
Hepatitis A Virus Genotype Distribution during a Decade of Universal Vaccination of Preadolescents


 ,
,  and
and 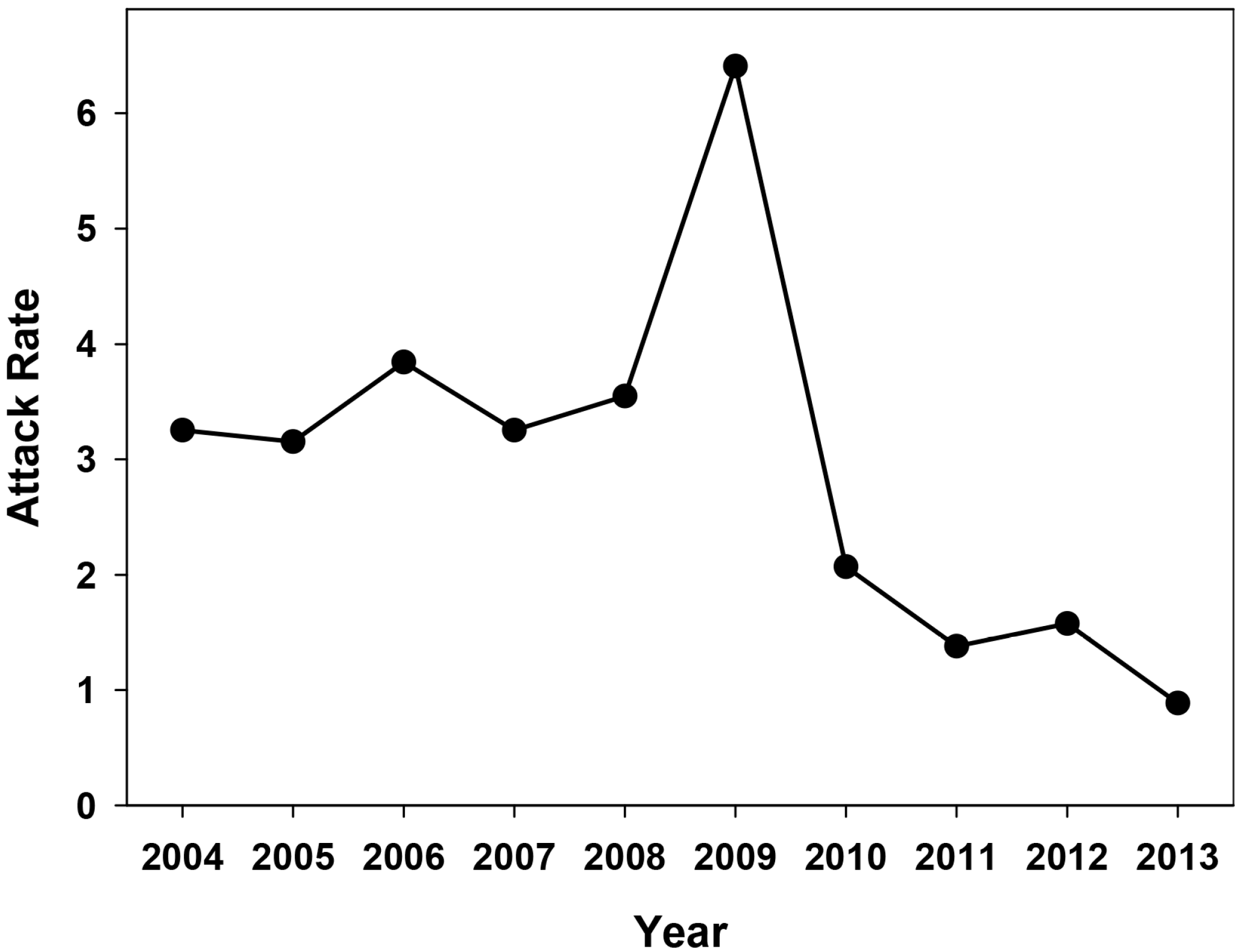{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
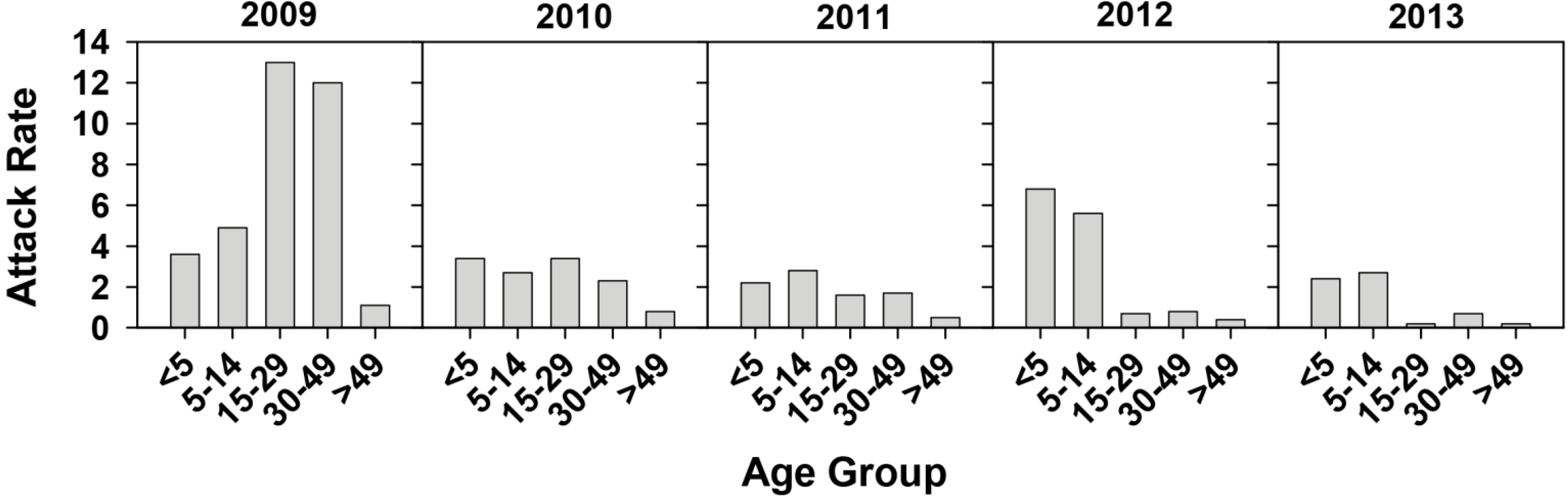
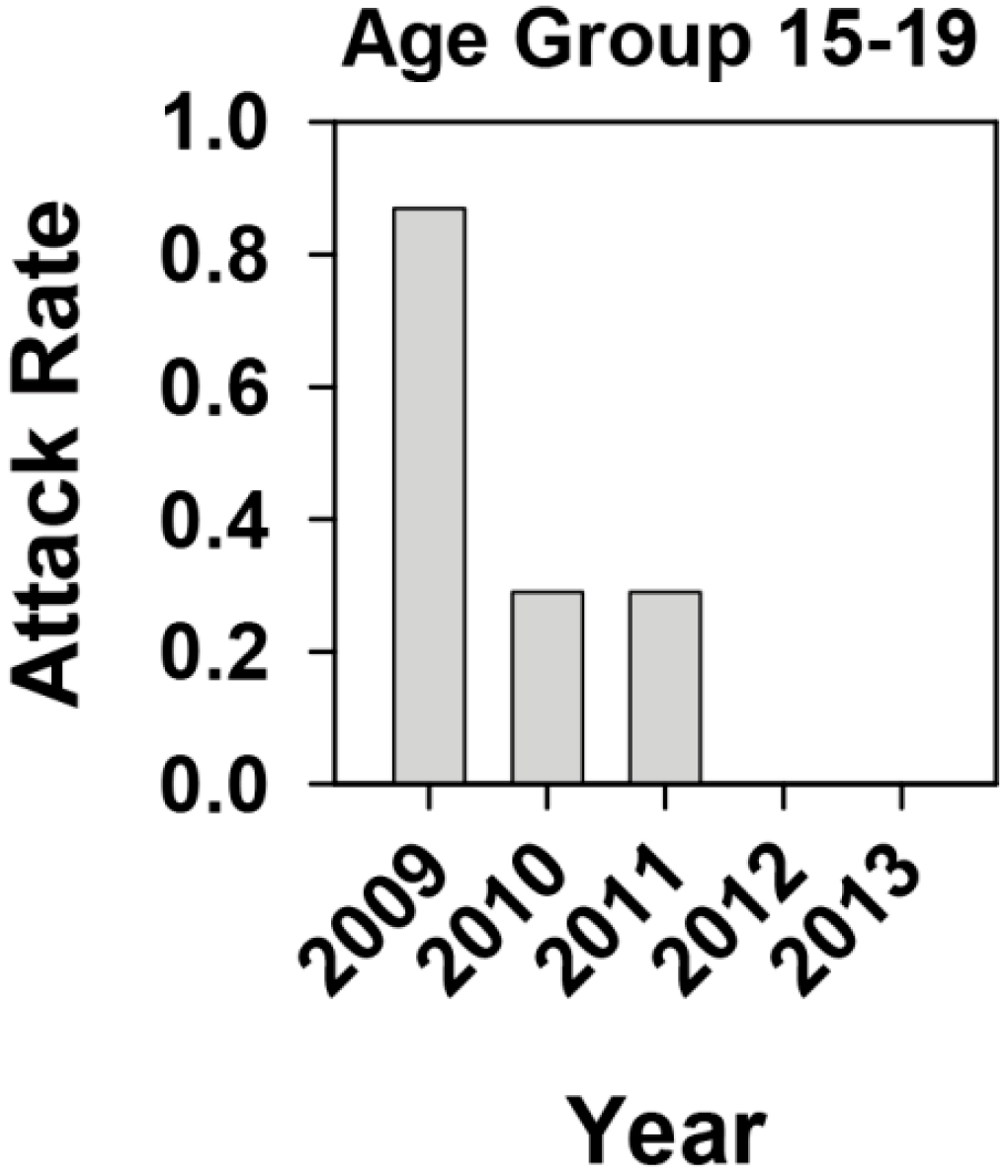
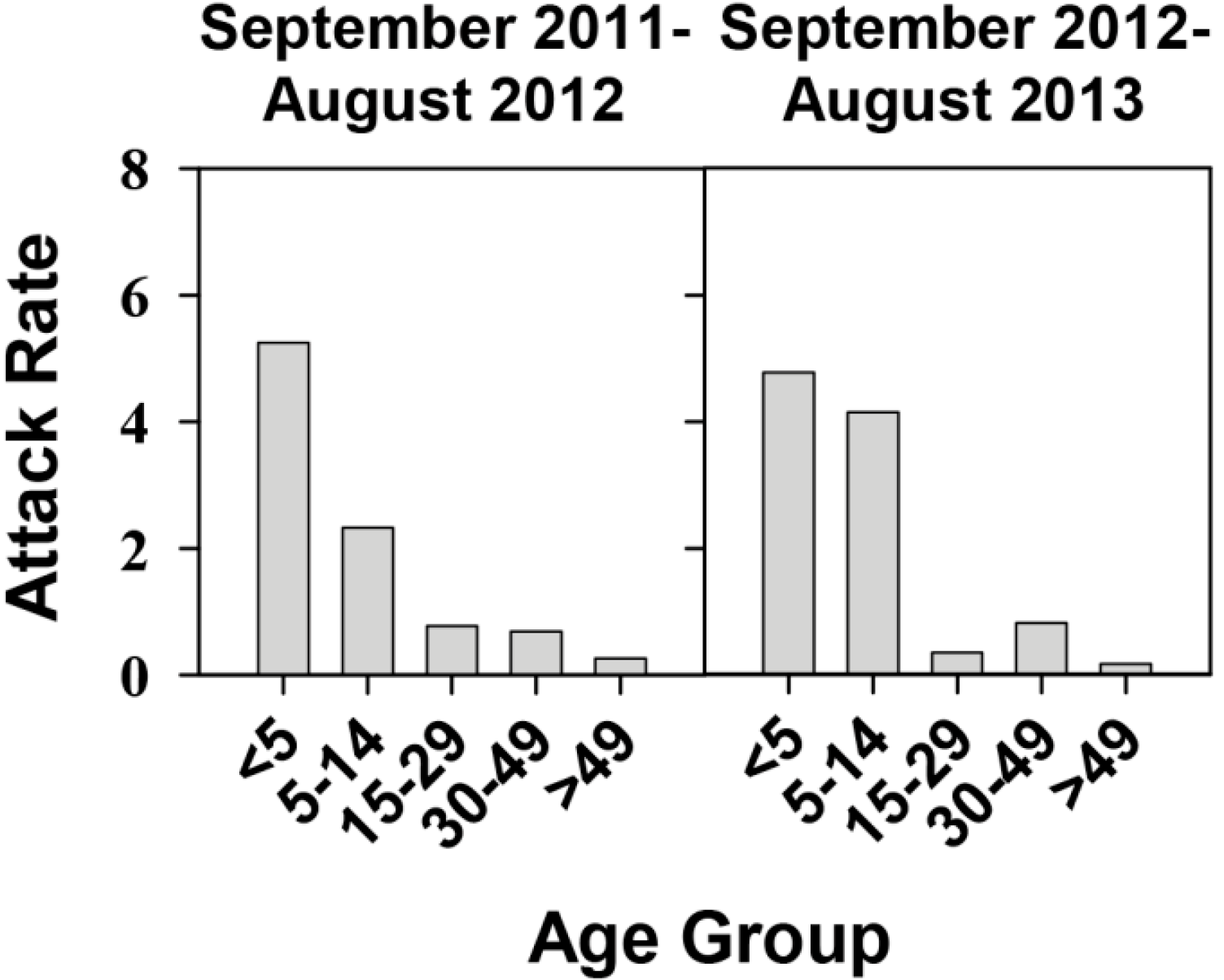
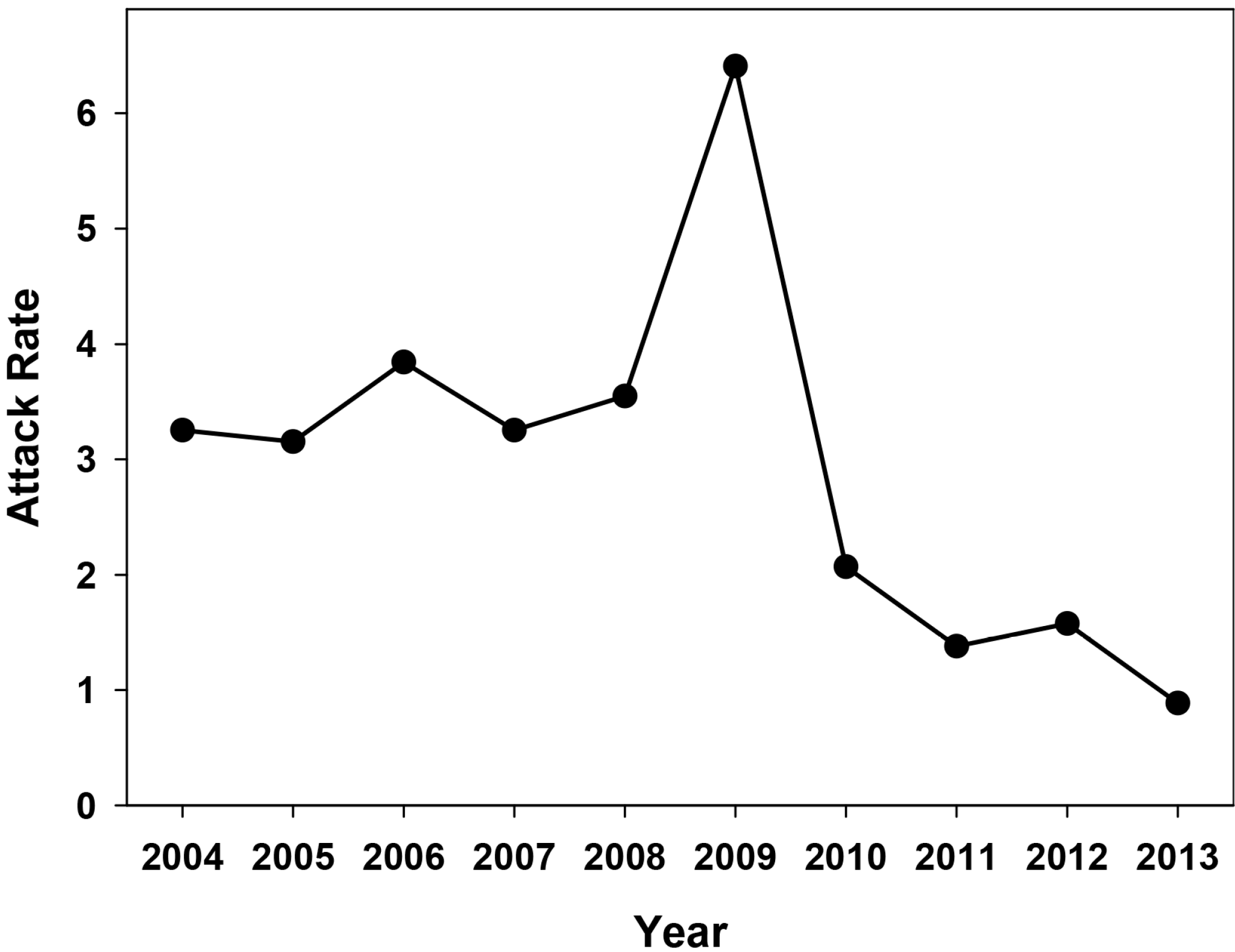
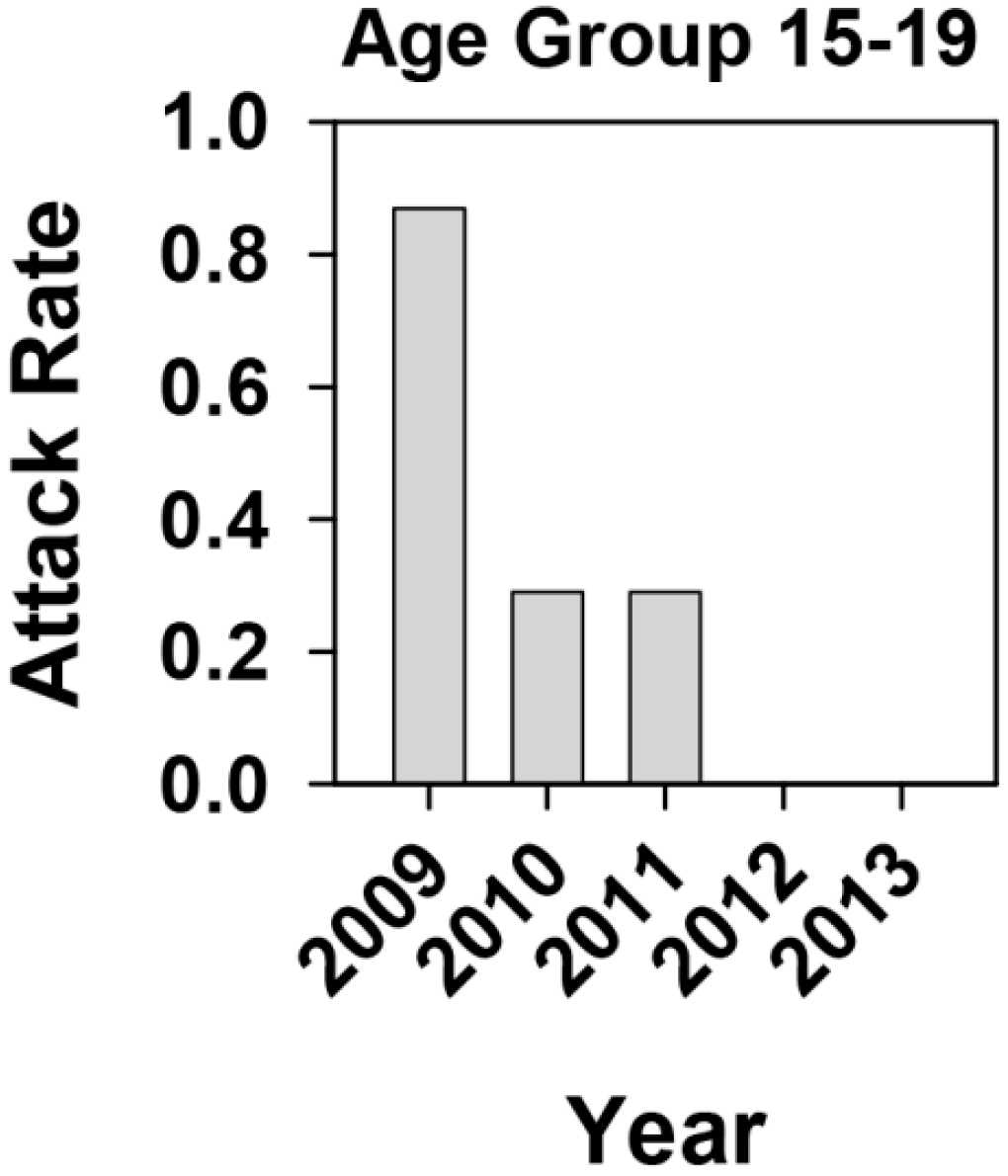
2.1. Effectiveness of the Universal Vaccination Campaign Revealed through the Evolution of the Global and Age-Group Specific Attack Rates

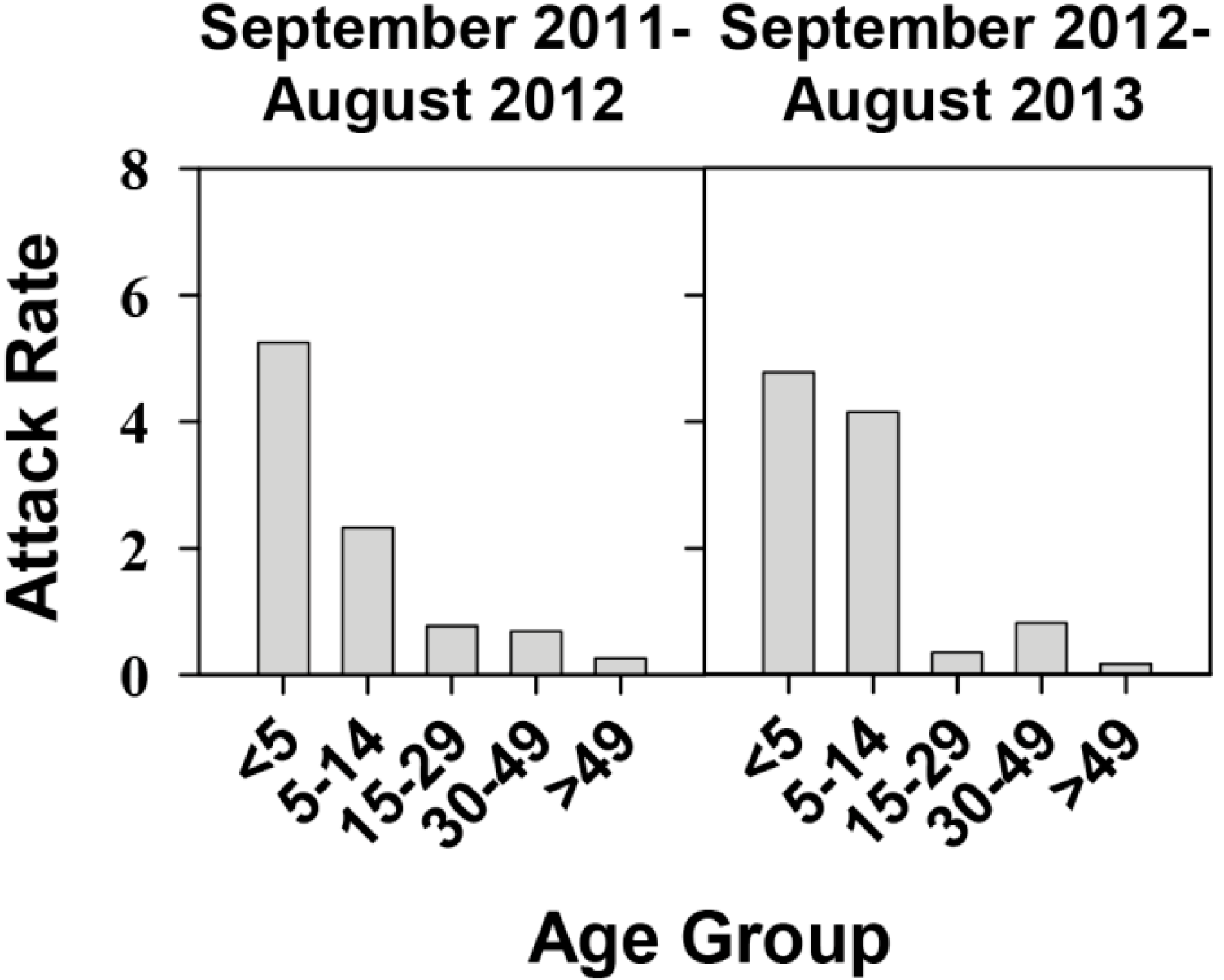
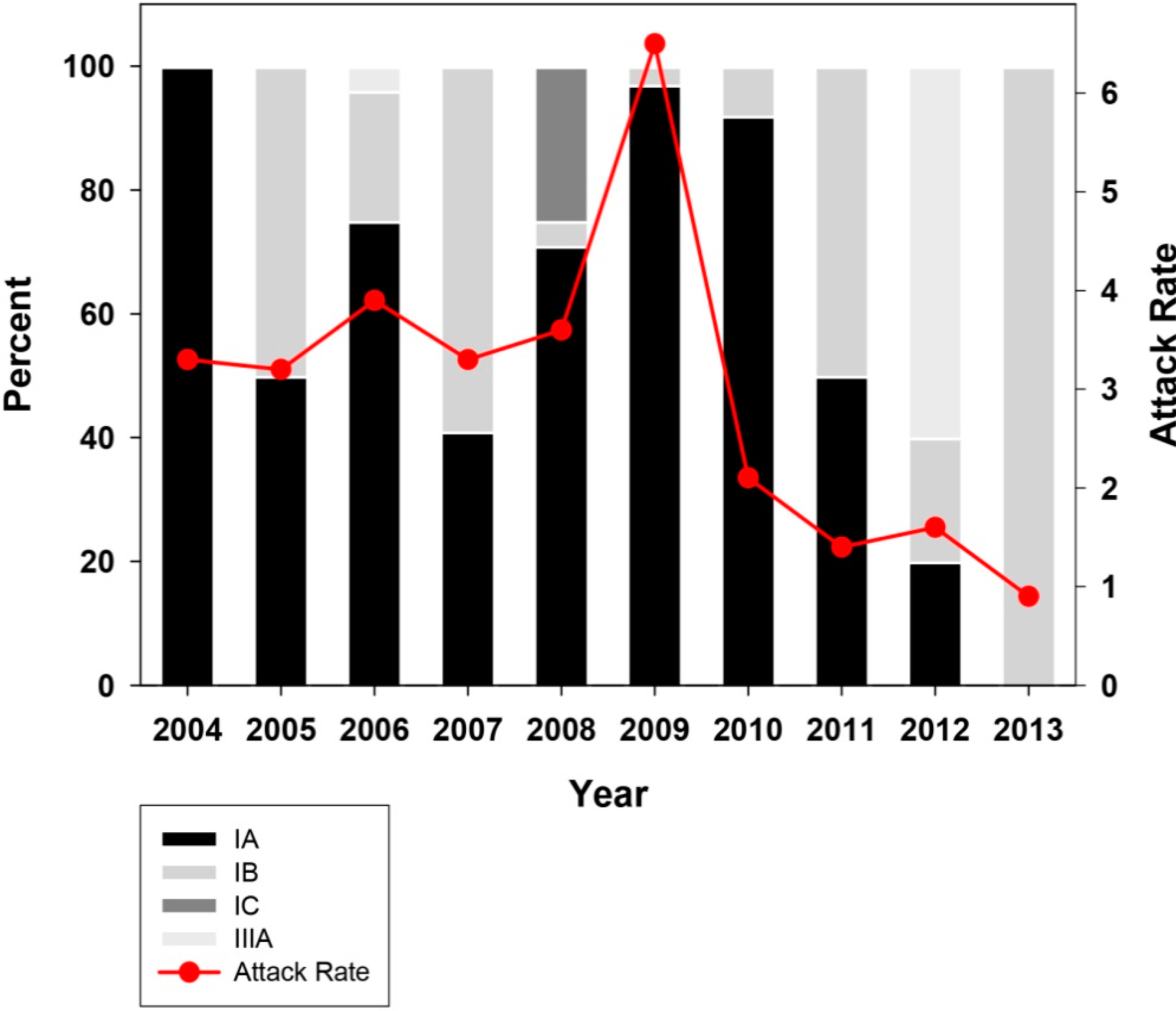
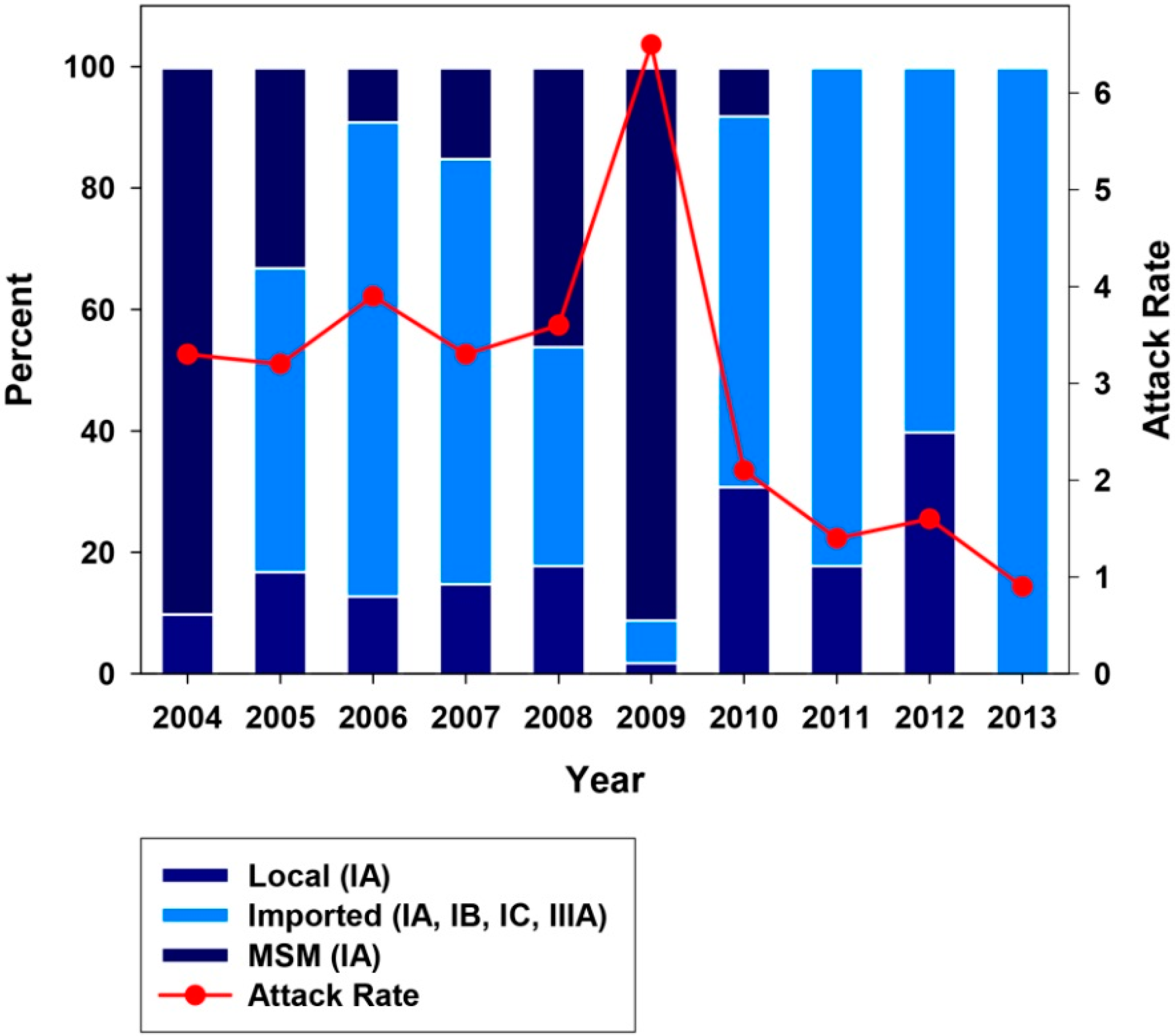
2.2. Genotype Analysis

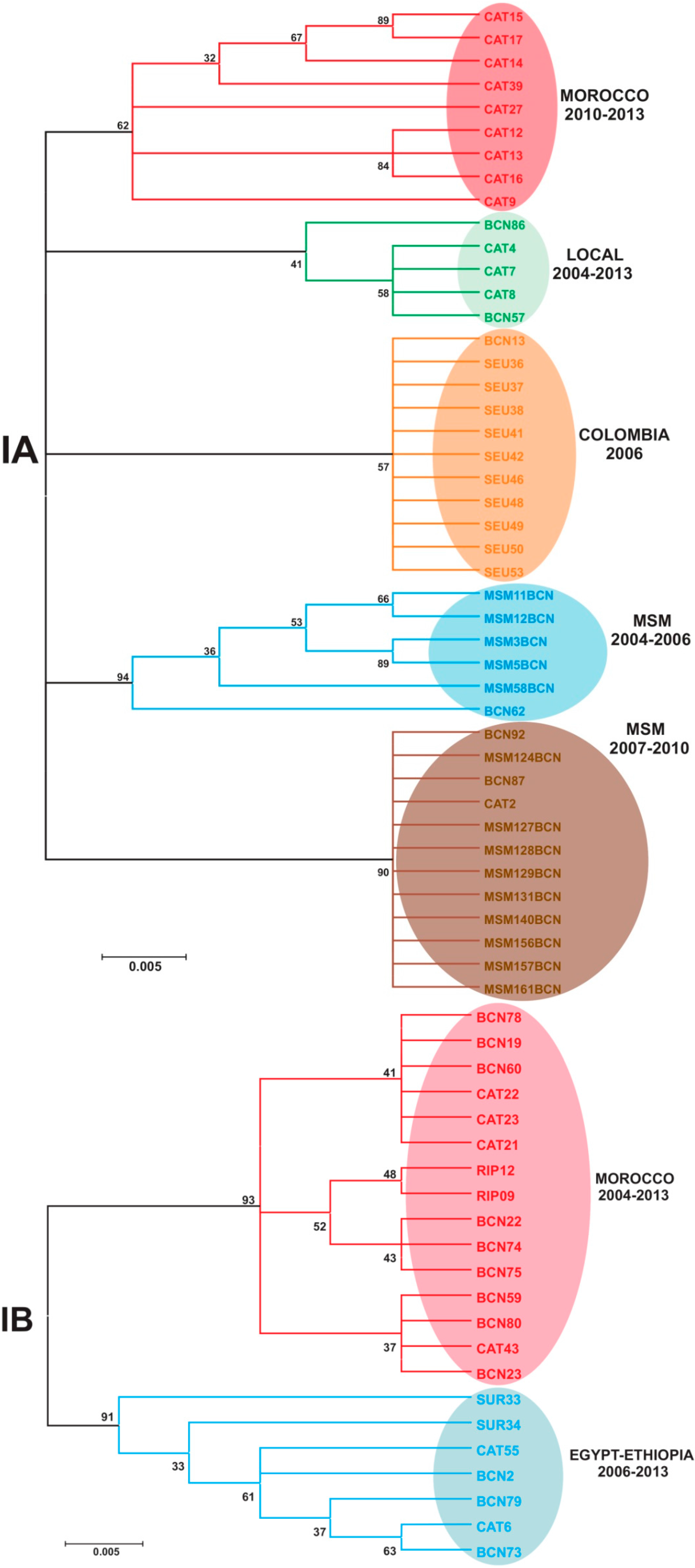

3. Experimental Section
3.1. Estimation of the Attack Rates
3.2. Epidemiological Data and Genotype Determination
3.3. Accession Numbers
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pérez-Sautu, U.; Costafreda, M.I.; Lite, J.; Sala, R.; Barrabeig, I.; Bosch, A.; Pintó, R.M. Molecular epidemiology of hepatitis A virus infections in Catalonia, Spain, 2005–2009: Circulation of newly emerging strains. J. Clin. Virol. 2011, 52, 98–102. [Google Scholar] [CrossRef]
