1. Introduction
Sulfur mustard (SM) or bis(2-chloroethyl) sulfide is a powerful bifunctional alkylating and vesicating chemical warfare agent used in numerous military conflicts during the 20th century [
1,
2]. SM is known as a blistering agent, first introduced as a weapon during World War I. Exposure to SM causes the depletion of cellular antioxidant thiols, mainly glutathione (GSH), which may lead to accumulation of reactive oxygen species (ROS) and reactive nitrogen species within cells, inactivation of sulfhydryl-containing enzymes, loss of calcium homeostasis, lipid peroxidation, cellular membrane breakdown and, ultimately, cell death [
3,
4,
5,
6,
7]. These events lead to oxidative stress and macromolecular damage, including DNA, RNA and protein damage, which trigger intricate signaling pathways and modulate gene expression, causing a series of SM-associated toxic responses [
5,
6,
8,
9]. In addition, SM or CEES (2-chloroethyl ethyl sulfide, SM analog) caused oxidative stress results in the 8-oxo-2-deoxyguanosine DNA adduct, as well as lipid and protein oxidation, which can cause inflammation and other toxic responses in skin [
10,
11].
SM is hydrophobic in nature. Therefore, it easily penetrates and accumulates in the lipid component of exposed tissues. In case of contact with skin, SM not only accumulates in the skin, but also distributes to other tissues, such as brain, kidney, muscle, spleen, liver and bone marrow, through the systemic circulation [
12]. SM-induced injuries can take several months to heal and require long-term medical treatment. Exposure to SM triggers an array of complex signal transduction pathways, suggesting the need for pleiotropic agents or combination therapies to treat SM-induced injuries. This is consistent with treating diseases with multi-components in traditional Chinese medicine. The exogenous addition of antioxidants, such as GSH,
N-acetyl cysteine (NAC), vitamin E, superoxide dismutase (SOD), catalase, sulforaphane and quercetin, was reported to attenuate lung and skin injury by SM/CEES [
5,
11,
13,
14,
15]; however, most of them exhibit a stronger protective effect than therapeutic potential [
5,
7,
16]. Due to the potential for use of SM as a chemical terrorism agent, there is renewed interest in developing effective chemopreventive and therapeutic agents.
In this study, we aimed to investigate the therapeutic effects of the mixture of SA and TFA (MSTF) on SM toxicity and to elucidate the potential molecular mechanisms of MSTF-mediated therapeutic efficacy in Sprague Dawley rats.
3. Discussion
Although oxidative stress and inflammation in vesicant-related injury are reported as major consequences of vesicant exposure, antioxidants, scavengers and anti-inflammatory drugs alone do not exhibit potent therapeutic effects [
16,
18]. This suggests that agents are needed with diverse activity towards multiple pathways, including targeting oxidative stress and inflammation. MSTF is the mixture of salvianolic acids (SA) of S
alvia miltiorrhiza and total flavonoids (TFA) of
Anemarrhena asphodeloides. SA is the most abundant water-soluble phenolic compound extracted from Radix S
alvia miltiorrhiza (Danshen). Due to the polyphenolic structure, SA was thought to be comprised of free radical scavengers. Indeed, SA was found to have potent anti-oxidative capabilities
in vitro and
in vivo [
19]. SA possessed a variety of biological and pharmaceutical activities, such as protecting effects against impaired vascular responsiveness in streptozotocin-induced diabetic rats, the cardioprotective effects on animal models of heart hypoxia/reoxygenation injury, the antiplatelet and antithrombotic activities in an arterio-venous shunt model and protective effects against carbon tetrachloride or concanavalin A-induced acute liver damage [
20,
21,
22,
23,
24]. Total flavonoids (TFA) were extracted from the rhizome of
Anemarrhena asphodeloides (Zhimu) used as an antipyretic, anti-inflammatory, anti-diabetic and antidepressant in traditional Chinese medicine [
25]. TFA is composed of 16 flavonoid compounds, including xanthones, mangiferin, ismangiferin, neomangiferin and 1,4,5,6,-tetrahydroxyxanthone. TFA showed valuable bioactivities, such as anti-oxidation, anti-virus, anti-inflammation, anti-skin aging and damage, as well as other activities [
26,
27]. Findings in the present study demonstrated the potent therapeutic efficacy of MSTF in attenuating SM toxic responses. The combination of different types of agents above in MSTF can benefit from each other with different roles in the formula and ultimately gain the goal of enhancing efficacy, which is consistent with the core thinking of traditional Chinese medicine theory. To our knowledge, this is the first time that such a combination has been tested in therapeutic efficacy on SM toxicity.
