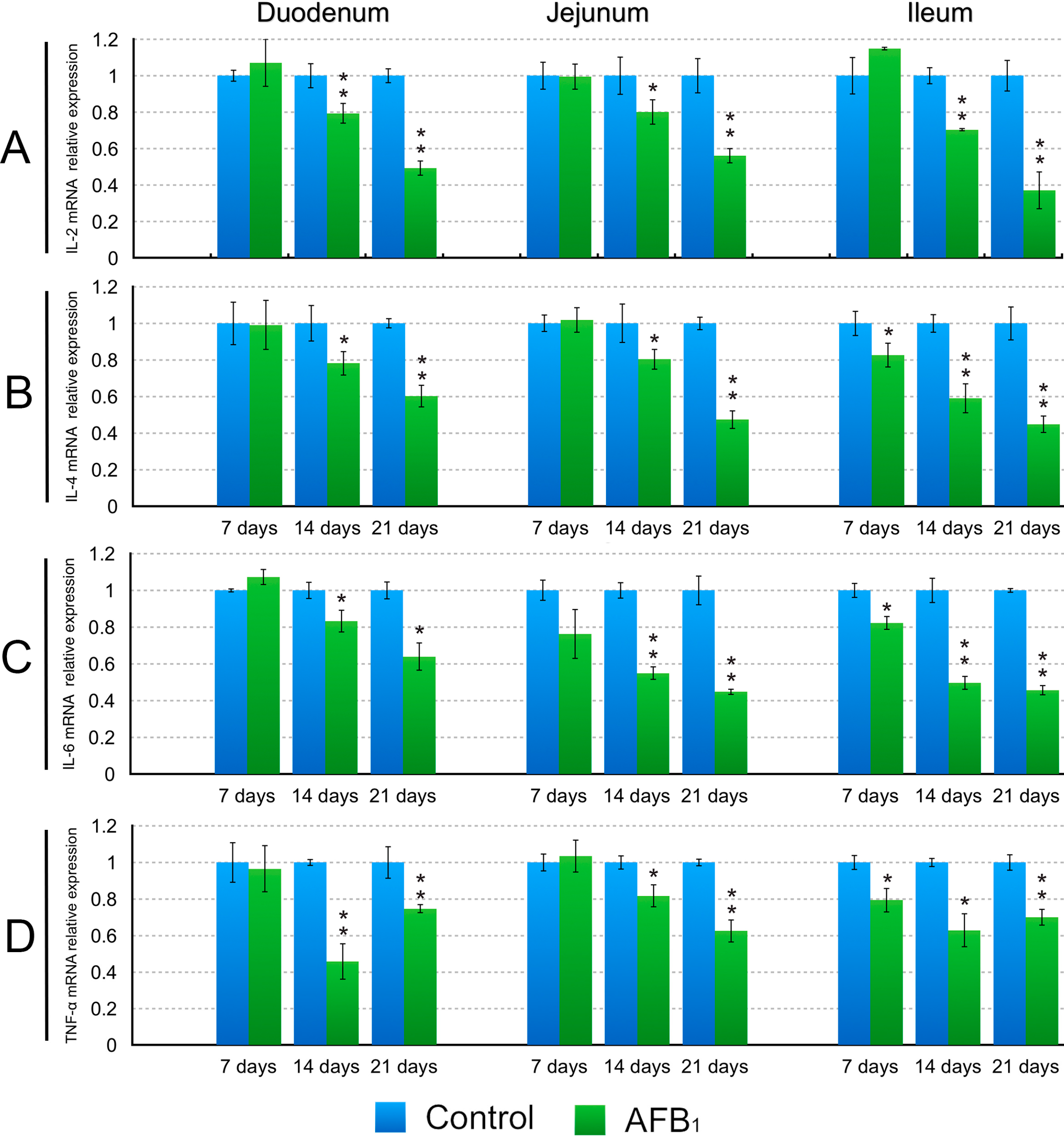
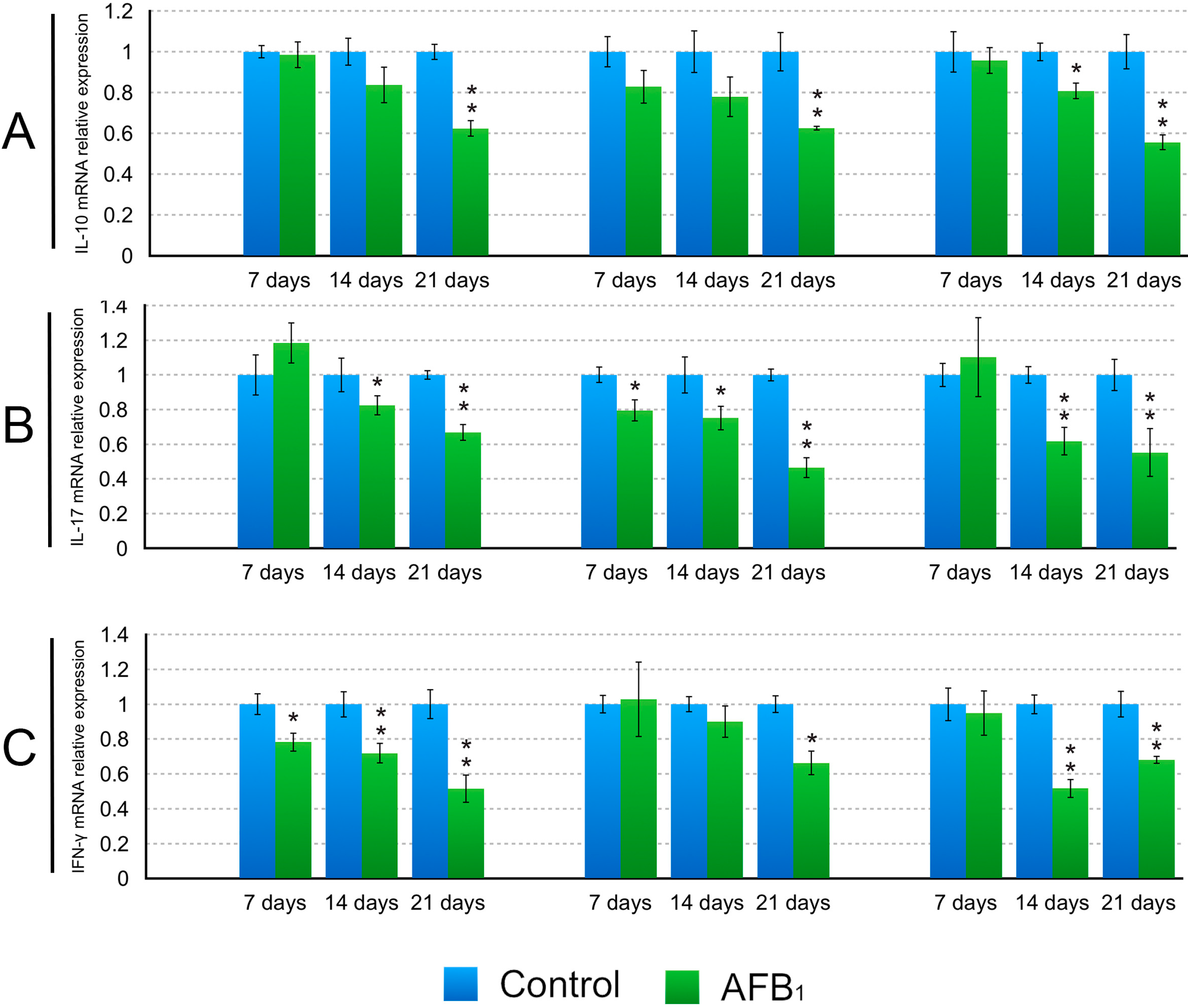
2.3. The Expression Levels of the IL-2, IL-4, IL-6, IL-10, IL-17, IFN-γ and TNF-α (LITAF) mRNA in the Small Intestinal Mucosa

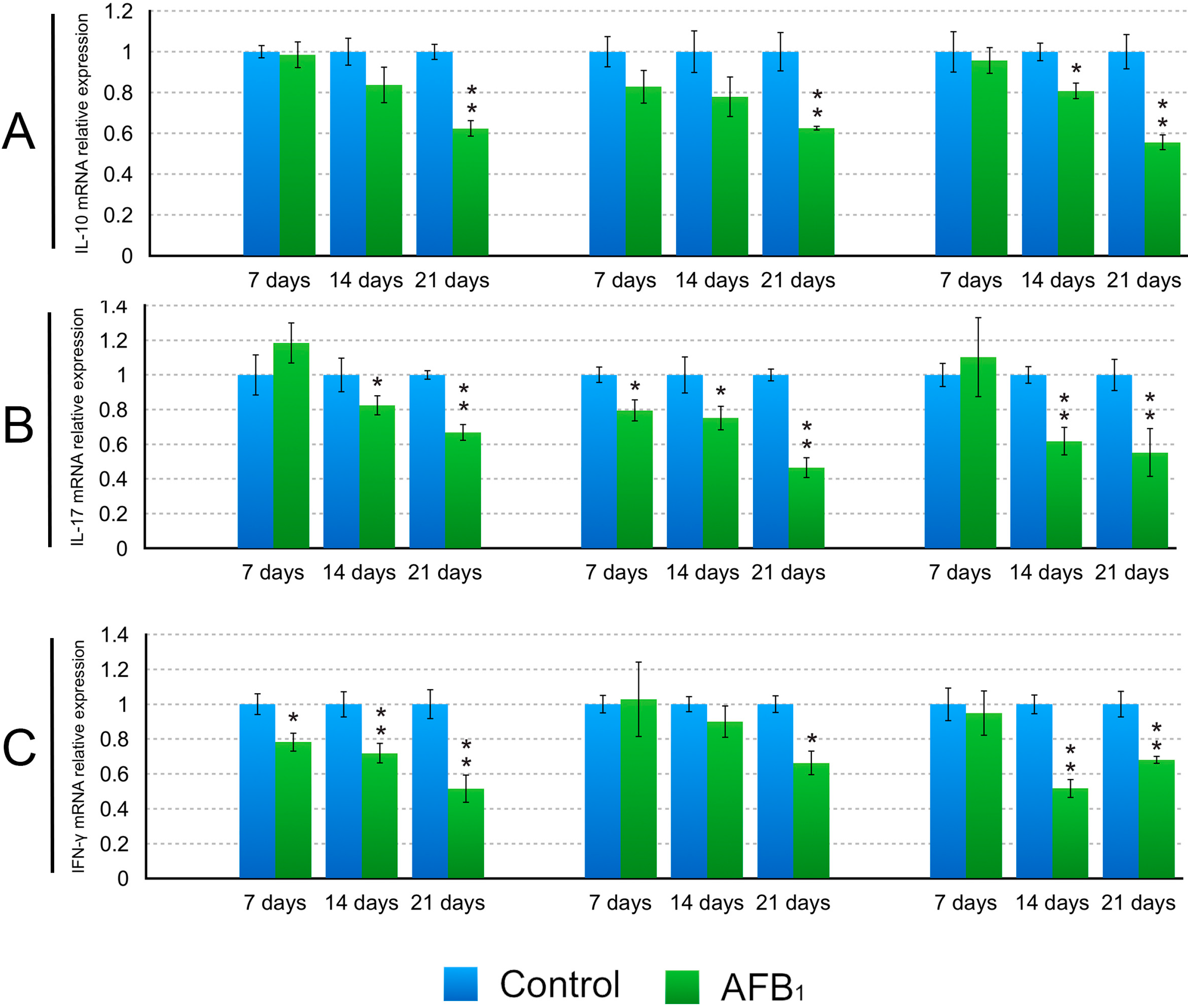
3. Discussion
4. Experimental Section
4.1. Animals and Diets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Content (%) | Nutrient | Content (%) |
|---|---|---|---|
| Corn | 51.95 | Crude protein (CP) | 21.50 |
| Soybean | 39.50 | Methionine (Met) | 0.50 |
| Rapeseed oil | 4.10 | Calcium (Ca) | 1.00 |
| dl-Metionine | 0.20 | All phosphorus (P) | 0.70 |
| Calcium hydrogen phosphate | 1.85 | Methionine + cysteine (Met + Cys) | 0.84 |
| Calcium carbonate | 1.30 | Lysine (Lys) | 1.15 |
| Sodium chloride | 0.40 | Threonine (Thr) | 0.83 |
| Trace element premix a | 0.50 | Metabolizable energy (ME) (MJ/Kg) | 29.90 |
| Choline | 0.17 | ||
| Multivitamins b | 0.03 | ||
| Total | 100 |
4.2. Determination of the Proportion of T-Cell Subsets in the Intestine by FCM
4.2.1. Isolation of the Intra-Epithelial Lymphocytes (IELs)
4.2.2. Isolation of Lamina Propria Lymphocytes (LPLs)
4.2.3. Flow Cytometry (FCM) Method
4.3. Determination of Cytokines mRNA Expression Levels in the Intestinal Mucosa by qRT-PCR
| Gene | Primer | Sequences (5'–3') | Product Size (bp) | Accession Number |
|---|---|---|---|---|
| IL-2 | F | CCCGTGGCTAACTAATCTGC | 114 | AF000631 |
| R | TTGAGCCCGTAGGTTACAGAA | |||
| IL-4 | F | TCTTCCTCAACATGCGTCAG | 108 | NM_1007079.1 |
| R | GGTCTGCTAGGAACTTCTCCAT | |||
| IL-6 | F | CAAGGTGACGGAGGAGGAC | 254 | AJ309540 |
| R | TGGCGAGGAGGGATTTCT | |||
| IL-10 | F | CGGGAGCTGAGGTGAA | 272 | AJ621614 |
| R | GTGAAGAAGCGGTGACAGC | |||
| IL-17 | F | CAGATGCTGGATGCCTAACC | 123 | AJ493595 |
| R | CCAGTGAGCGTTTGCTGATA | |||
| INF-γ | F | AGCTGACGGTGGACCTATTATT | 259 | Y07922 |
| R | GGCTTTGCGCTGGATTC | |||
| TNF-α (LITAF) | F | TGTGTATGTGCAGCAACCCGTAGT | 229 | AY765397 |
| R | GGCATTGCAATTTGGACAGAAGT | |||