- Faber, M.S.; Stark, K.; Behnke, S.C.; Schreier, E.; Frank, C. Epidemiology of hepatitis A virus infections, Germany, 2007–2008. Emerg. Infect. Dis. 2009, 15, 1760–1768. [Google Scholar] [CrossRef] [PubMed]
- Klevens, R.; Miller, J.T.; Iqbal, K.; Thomas, A.; Rizzo, E.M.; Hanson, H.; Sweet, K.; Phan, Q.; Cronquist, A.; Khudyakov, Y.; et al. The evolving epidemiology of hepatitis A in the United States: Incidence and molecular epidemiology from population-based surveillance, 2005–2007. Arch. Intern. Med. 2010, 170, 1811–1818. [Google Scholar]
- Stene-Johansen, K.; Tjon, G.; Schreier, E.; Bremer, V.; Bruisten, S.; Ngui, S.L.; King, M.; Pintó, R.M.; Aragonè, L.; Mazick, A.; et al. Molecular epidemiological studies show that hepatitis A virus is endemic among active homosexual men in europe. J. Med. Virol. 2007, 79, 356–365. [Google Scholar]
- Tortajada, C.; de Olalla, P.; Díez, E.; Pintó, R.; Bosch, A.; Pérez, U.; Sanz, M.; Cayla, J. Hepatitis A among men who have sex with men in barcelona, 1989–2010: Insufficient control and need for new approaches. BMC Infect. Dis. 2012, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Stene-Johansen, K.; Jenum, P.A.; Hoel, T.; Blystad, H.; Sunde, H.; Skaug, K. An outbreak of hepatitis A among homosexuals linked to a family outbreak. Epidemiol. Infect. 