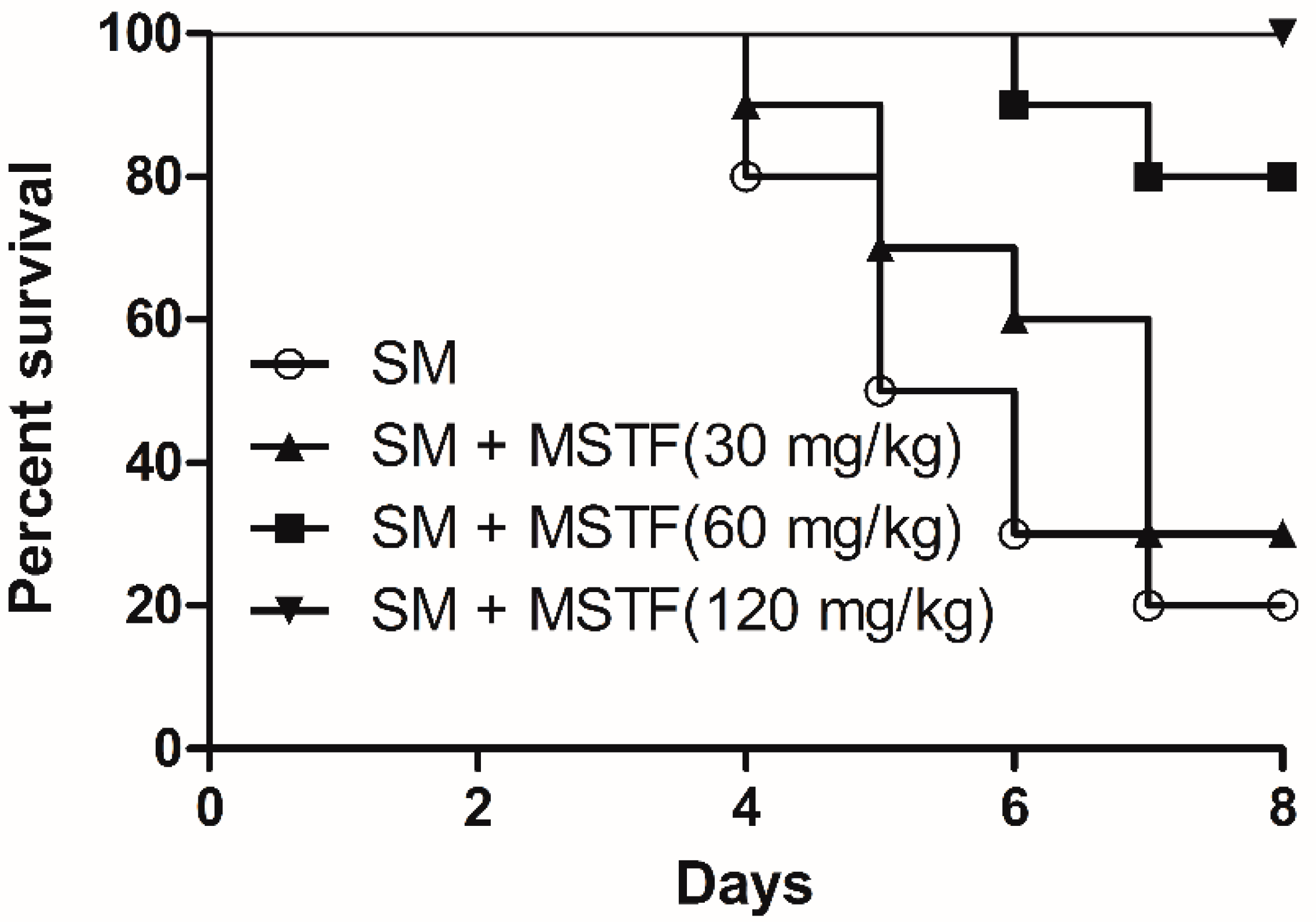
The threat of warfare mustard vesicants’ exploitation also as potential terrorist agents poses challenges to vesicant research to develop effective medical countermeasures to vesicant injuries [
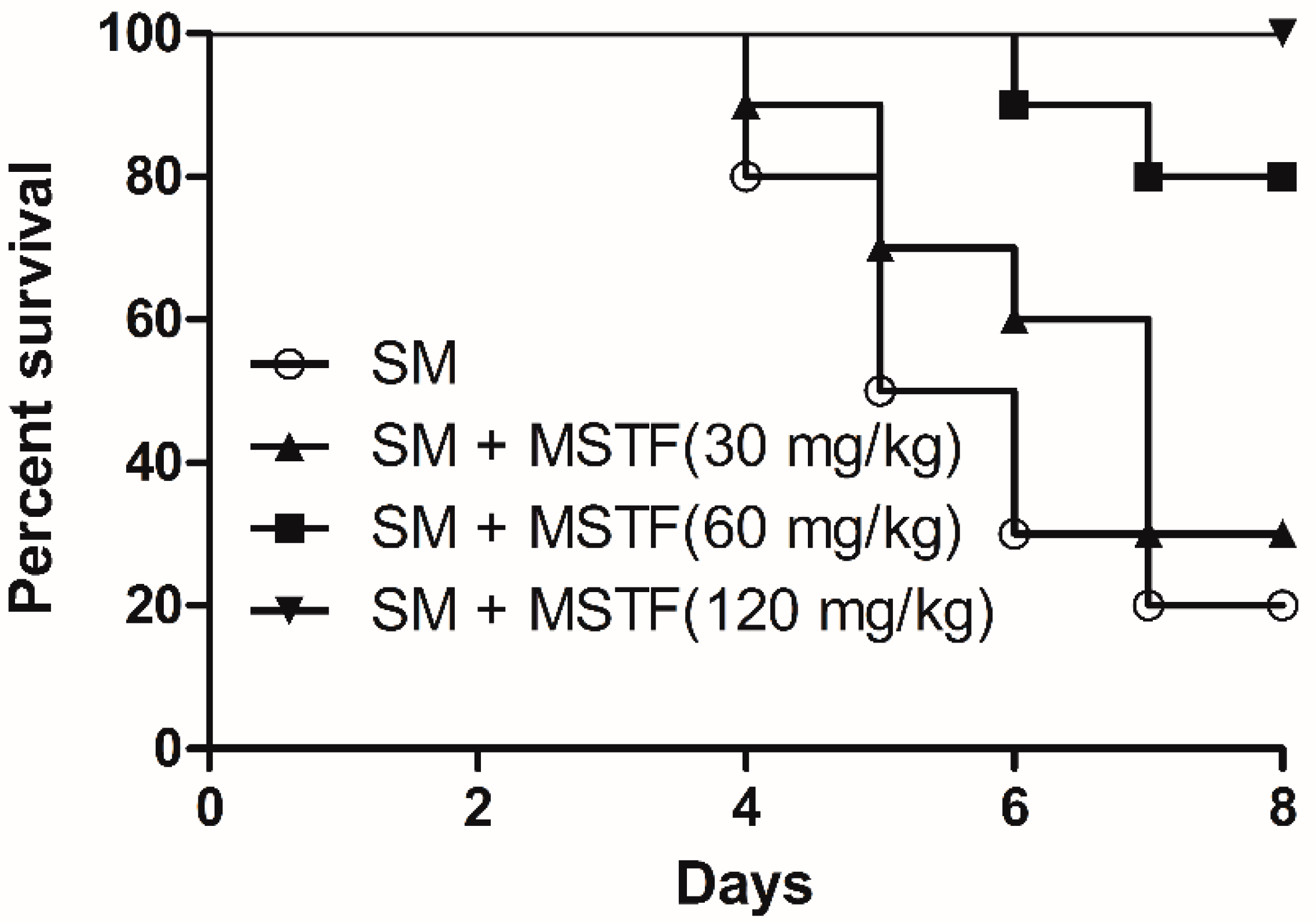