| β-Actin | F | TGCTGTGTTCCCATCTATCG | 150 | L08165 |
| R | TTGGTGACAATACCGTGTTCA |
4.4. Statistics
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar]
- Golli-Bennour, E.E.; Kouidhi, B.; Bouslimi, A.; Abid Essefi, S.; Hassen, W.; Bacha, H. Cytotoxicity and genotoxicity induced by aflatoxin B1, ochratoxin A, and their combination in cultured Vero cells. J. Biochem. Mol. Toxicol. 2010, 24, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.; Marin, D.; Galtier, P.; Bertin, G.; Taranu, I.; Oswald, I.; Mengheri, E. Modulation of the immune response by a group of fungal food contaminant, the aflatoxins. In Nutrition and Immunity; Mengheri, E., Roselli, M., Britti, M.S., Finamore, A., Eds.; Research Signpost: Kerala, India, 2006; pp. 147–166. [Google Scholar]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin B1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Raisuddin, S.; Singh, K.; Zaidi, S.; Paul, B.; Ray, P. Immunosuppressive effects of aflatoxin in growing rats. Mycopathologia 1993, 124, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Dugyala, R.R.; Sharma, R.P. The effect of aflatoxin B1 on cytokine mRNA and corresponding protein levels in peritoneal macrophages and splenic lymphocytes. Int. J. Immunopharmacol. 1996, 18, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Meissonnier, G.M.; Pinton, P.; Laffitte, J.; Cossalter, A.M.; Gong, Y.Y.; Wild, C.P.; Bertin, G.; Galtier, P.; Oswald, I.P. Immunotoxicity of aflatoxin B1: Impairment of the cell-mediated response to vaccine antigen and modulation of cytokine expression. Toxicol. Appl. Pharmacol. 2008, 231, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Tomkova, I.; Sevcikova, Z.; Levkut, M.; Revajova, V.; Conkova, E.; Laciakova, A.; Lenhardt, L. Effect of aflatoxin B1 on CD3 T cells and alkaline phosphatase in the intestine of mice. Mycopathologia 2002, 154, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Shu, G.; Peng, X.; Fang, J.; Cui, H.; Chen, J.; Wang, F.; Chen, Z.; Zuo, Z.; Deng, J.; et al. Protective role of sodium selenite on histopathological lesions, decreased T-cell subsets and increased apoptosis of thymus in broilers intoxicated with aflatoxin B1. Food Chem. Toxicol. 2013, 59, 446–454. [Google Scholar]