2002, 129, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Pintó, R.M.; D’Andrea, L.; Pérez-Rodríguez, F.J.; Costafreda, M.I.; Ribes, E.; Guix, S.; Bosch, A. Hepatitis A virus evolution and the potential emergence of new variants escaping the presently available vaccines. Future Microbiol. 2012, 7, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, A.; Oviedo, M.; Carmona, G.; Batalla, J.; Bruguera, M.; Salleras, L.; Plasencia, A. Impact and effectiveness of a mass hepatitis A vaccination programme of preadolescents seven years after introduction. Vaccine 2008, 26, 1737–1741. [Google Scholar] [CrossRef] [PubMed]
- Tortajada, C.; de Olalla, P.G.; Pintó, R.M.; Bosch, A.; Cayla, J. Outbreak of hepatitis A among men who have sex with men in barcelona, spain, september 2008–march 2009. Euro Surveill. 2009, 14, 2–4. [Google Scholar] [PubMed]
- Pintó, R.M.; Costafreda, M.I.; Bosch, A. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sautu, U.; Costafreda, M.I.; Cayla, J.; Tortajada, C.; Lite, J.; Bosch, A.; Pintó, R.M. Hepatitis A virus vaccine escape variants and potential new serotype emergence. Emerg. Infect. Dis. 2011, 17, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Aragonès, L.; Bosch, A.; Pintó, R.M. Hepatitis a virus mutant spectra under the selective pressure of monoclonal antibodies: Codon usage constraints limit capsid variability. J. Virol. 2008, 82, 1688–1700. [Google Scholar] [CrossRef] [PubMed]