18]. SM-induced injuries can take several months to heal and require long-term medical treatment. Exposure to SM triggers an array of complex signal transduction pathways, suggesting the need for pleiotropic agents or combination therapies to treat SM-induced injuries. The present study revealed that the treatment with MSTF significantly increased the survival levels of rats (
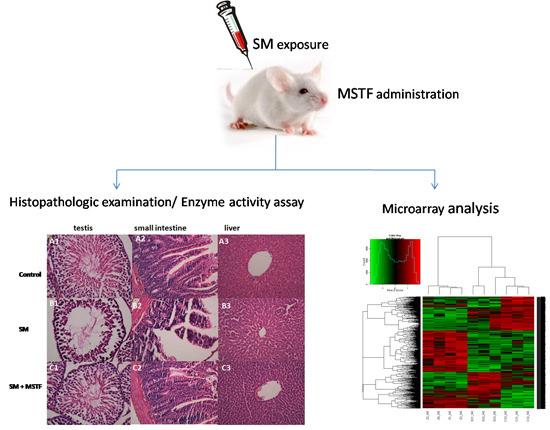
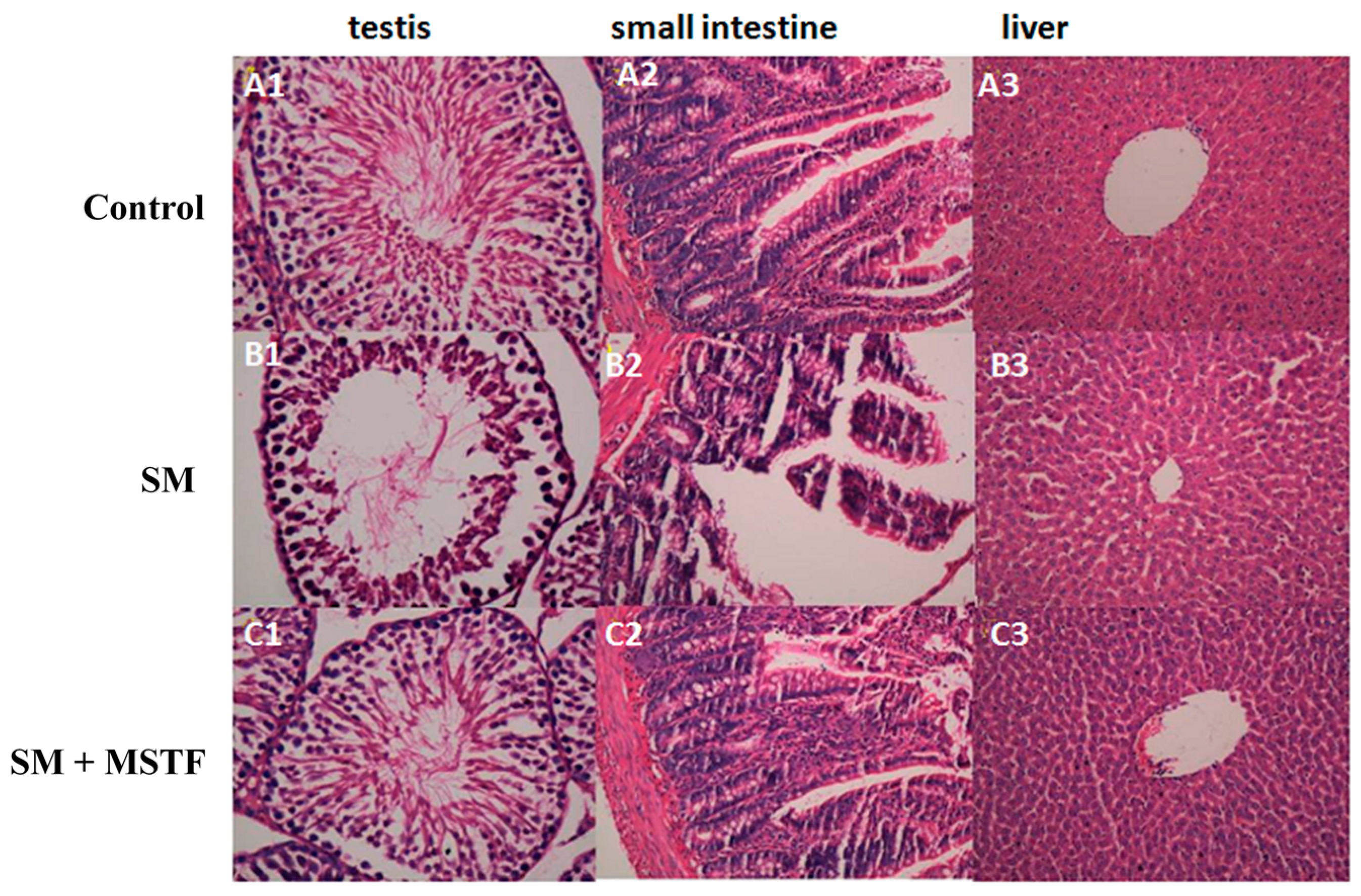
Figure 1) and attenuated the SM-induced morphological changes in the testis, small intestine and liver tissues (
Figure 2), confirming the potent therapeutic efficacy of MSTF in attenuating SM-induced toxic responses.
