
Evodiamine Attenuates PDGF-BB-Induced Migration of Rat Vascular Smooth Muscle Cells through Activating PPARγ


Abstract
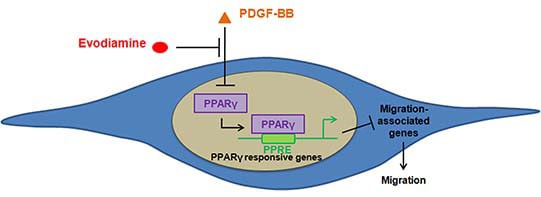
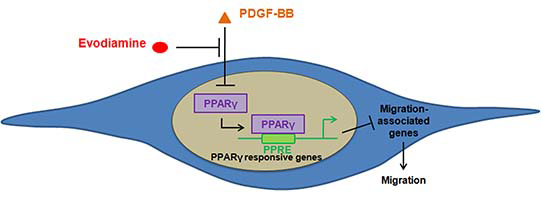
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
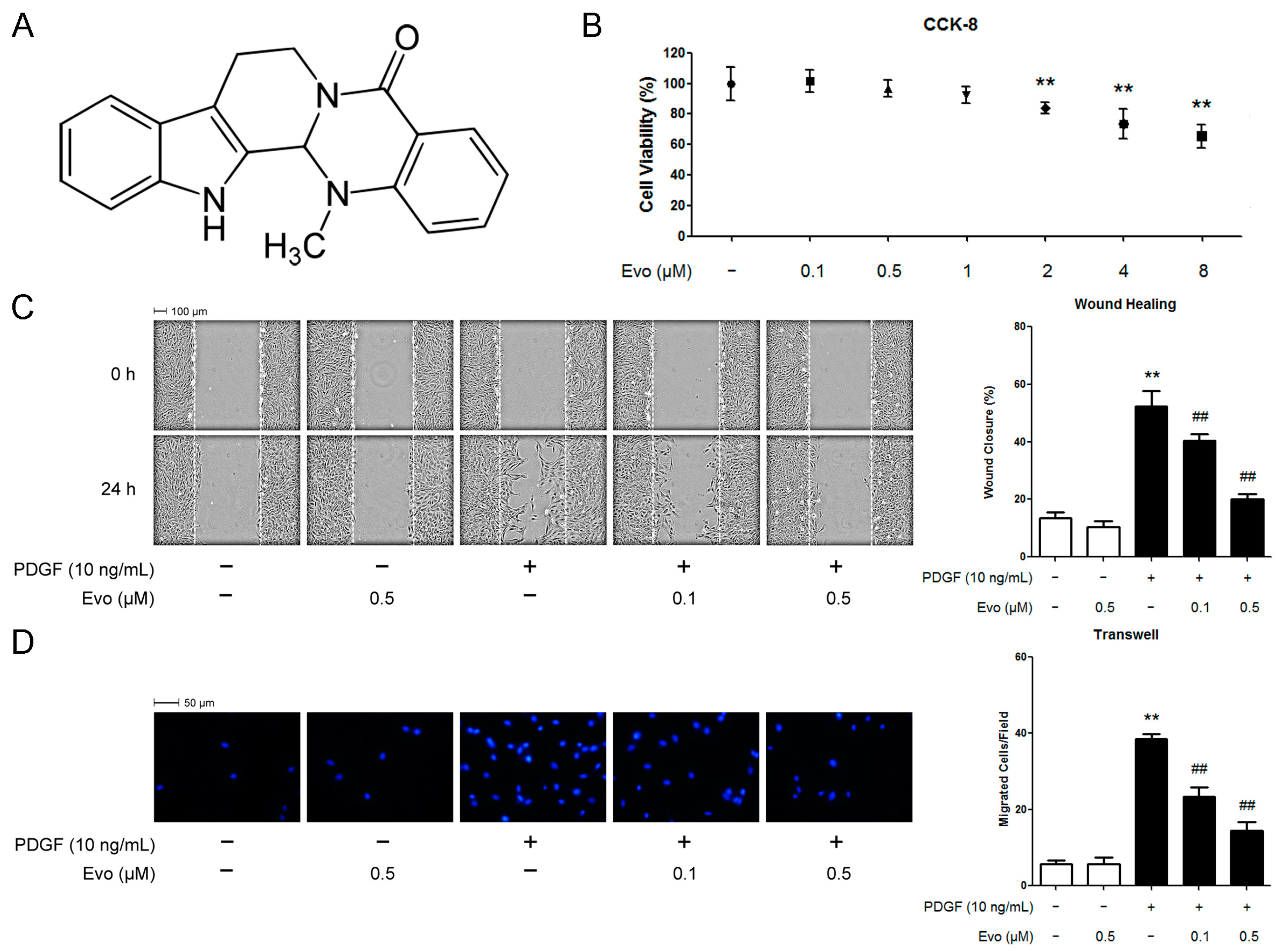
2.1. Evodiamine Inhibits Platelet-Derived Growth Factor (PDGF)-BB-Induced Vascular Smooth Muscle Cell (VSMC) Migration
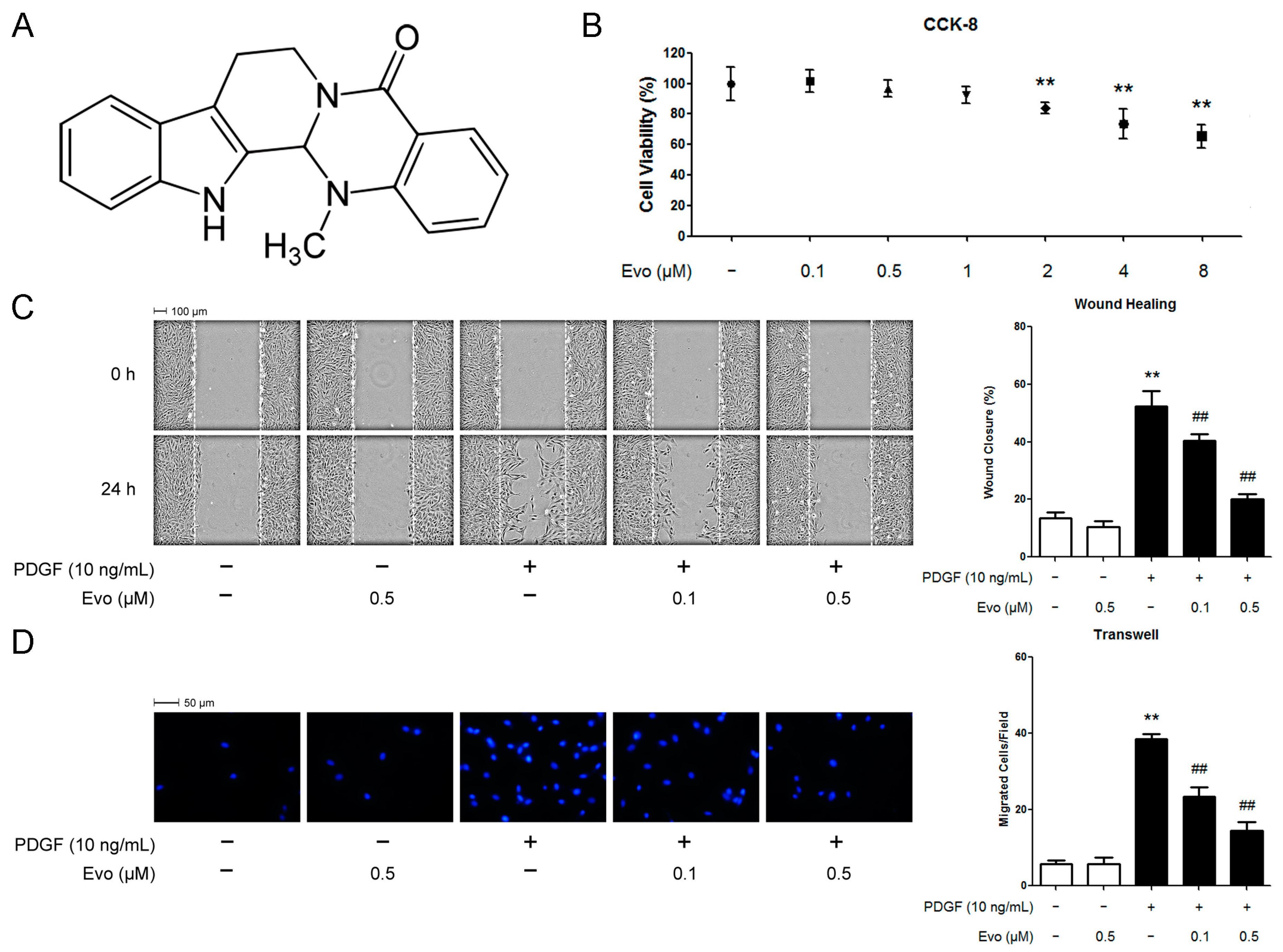
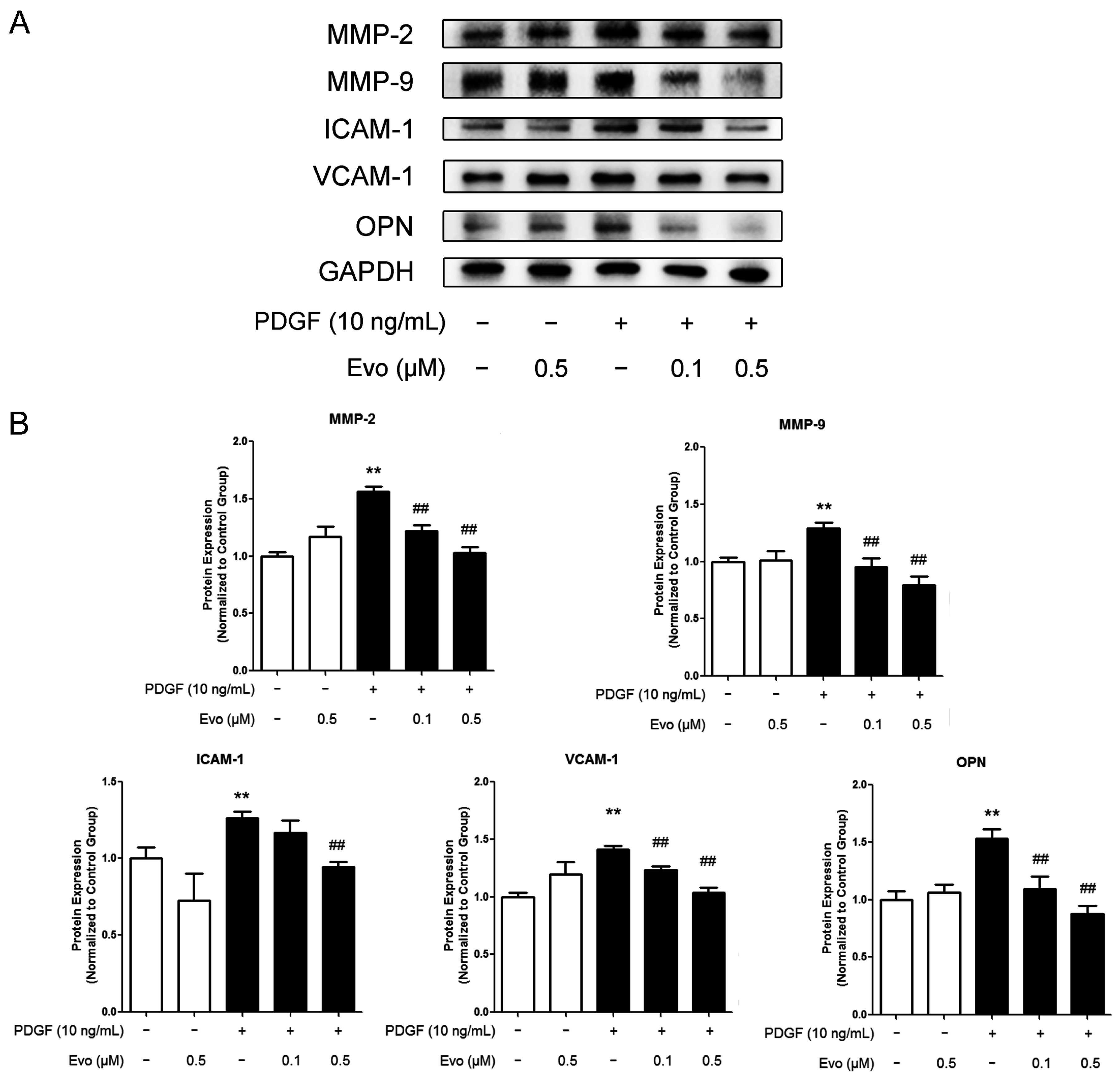
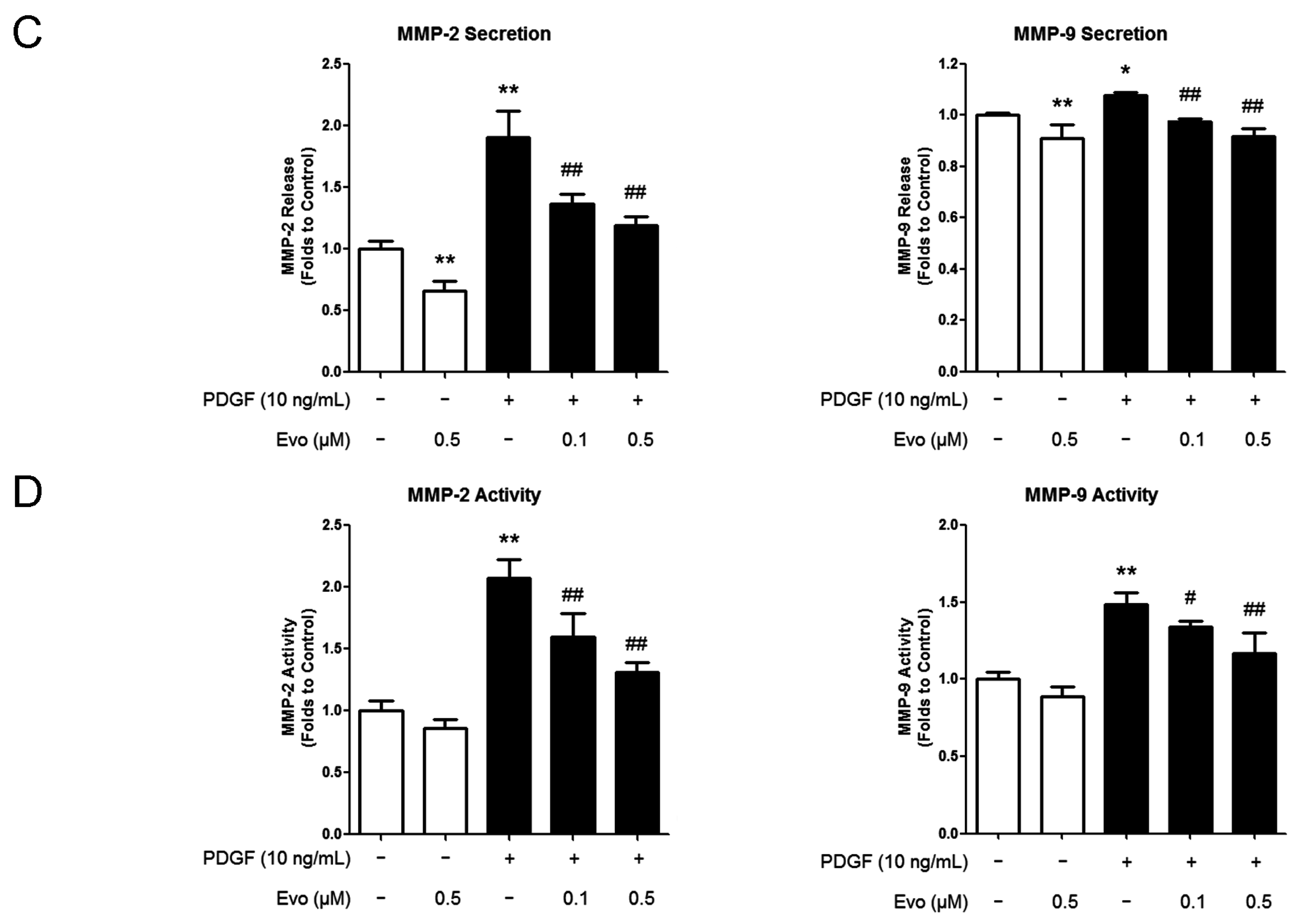
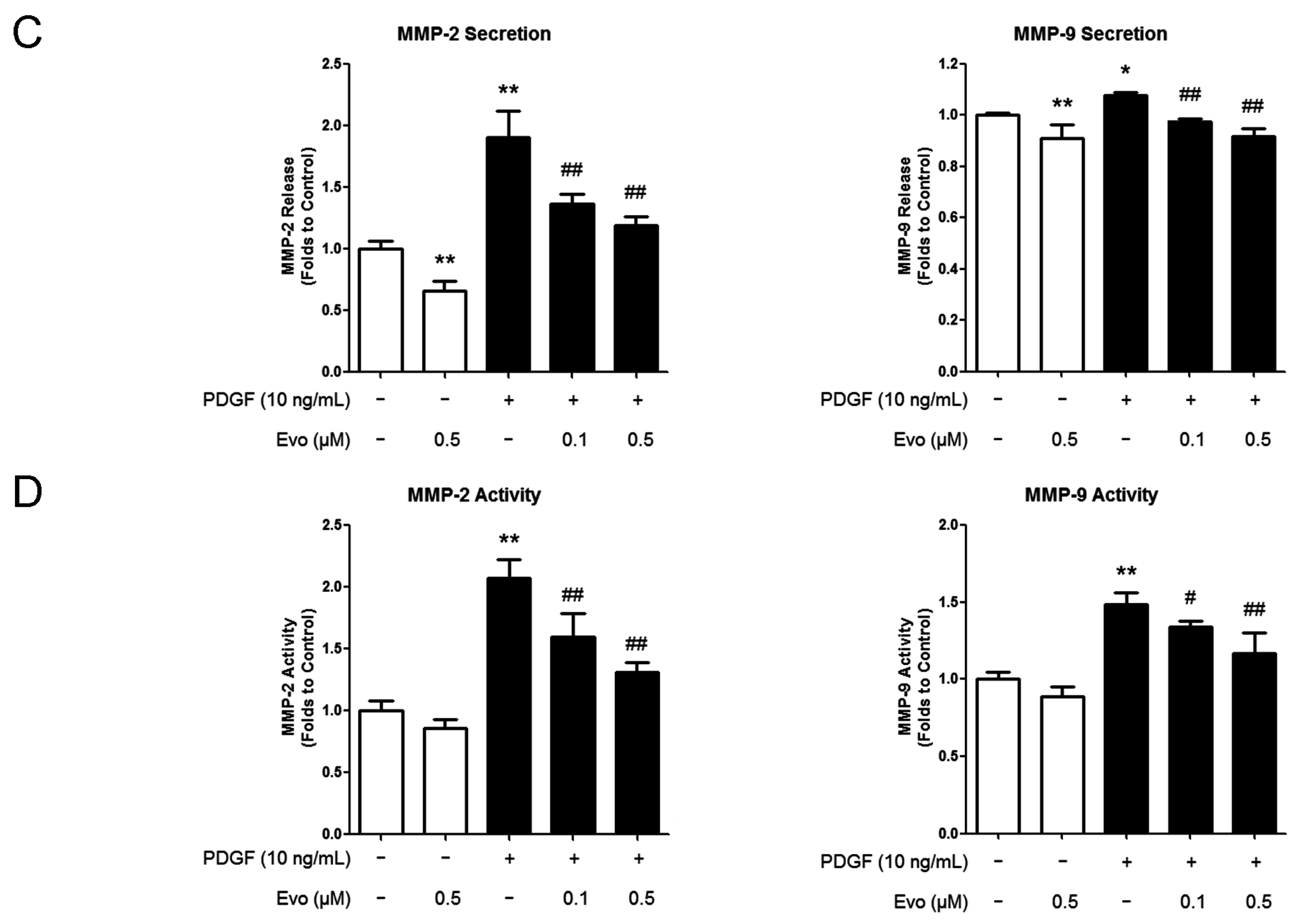
2.2. Evodiamine Inhibits the Expression of Migration-Associated Regulators

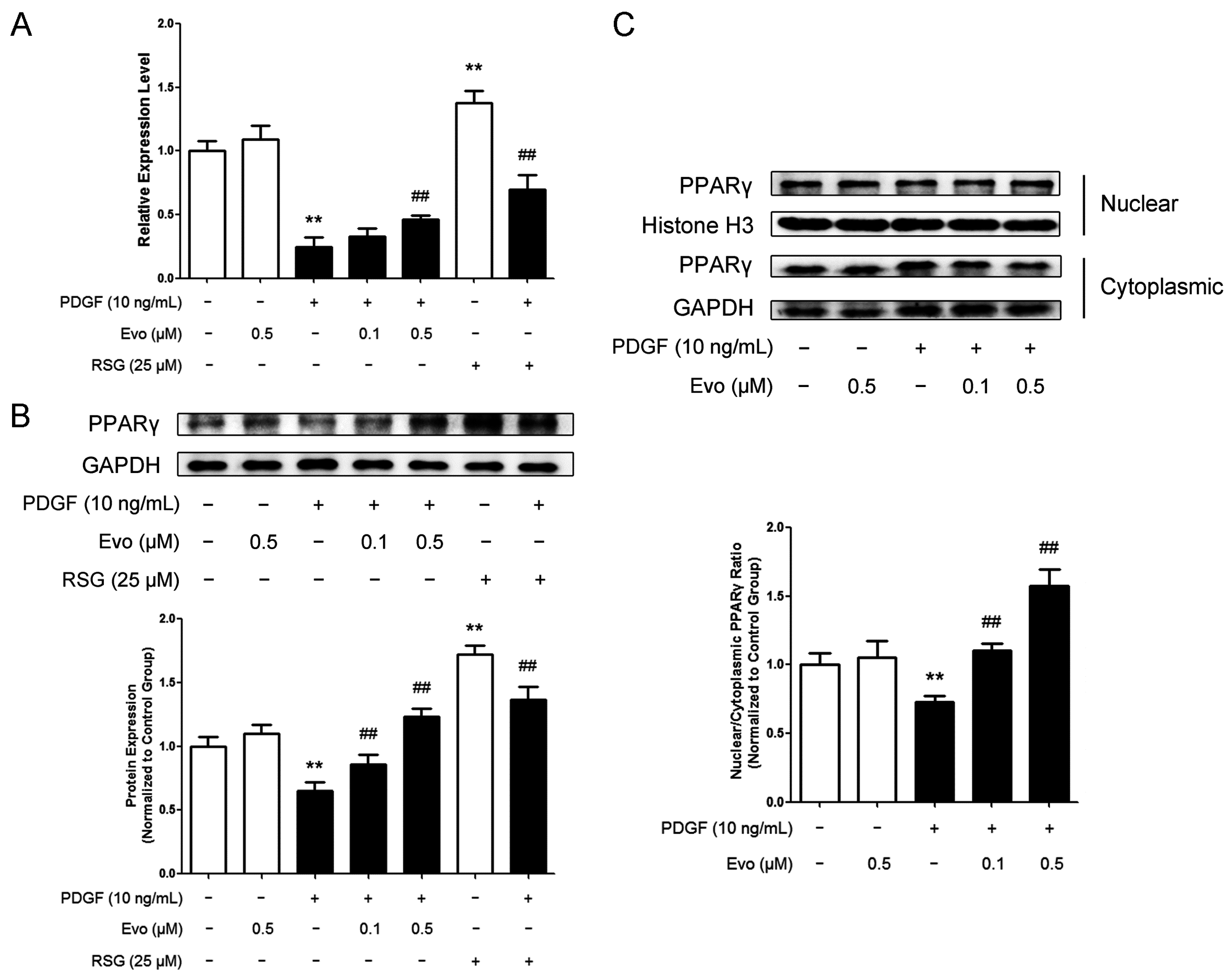
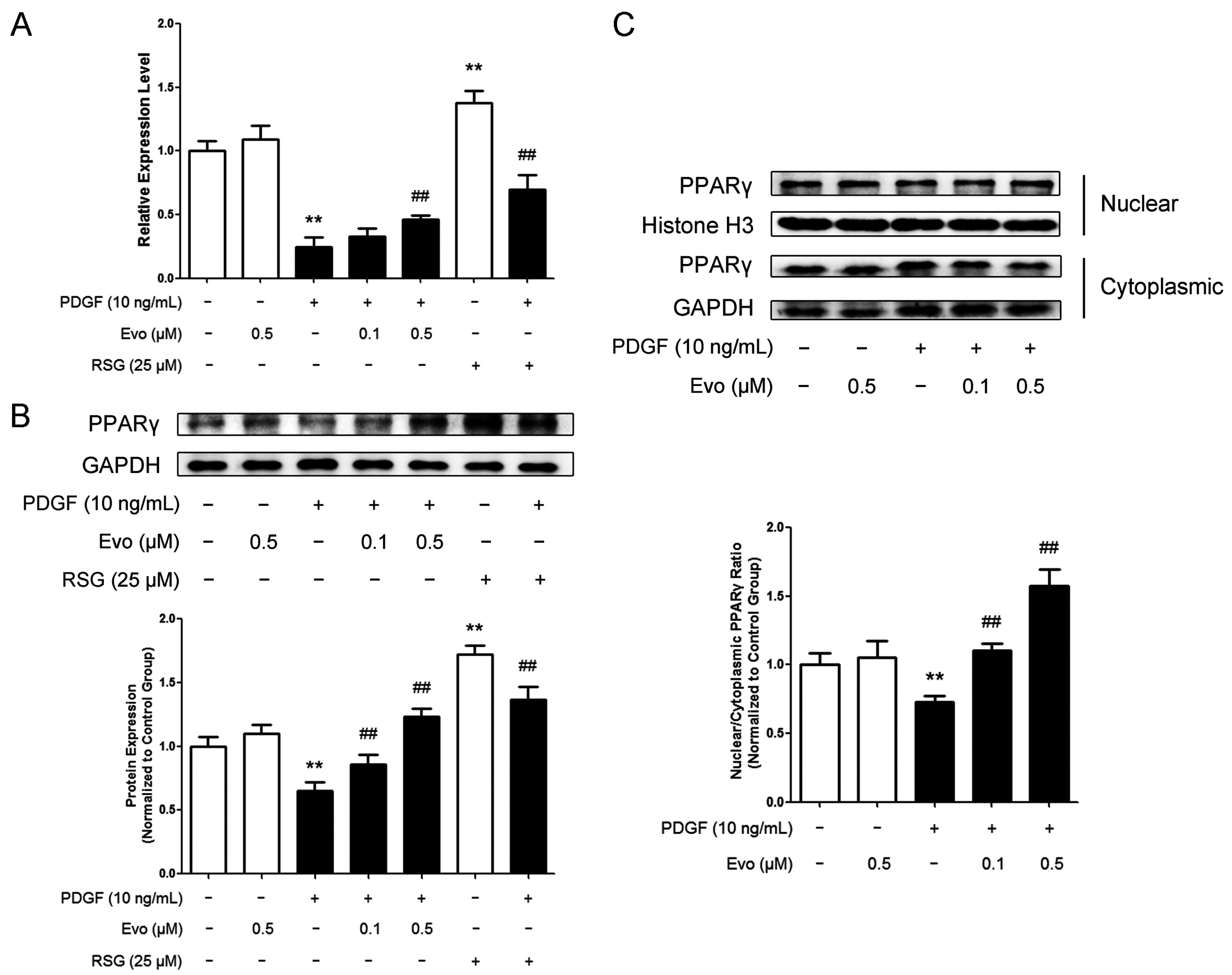
2.3. Evodiamine Increases Peroxisome Proliferator-Activated Receptor γ (PPARγ) Expression and Nuclear Translocation in VSMCs
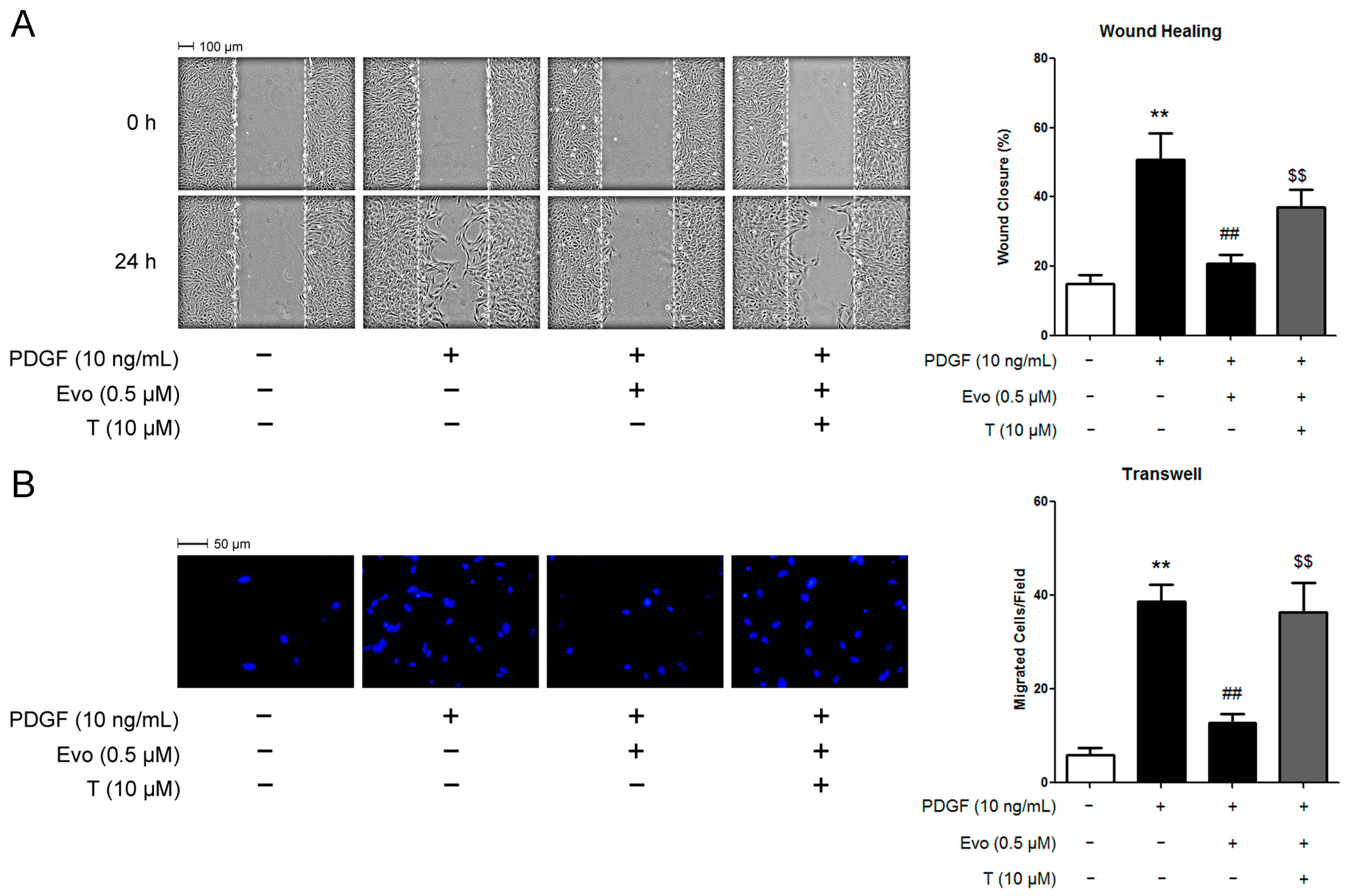
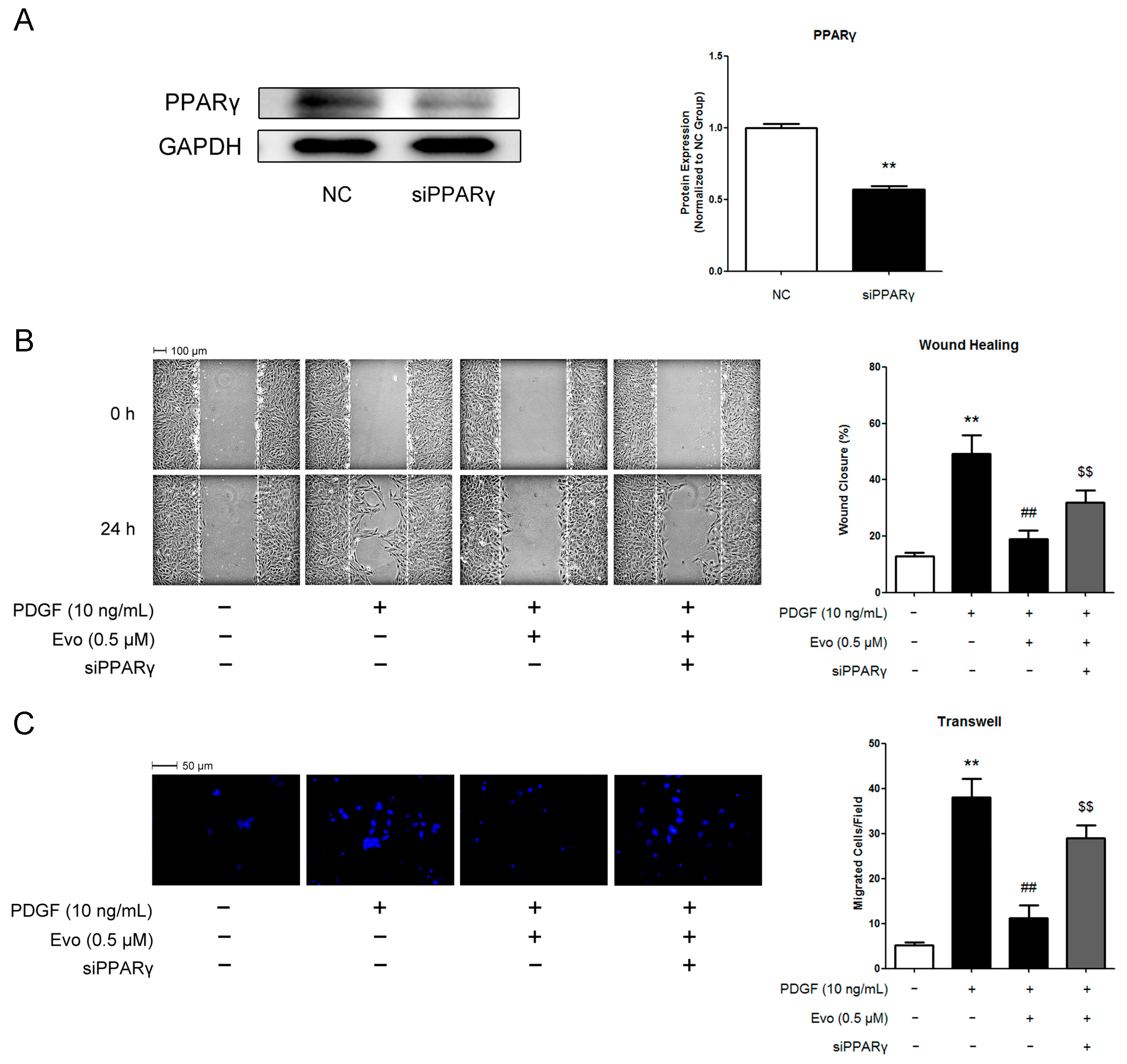
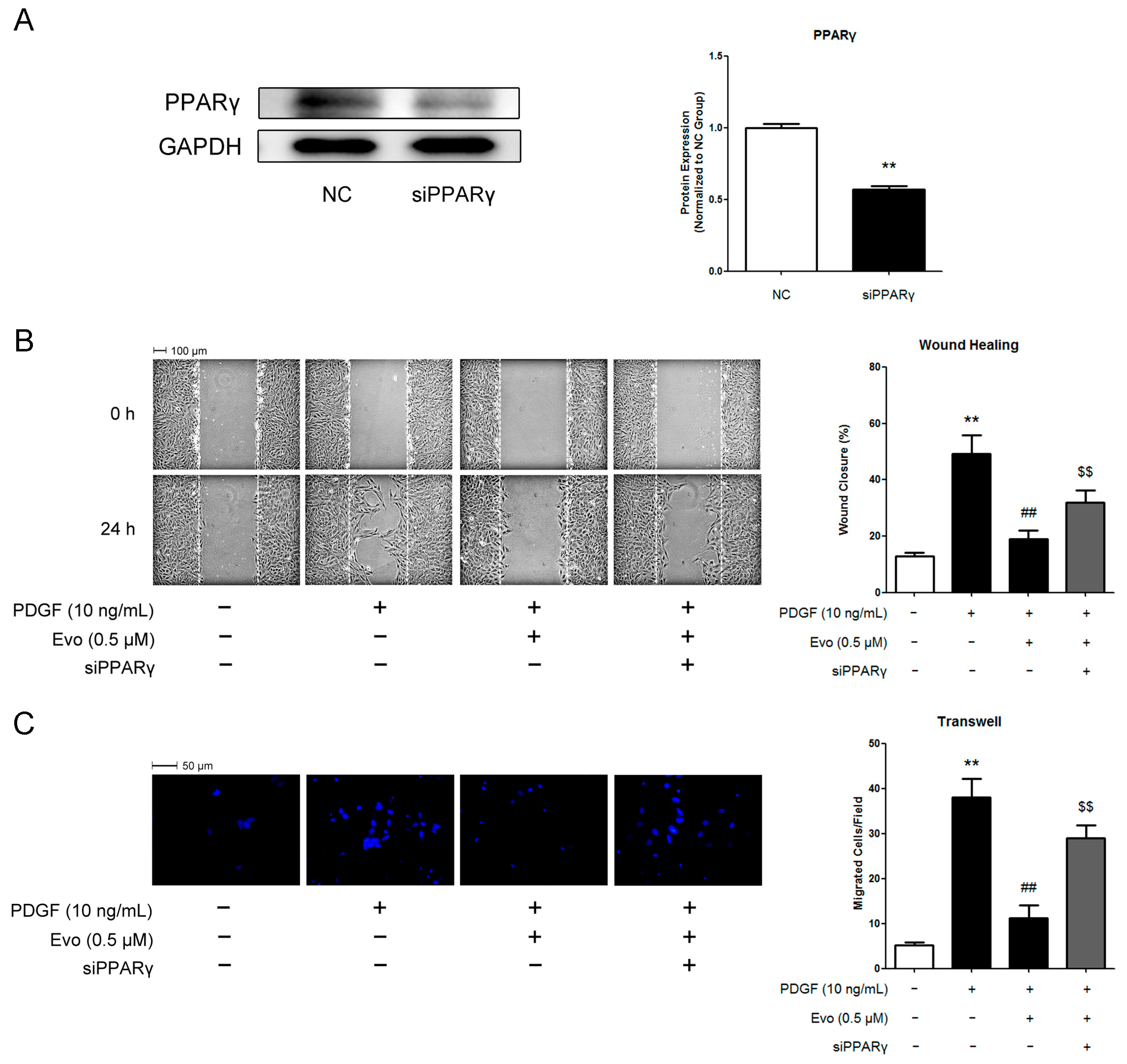
2.4. PPARγ Mediates the Actions of Evodiamine in Regulating VSMC Migration

3. Discussion
4. Experimental Section
4.1. Chemicals and Materials
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Wound Healing Assay
4.5. Boyden Chamber Assay
4.6. Western Blotting Analysis
4.7. Matrix Metalloproteinase (MMP)-2 and MMP-9 Activity Assays