- Costafreda, M.I.; Ribes, E.; Franch, À; Bosch, A.; Pintó, R.M. A single mutation in the glycophorin a binding site of hepatitis a virus enhances virus clearance from the blood and results in a lower fitness variant. J. Virol. 2012, 86, 7887–7895. [Google Scholar]
- Sánchez, G.; Aragonès, L.; Costafreda, M.I.; Ribes, E.; Bosch, A.; Pintó, R.M. Capsid region involved in hepatitis a virus binding to glycophorin a of the erythrocyte membrane. J. Virol. 2004, 78, 9807–9813. [Google Scholar] [CrossRef] [PubMed]
- Bonanni, P.; Bechini, A.; Pesavento, G.; Guadagno, R.; Santini, M.G.; Baretti, S.; Bartoloni, A.; Taliani, G. Primary hepatitis A vaccination failure is a rare although possible event: Results of a retrospective study. Vaccine 2006, 24, 6053–6057. [Google Scholar] [CrossRef] [PubMed]
- Desbois, D.; Couturier, E.; Mackiewicz, V.; Graube, A.; Letort, M.J.; Dussaix, E.; Roque-Afonso, A.M. Epidemiology and genetic characterization of hepatitis A virus genotype IIa. J. Clin. Microbiol. 2010, 48, 3306–3315. [Google Scholar] [CrossRef] [PubMed]
- Costa-Mattioli, M.; Napoli, A.D.; Ferré, V.; Billaudel, S.; Pérez-Bercoff, R.; Cristina, J. Genetic variability of hepatitis A virus. J. Gen. Virol. 2003, 84, 3191–3201. [Google Scholar] [CrossRef] [PubMed]