Exposure to SM or CEES (2-chloroethyl ethyl sulfide, SM analog) has been shown to deplete glutathione (GSH) stores
in vitro and
in vivo [
13,
28,
29]. GSH depletion may inhibit further clearance of SM and, importantly, elicit oxidative stress, lipid peroxidation and macromolecular damage [
10,
28,
30]. In line with these ideas, supplementation with GSH or GSH analogs protected against CEES- or SM-induced cytotoxicity in a number of cell lines [
15], whereas blocking GSH synthesis rendered cells more sensitive to mustard toxicity [
13]. Treatment with GSH was shown to increase the viability of keratinocytes exposed to CEES [
7]. Consistent with these, in this study, MSTF treatment 1 h after SM subcutaneous injection significantly elevated the levels of GSH compared to the SM-only group, which strongly suggests that MSTF improves the animal survival level and inhibits the toxic effects of SM by elevating the levels of GSH. A previous study reported the dose- and time-dependent effects of SM on the antioxidant system and lipid peroxidation in liver and brain of rats [
31]. At SM doses lower than 10 mg/kg after two and seven days of exposure, no significant change in GSH and malondialdehyde (MDA) levels were observed [
31]. In this study, at a single dose of 3.5 mg/kg SM after seven days of exposure, no significant changes in superoxide dismutase (SOD) and MDA levels were observed.
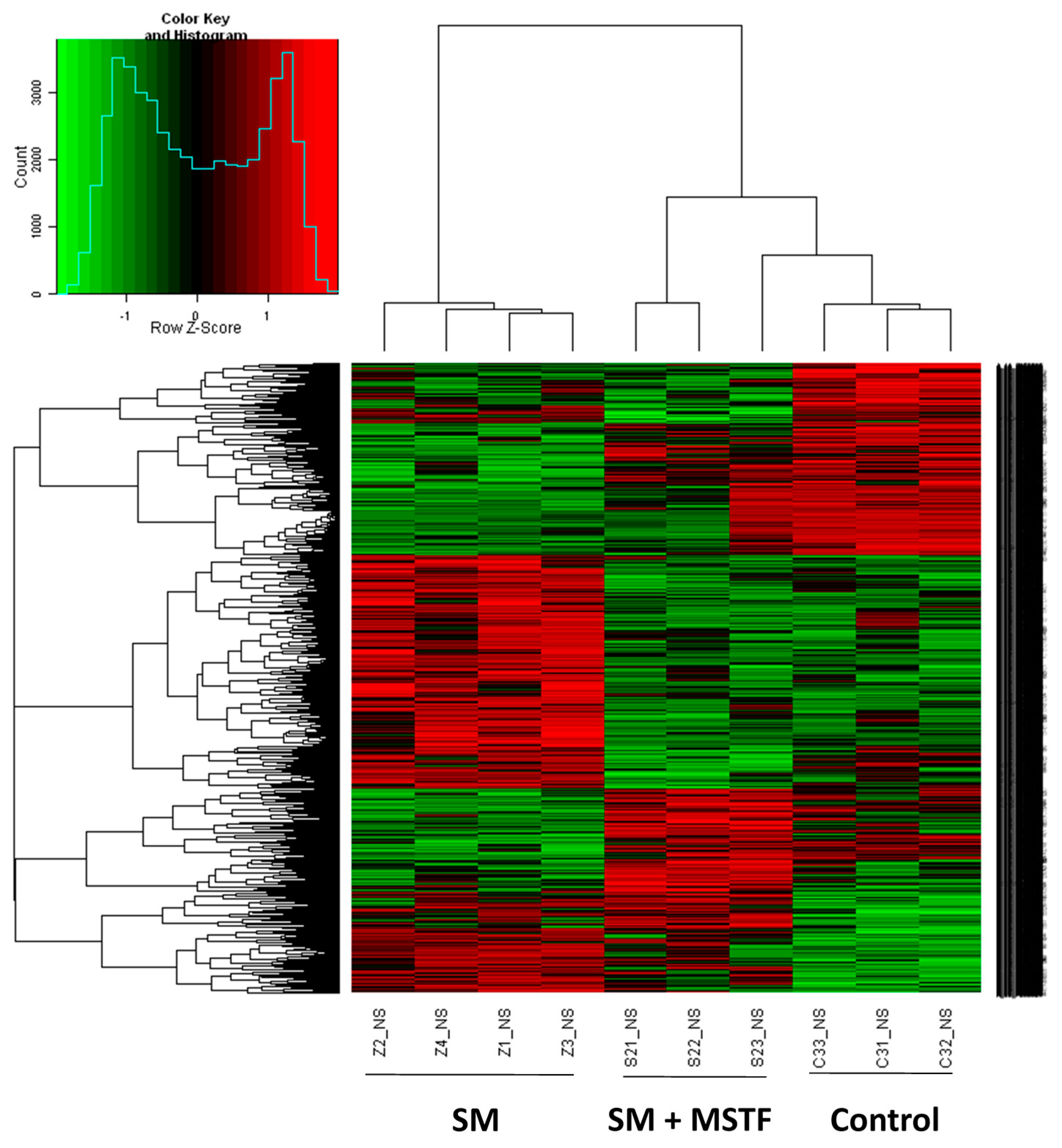
The liver was considered to be a major target organ of SM-induced toxicity either after a single dose or repeated doses [
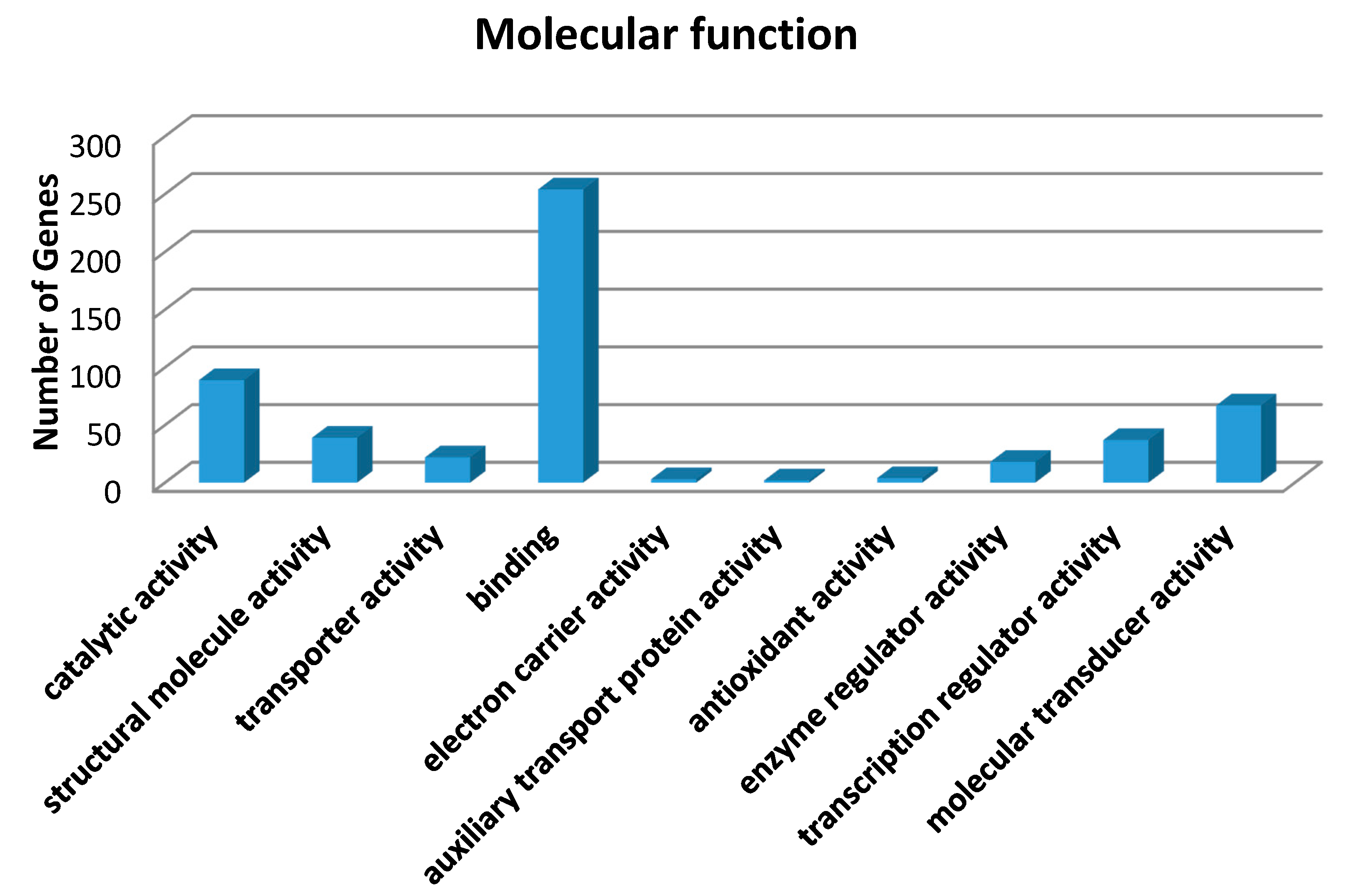
17]. To gain insight into the molecular mechanism of the therapeutic effects of MSTF on SM toxicity, gene expression analysis was conducted on rats liver tissues using microarrays. In the present study, gene expression profiles revealed a dramatic effect of MSTF on alterations of gene expression