- Chen, K.; Yuan, S.; Chen, J.; Peng, X.; Wang, F.; Cui, H.; Fang, J. Effects of sodium selenite on the decreased percentage of T cell subsets, contents of serum IL-2 and IFN-γ induced by aflatoxin B1 in broilers. Res. Vet. Sci. 2013, 95, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Peng, X.; Fang, J.; Cui, H.; Zuo, Z.; Deng, J.; Chen, Z.; Geng, Y.; Lai, W.; Tang, L.; et al. Effects of dietary selenium on histopathological changes and T cells of spleen in broilers exposed to aflatoxin B1. Int. J. Environ. Res. Public Health 2014, 11, 1904–1913. [Google Scholar]
- Cukrova, V.; Kurita, N.; Akao, M. An early effect of aflatoxin B1 administered in vivo on the growth of bone marrow CFU-GM and the production of some cytokines in rats. Mycopathologia 1992, 120, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Qian, G.; Tang, L.; Guo, X.; Wang, F.; Massey, M.E.; Su, J.; Guo, T.L.; Williams, J.H.; Phillips, T.D.; Wang, J.S. Aflatoxin B1 modulates the expression of phenotypic markers and cytokines by splenic lymphocytes of male F344 rats. J. Appl. Toxicol. 2014, 34, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Jakab, G.J.; Hmieleski, R.R.; Zarba, A.; Hemenway, D.R.; Groopman, J.D. Respiratory aflatoxicosis: Suppression of pulmonary and systemic host defenses in rats and mice. Toxicol. Appl. Pharmacol. 1994, 125, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.G.; Zhao, L.H.; Wei, H.; Duan, G.X.; Zhang, J.Y.; Ji, C. Effects of lipoic acid on immune function, the antioxidant defense system, and inflammation-related genes expression of broiler chickens fed aflatoxin contaminated diets. Int. J. Mol. Sci. 2014, 15, 5649–5662. [Google Scholar] [CrossRef] [PubMed]