4.8. Enzyme Linked Immunosorbent Assay (ELISA)
4.9. Real-Time RT-PCR
4.10. PPARγ RNAi
4.11. Data Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gerthoffer, W.T. Mechanisms of vascular smooth muscle cell migration. Circ. Res. 2007, 100, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Lacolley, P.; Regnault, V.; Nicoletti, A.; Li, Z.; Michel, J.B. The vascular smooth muscle cell in arterial pathology: A cell that can take on multiple roles. Cardiovasc. Res. 2012, 95, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharmacol. 2009, 78, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Galis, Z.S.; Johnson, C.; Godin, D.; Magid, R.; Shipley, J.M.; Senior, R.M.; Ivan, E. Targeted disruption of the matrix metalloproteinase-9 gene impairs smooth muscle cell migration and geometrical arterial remodeling. Circ. Res. 2002, 91, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Marx, N.; Schonbeck, U.; Lazar, M.A.; Libby, P.; Plutzky, J. Peroxisome proliferator-activated receptor γ activators inhibit gene expression and migration in human vascular smooth muscle cells. Circ. Res. 1998, 83, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Jin, N.; Wang, G.; Zhang, F.; Mao, J.; Wang, X. Peroxisome proliferator-activated receptor γ agonist improves arterial stiffness in patients with type 2 diabetes mellitus and coronary artery disease. Metabolism 2007, 56, 1396–1401. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Xiong, Z.B.; Tang, A.L.; Ma, H.; Ma, Y.D.; Wu, J.G.; Dong, Y.G. Rosiglitazone-induced myocardial protection against ischaemia-reperfusion injury is mediated via a phosphatidylinositol 3-kinase/Akt-dependent pathway. Clin. Exp. Pharmacol. Physiol. 2010, 37, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Wakino, S.; Kintscher, U.; Kim, S.; Yin, F.; Hsueh, W.A.; Law, R.E. Peroxisome proliferator-activated receptor γ ligands inhibit retinoblastoma phosphorylation and G1→S transition in vascular smooth muscle cells. J. Biol. Chem. 2000, 275, 22435–22441. [Google Scholar] [CrossRef] [PubMed]
- Joner, M.; Finn, A.V.; Farb, A.; Mont, E.K.; Kolodgie, F.D.; Ladich, E.; Kutys, R.; Skorija, K.; Gold, H.K.; Virmani, R. Pathology of drug-eluting stents in humans: Delayed healing and late thrombotic risk. J. Am. Coll. Cardiol. 2006, 48, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Hu, C. Evodiamine: A novel anti-cancer alkaloid from Evodia Rutaecarpa. Molecules 2009, 14, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, M.; Suzuki, H. Inhibition by evodiamine of hepatocyte growth factor-induced invasion and migration of tumor cells. Biol. Pharm. Bull. 2004, 27, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wang, X.F.; Zhou, Q.M.; Zhang, T.L.; Lu, Y.Y.; Zhang, H.; Su, S.B. Evodiamine induces apoptosis and inhibits metastasis in MDA-MB-231 human breast cancer cells in vitro and in vivo. Oncol. Rep. 2013, 30, 685–694. [Google Scholar] [PubMed]
- Chiou, W.F.; Chou, C.J.; Shum, A.Y.; Chen, C.F. The vasorelaxant effect of evodiamine in rat isolated mesenteric arteries: Mode of action. Eur. J. Pharmacol. 1992, 215, 277–283. [Google Scholar] [CrossRef]
- Hung, P.H.; Lin, L.C.; Wang, G.J.; Chen, C.F.; Wang, P.S. Inhibitory effect of evodiamine on aldosterone release by zona glomerulosa cells in male rats. Chin. J. Physiol. 2001, 44, 53–57. [Google Scholar] [PubMed]
- Heo, S.K.; Yun, H.J.; Yi, H.S.; Noh, E.K.; Park, S.D. Evodiamine and rutaecarpine inhibit migration by light via suppression of NADPH oxidase activation. J. Cell. Biochem. 2009, 107, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Matrix metalloproteinases regulate migration, proliferation, and death of vascular smooth muscle cells by degrading matrix and non-matrix substrates. Cardiovasc. Res. 2006, 69, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Ballantyne, C.M.; Sharrett, A.R.; Smith, L.C.; Davis, C.E.; Gotto, A.M., Jr.; Boerwinkle, E. Circulating adhesion molecules VCAM-1, ICAM-1, and E-selectin in carotid atherosclerosis and incident coronary heart disease cases: The atherosclerosis risk in communities (ARIC) study. Circulation 1997, 96, 4219–4225. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Han, M.; Wen, J.K.; Li, A.Y. Osteopontin stimulates vascular smooth muscle cell migration by inducing FAK phosphorylation and ILK dephosphorylation. Biochem. Biophys. Res. Commun. 2007, 356, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Rebhun, J.F.; Roloff, S.J.; Velliquette, R.A.; Missler, S.R. Identification of evodiamine as the bioactive compound in evodia (Evodia Rutaecarpa Benth.) fruit extract that activates human peroxisome proliferator-activated receptor γ (PPARγ). Fitoterapia 2014, 101, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Li, Z.; Yuan, F. Evodiamine might inhibit TGF-β1-induced epithelial-mesenchymal transition in NRK52E cells via smad and PPAR-γ pathway. Cell Biol. Int. 2014, 38, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, W.A.; Jackson, S.; Law, R.E. Control of vascular cell proliferation and migration by PPAR-γ: A new approach to the macrovascular complications of diabetes. Diabetes Care 2001, 24, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Ching, L.C.; Zhao, J.F.; Shyue, S.K.; Lee, H.F.; Kou, Y.R.; Lee, T.S. Essential role of transient receptor potential vanilloid type 1 in evodiamine-mediated protection against atherosclerosis. Acta Physiol. 2013, 207, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Metalloproteinases promote plaque rupture and myocardial infarction: A persuasive concept waiting for clinical translation. Matrix Biol. 2015, 44–46, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Kawamura, T.; Masutani, K.; Peng, X.; Stolz, D.B.; Pribis, J.P.; Billiar, T.R.; Sun, X.; Bermudez, C.A.; Toyoda, Y.; et al. Oral intake of hydrogen-rich water inhibits intimal hyperplasia in arterialized vein grafts in rats. Cardiovasc. Res. 2012, 94, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Tumor angiogenesis: Mmp-mediated induction of intravasation- and metastasis-sustaining neovasculature. Matrix Biol. 2015, 44–46, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.L.; Yu, C.C.; Chen, C.S.; Guh, J.H. Tubulin-binding agents down-regulate matrix metalloproteinase-2 and -9 in human hormone-refractory prostate cancer cells–A critical role of CDK1 in mitotic entry. Biochem. Pharmacol. 2015, 94, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vascul. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.L.; Wu, X.J.; Liu, Y.; Xie, J.Q. Berberine counteracts enhanced il-8 expression of AGS cells induced by evodiamine. Life Sci. 2013, 93, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.; Chang, L.; Fan, Y.; Zhang, J.; Chen, Y.E. PPARs and the cardiovascular system. Antioxid. Redox Signal. 