caused by SM. Interestingly, MSTF treatment prevented SM-induced differential expression of 93.8% (973 genes) of genes. We focused on these genes regulated by SM, and these alterations of expression were abolished or attenuated by MSTF treatment, as these genes might be regulated by MSTF and involved in reversing the toxic effects of SM exposure. Gene ontology mapping of these gene sets revealed several major biological processes, which were cellular process, biological regulation, metabolic processes and response to stimulus (
Figure 4). Pathway enrichment analysis indicated that these genes were mainly involved in a total of 36 pathways (
Figure 5). Many of these pathways were reported to be induced by SM in the previous studies, such as the MAPK signaling pathway [
32,
33,
34,
35,
36], pathways in cancer [
32], antigen processing and presentation [
37], cell adhesion molecules (CAMs) [
36,
37], cell cycle [
32], the p53 signaling pathway [
32,
36], vascular smooth muscle contraction [
32], progesterone-mediated oocyte maturation [
32], hematopoietic cell lineage [
32,
36], neuroactive ligand-receptor interaction [
36], purine metabolism [
32,
36], chemokine signaling pathway [
32], tight junction [
32], bladder cancer [
32] and amebiasis [
32]. Oxidative stress and inflammation in vesicant-related injury were reported as major consequences of SM exposure [
16,
18]. MAPK signaling pathways play critical roles in responding to oxidative stress and inflammation [
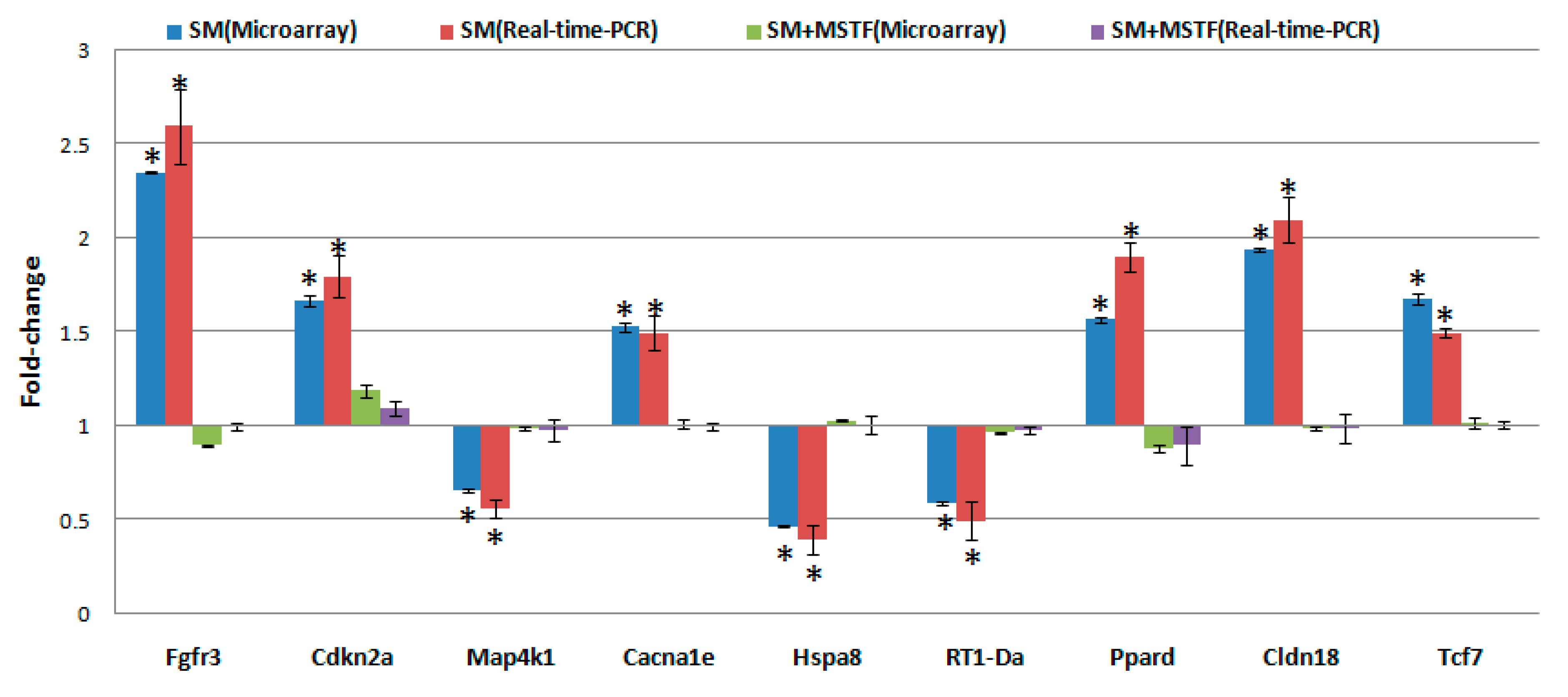
38]. Interestingly, eight of nine SM-induced genes in antigen processing and presentation were significantly downregulated, and these downregulations were abolished or attenuated by treatment with MSTF (
Figure 6). In addition, the previous study reported that silibinin (C
25H
22O
10), a flavanone isolated from the seeds of