- Bondy, G.S.; Pestka, J.J. Immunomodulation by fungal toxins. J. Toxicol. Environ. Health B 2000, 3, 109–143. [Google Scholar] [CrossRef]
- Lillehoj, H.S.; Chung, K.S. Postnatal development of T-lymphocyte subpopulations in the intestinal intraepithelium and lamina propria in chickens. Vet. Immunol. Immunopathol. 1992, 31, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Wittig, B.; Zeitz, M. The gut as an organ of immunology. Int. J. Colorectal. Dis. 2003, 18, 181–187. [Google Scholar] [PubMed]
- Blaschitz, C.; Raffatellu, M. Th17 cytokines and the gut mucosal barrier. J. Clin. Immunol. 2010, 30, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Kolars, J.C.; Benedict, P.; Schmiedlin Ren, P.; Watkins, P.B. Aflatoxin B1-adduct formation in rat and human small bowel enterocytes. Gastroenterology 1994, 106, 433–439. [Google Scholar] [PubMed]
- Huang, A.; Shibata, E.; Nishimura, H.; Igarashi, Y.; Isobe, N.; Yoshimura, Y. Effects of probiotics on the localization of T cell subsets in the intestine of broiler chicks. J. Poult. Sci. 2013, 50, 275–281. [Google Scholar] [CrossRef]
- Yarru, L.; Settivari, R.; Gowda, N.; Antoniou, E.; Ledoux, D.; Rottinghaus, G. Effects of turmeric (Curcuma longa) on the expression of hepatic genes associated with biotransformation, antioxidant, and immune systems in broiler chicks fed aflatoxin. Poult. Sci. 2009, 88, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.; Taranu, I.; Bunaciu, R.; Pascale, F.; Tudor, D.; Avram, N.; Sarca, M.; Cureu, I.; Criste, R.; Suta, V. Changes in performance, blood parameters, humoral and cellular immune responses in weanling piglets exposed to low doses of aflatoxin. J. Anim. Sci. 2002, 80, 1250–1257. [Google Scholar] [PubMed]
- Bruneau, J.C.; Stack, E.; O’Kennedy, R.; Loscher, C.E. Aflatoxins B1, B2 and G1 modulate cytokine secretion and cell surface marker expression in J774A. 1 murine macrophages. Toxicol. In Vitro 2012, 26, 686–693. [Google Scholar] [CrossRef]