- Tjon, G.M.S.; Wijkmans, C.J.; Coutinho, R.A.; Koek, A.G.; van den Hoek, J.A.R.; Leenders, A.C.A.P.; Schneeberger, P.M.; Bruisten, S.M. Molecular epidemiology of hepatitis A in Noord-Bbrabant, The Netherlands. J. Clin. Virol. 2005, 32, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Coudray-Meunier, C.; Fraisse, A.; Mokhtari, C.; Martin-Latil, S.; Roque-Afonso, A.M.; Perelle, S. Hepatitis A virus subgenotyping based on RT-qPCR assays. BMC Microbiol. 2014, 14, 296. [Google Scholar] [CrossRef] [PubMed]
- Gillesberg Lassen, S.; Soborg, B.; Midgley, S.E.; Steens, A.; Vold, L.; Stene-Johansen, K.; Rimhanen-Finne, R.; Kontio, M.; Lofdahl, M.; Sundqvist, L.; et al. Ongoing multi-strain food-borne hepatitis A outbreak with frozen berries as suspected vehicle: Four Nordic countries affected, October 2012 to April 2013. Euro Surveill. 2013, 18, 20467. [Google Scholar]
- Miyamura, T.; Ishii, K.; Kanda, T.; Tawada, A.; Sekimoto, T.; Wu, S.; Nakamoto, S.; Arai, M.; Fujiwara, K.; Imazeki, F.; et al. Possible widespread presence of hepatitis A virus subgenotype IIIA in Japan: Recent trend of hepatitis a causing acute liver failure. Hepatol. Res. 2012, 42, 248–253. [Google Scholar]