Silybum marianum, attenuated sulfur mustard analog (CEES)-induced skin injury by targeting multiple pathways [
16]. These data suggest that MSTF attenuates SM-induced injury by targeting multiple pathways, including MAPK signaling pathways, as well as antigen processing and presentation. These studies further give insight into the antioxidant and potential pleiotropic mechanisms of MSTF in reversing the toxic effects of SM. MSTF should be further assessed as a potential therapeutic against SM-induced injuries.
4. Materials and Methods
4.1. Chemicals
SM was prepared by the Chemical Biological Defence Section (Second Military Medical University, Shanghai, China) and was found to be above 99% pure by gas chromatographic analysis. Salvianolic acids (SA) were isolated from Radix Salvia miltiorrhiza (Danshen) and obtained from Shanghai Winherb Medical Technology Co. (Shanghai, China). Total flavonoids (TFA) of Anemarrhena asphodeloides (Zhimu) were obtained from Guangzhou LifeTech Pharmaceuticals Co. (Guangzhou, China). MSTF was the mixture of SA and TFA (the weight ratio of SA:TFA is 1:4). All other chemicals used were of analytical grade (Sinopharm Chemial Reagent Co., Shanghai, China).
4.2. Animals
SD (Sprague Dawley, SD) male rats (200 ± 20 g) were purchased from SLAC laboratory Animal Co., Ltd. (SLAC, Shanghai, China) and divided randomly into several groups. Animals were kept under standard laboratory conditions of temperature, pressure, humidity and a 12-h photoperiod. All animal procedures were performed according to the Guide for the Care and Use of Laboratory Animals recommended by the National Institutes of Health and approved by the Ethics Committee for Animal Experimentation of the Second Military Medical University.
4.3. Subcutaneous Injection with SM and Drug Administration