- Girish, C.; Smith, T. Impact of feed-borne mycotoxins on avian cell-mediated and humoral immune responses. World Mycotoxin J. 2008, 1, 105–121. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, C.; Mattner, J.; Schleicher, U. The role of type I interferons in non-viral infections. Immunol. Rev. 2004, 202, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.L.; Hwang, J.B.; Park, J.J.; Kim, S.G. Expression of transforming growth factor β1, transforming growth factor type I and II receptors, and TNF-α in the mucosa of the small intestine in infants with food protein–induced enterocolitis syndrome. J. Allergy Clin. Immunol. 2002, 109, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, C.; Durual, S.; Estienne, M.; Bouzakri, K.; Heim, M.H.; Blin, N.; Cuber, J.C. IL-4 and IL-13 up-regulate intestinal trefoil factor expression: Requirement for STAT6 and de novo protein synthesis. J. Immunol. 2004, 172, 3775–3783. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Hirano, T.; Taga, T.; Kishimoto, T. Biology of multifunctional cytokines: IL 6 and related molecules (IL1 and TNF). FASEB J. 1990, 4, 2860–2867. [Google Scholar] [PubMed]
- Rothwell, L.; Young, J.R.; Zoorob, R.; Whittaker, C.A.; Hesketh, P.; Archer, A.; Smith, A.L.; Kaiser, P. Cloning and characterization of chicken IL-10 and its role in the immune response to Eimeria maxima. J. Immunol. 2004, 173, 2675–2682. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Martin, M.T.; Targan, S.R. Regulation of immune responses of the intestinal mucosa. Crit. Rev. Immunol. 1996, 16, 277–309. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in affecting broiler’s performance, immunity, and gastrointestinal tract: A review of history and contemporary issues. Toxins (Basel) 2011, 3, 566–590. [Google Scholar] [CrossRef]
- Yunus, A.; Ghareeb, K.; Abd-El-Fattah, A.; Twaruzek, M.; Böhm, J. Gross intestinal adaptations in relation to broiler performance during chronic aflatoxin exposure. Poult. Sci. 2011, 90, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Kaoud, H.A. Innovative methods for the amelioration of aflatoxin (AFB1) effect in broiler chicks. Sci. J. Appl. Res. 2013, 1, 15–19. [Google Scholar]