- Kumbang, J.; Ejide, S.; Tedder, R.S.; Ngui, S.L. Outbreak of hepatitis A in an extended family after importation by non-immune travellers. Epidemiol. Infect. 2012, 140, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Stene-Johansen, K.; Jonassen, T.O.; Skaug, K. Characterization and genetic variability of hepatitis a virus genotype IIIA. J. Gen. Virol. 2005, 86, 2739–2745. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.A.; Walimbe, A.M.; Cherian, S.; Arankalle, V.A. Full length genomes of genotype IIIA hepatitis A virus strains (1995–2008) from India and estimates of the evolutionary rates and ages. Infect. Genet. Evol. 2009, 9, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.; Lee, H.J.; Jang, J.-H.; Kim, J.S.; Lee, S.H.; Kim, J.-W.; Park, S.J.; Park, Y.M.; Hwang, S.G.; Rim, K.S.; et al. Hepatitis A virus genotype and its correlation with the clinical outcome of acute hepatitis A in Korea: 2006–2008. J. Med. Virol. 2011, 83, 2073–2081. [Google Scholar]
- Hussain, Z.; Husain, S.; Almajhdi, F.; Kar, P. Immunological and molecular epidemiological characteristics of acute and fulminant viral hepatitis A. Virol. J. 2011, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Chitambar, S.; Joshi, M.; Lole, K.; Walimbe, A.; Vaidya, S. Cocirculation of and coinfections with hepatitis A virus subgenotypes IIIA and IB in patients from Pune, Western India. Hepatol. Res. 2007, 37, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Wasley, A.; Samandari, T.; Bell, B.P. Incidence of hepatitis A in the United States in the era of vaccination. J. Am. Med. Assoc. 2005, 294, 194–201. [Google Scholar] [CrossRef]