2009, 11, 1415–1452. [Google Scholar] [CrossRef] [PubMed]
- Goetze, S.; Kintscher, U.; Kim, S.; Meehan, W.P.; Kaneshiro, K.; Collins, A.R.; Fleck, E.; Hsueh, W.A.; Law, R.E. Peroxisome proliferator-activated receptor-γ ligands inhibit nuclear but not cytosolic extracellular signal-regulated kinase/mitogen-activated protein kinase-regulated steps in vascular smooth muscle cell migration. J. Cardiovasc. Pharmacol. 2001, 38, 909–921. [Google Scholar] [CrossRef] [PubMed]
- Salomone, S.; Drago, F. Effects of PPARγ ligands on vascular tone. Curr. Mol. Pharmacol. 2012, 5, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Zhang, S.; Driss, A.; Liu, Z.R.; Kim, H.R.; Wang, Y.; Ritenour, C.; Zhau, H.E.; Kucuk, O.; Chung, L.W.; et al. PDGF upregulates Mcl-1 through activation of β-catenin and HIF-1α-dependent signaling in human prostate cancer cells. PLoS ONE 2012, 7, e30764. [Google Scholar] [CrossRef] [PubMed]
- Farmer, S.R. Regulation of PPARγ activity during adipogenesis. Int. J. Obes. 2005, 29 (Suppl. 1), S13–S16. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Feng, S.; Wei, L.; Wang, Z.; Hong, D.; Wang, Q. Evodiamine, a novel inhibitor of the wnt pathway, inhibits the self-renewal of gastric cancer stem cells. Int. J. Mol. Med. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.; Mohai, L.G.; Schwartz, S.M. Induction of polyploidy in cultures of neonatal rat aortic smooth muscle cells. Circ. Res. 1986, 59, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Ding, Y.; Tao, W.; Zhang, W.; Liang, T.; Liu, C. Naringenin inhibits TNF-α induced VSMC proliferation and migration via induction of HO-1. Food Chem. Toxicol. 2012, 50, 3025–3031. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, X.; Chen, S.; Liu, M.; Liang, T.; Liu, C. Evodiamine Attenuates PDGF-BB-Induced Migration of Rat Vascular Smooth Muscle Cells through Activating PPARγ. Int. J. Mol. Sci. 2015, 16, 28180-28193. https://doi.org/10.3390/ijms161226093
Ge X, Chen S, Liu M, Liang T, Liu C. Evodiamine Attenuates PDGF-BB-Induced Migration of Rat Vascular Smooth Muscle Cells through Activating PPARγ. International Journal of Molecular Sciences. 2015; 16(12):28180-28193. https://doi.org/10.3390/ijms161226093
Chicago/Turabian StyleGe, Xie, Siyu Chen, Mei Liu, Tingming Liang, and Chang Liu. 2015. "Evodiamine Attenuates PDGF-BB-Induced Migration of Rat Vascular Smooth Muscle Cells through Activating PPARγ" International Journal of Molecular Sciences 16, no. 12: 28180-28193. https://doi.org/10.3390/ijms161226093