- Montufar-Solis, D.; Klein, J.R. An improved method for isolating intraepithelial lymphocytes (IELs) from the murine small intestine with consistently high purity. J. Immunol. Methods 2006, 308, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Resendiz-Albor, A.A.; Esquivel, R.; Lopez Revilla, R.; Verdin, L.; Moreno-Fierros, L. Striking phenotypic and functional differences in lamina propria lymphocytes from the large and small intestine of mice. Life Sci. 2005, 76, 2783–2803. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Kaiser, P. Effects of stress, mimicked by administration of corticosterone in drinking water, on the expression of chicken cytokine and chemokine genes in lymphocytes. Stress 2009, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.H.; Lillehoj, H.S.; Lillehoj, E.P.; Lee, S.H. Changes in immune-related gene expression and intestinal lymphocyte subpopulations following Eimeria maxima infection of chickens. Vet. Immunol. Immunopathol. 2006, 114, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Peng, X.; Fang, J.; Cui, H.; Yu, Z.; Chen, Z. Effects of Aflatoxin B1 on T-Cell Subsets and mRNA Expression of Cytokines in the Intestine of Broilers. Int. J. Mol. Sci. 2015, 16, 6945-6959. https://doi.org/10.3390/ijms16046945
Jiang M, Peng X, Fang J, Cui H, Yu Z, Chen Z. Effects of Aflatoxin B1 on T-Cell Subsets and mRNA Expression of Cytokines in the Intestine of Broilers. International Journal of Molecular Sciences. 2015; 16(4):6945-6959. https://doi.org/10.3390/ijms16046945
Chicago/Turabian StyleJiang, Min, Xi Peng, Jing Fang, Hengmin Cui, Zhengqiang Yu, and Zhengli Chen. 2015. "Effects of Aflatoxin B1 on T-Cell Subsets and mRNA Expression of Cytokines in the Intestine of Broilers" International Journal of Molecular Sciences 16, no. 4: 6945-6959. https://doi.org/10.3390/ijms16046945