- Chironna, M.; Prato, R.; Sallustio, A.; Martinelli, D.; Tafuri, S.; Quarto, M.; Germinario, C. Hepatitis A in Puglia (South Italy) after 10 years of universal vaccination: Need for strict monitoring and catch-up vaccination. BMC Infect. Dis. 2012, 12, 271. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, A.; Bruguera, M.; Plans, P.; Costa, J.; Salleras, L.L. Prevalence of hepatitis A antibodies in schoolchildren in Catalonia (Spain) after the introduction of universal hepatitis A immunization. J. Med. Virol. 2004, 73, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Bonanni, P.; Boccalini, S.; Bechini, A. Vaccination against hepatitis A in children: A review of the evidence. Ther. Clin. Risk Manag. 2007, 3, 1071–1076. [Google Scholar] [PubMed]
- Robertson, B.H.; Jansen, R.W.; Khanna, B.; Totsuka, A.; Nainan, O.V.; Siegl, G.; Widell, A.; Margolis, H.S.; Isomura, S.; Ito, K.; et al. Genetic relatedness of hepatitis A virus strains recovered from different geographical regions. J. Gen. Virol. 1992, 73, 1365–1377. [Google Scholar]
- Sánchez, G.; Pintó, R.M.; Vanaclocha, H.; Bosch, A. Molecular characterization of hepatitis A virus isolates from a transcontinental shellfish-borne outbreak. J. Clin. Microbiol. 2002, 40, 4148–4155. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, G.; Bosch, A.; Pintó, R.M. Genome variability and capsid structural constraints of hepatitis A virus. J. Virol. 2003, 77, 452–459. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Andrea, L.; Pérez-Rodríguez, F.J.; De Castellarnau, M.; Manzanares, S.; Lite, J.; Guix, S.; Bosch, A.; Pintó, R.M. Hepatitis A Virus Genotype Distribution during a Decade of Universal Vaccination of Preadolescents. Int. J. Mol. Sci. 2015, 16, 6842-6854. https://doi.org/10.3390/ijms16046842
D’Andrea L, Pérez-Rodríguez FJ, De Castellarnau M, Manzanares S, Lite J, Guix S, Bosch A, Pintó RM. Hepatitis A Virus Genotype Distribution during a Decade of Universal Vaccination of Preadolescents. International Journal of Molecular Sciences. 2015; 16(4):6842-6854. https://doi.org/10.3390/ijms16046842
Chicago/Turabian StyleD’Andrea, Lucía, Francisco J. Pérez-Rodríguez, Montserrat De Castellarnau, Sandra Manzanares, Josep Lite, Susana Guix, Albert Bosch, and Rosa M. Pintó. 2015. "Hepatitis A Virus Genotype Distribution during a Decade of Universal Vaccination of Preadolescents" International Journal of Molecular Sciences 16, no. 4: 6842-6854. https://doi.org/10.3390/ijms16046842