Rats were randomly assigned to one of the five following treatment groups (10 rats per group): normal control, SM and SM + MSTF low, medium and high dose (30, 60, 120 mg/kg body weight/day). For the subcutaneous injection, SM was dissolved in propylene glycol, and 0.1 mL were given using the gas-tight syringe on the back skin of the rats on a circular area of about 1.5 cm in diameter (the hair on the application site was closely clipped 24 h before SM application). The SM was applied as a single dose of 3.5 mg/kg body weight equivalent to 0.7 LD
50 (7-day observation for mortality) [
39,
40]. The equivalent volume of solvent was given to control rats. MSTF (120 mg/kg) dissolved in double-distilled water was administered intragastrically 1 h after the application of SM, and after, the same dose of MSTF was given every 24 h. For the survival study, different doses of MSTF were administered to the rats once each day for 7 consecutive days 1 h after SM treatment. We monitored animals three times daily for a period of 7 days to determine survival rates, and moribund animals were euthanized according to humane endpoints. The clinical criteria of moribund are being in the state of dying with no expectation of recovery, where animals display a combination of the following: lowered body temperature, continuous shaking, hunched back, impaired or slow motion and inability to maintain sternal recumbency [
41]. Moribund animals were placed in a separate cage with CO
2 until no breathing was observed, followed by a cervical dislocation as a secondary confirmatory method of euthanasia. Any surviving animals at the end of the study were also subjected to euthanasia by the application of CO
2 followed by cervical dislocation.
After 7 days, blood samples were obtained from periorbital sinus vein for the evaluation of oxidative markers, such as superoxide dismutase (SOD), reduced glutathione (GSH) and malondialdehyde (MDA).
4.4. Enzyme Activity Assay
Blood obtained from the periorbital sinus vein was collected and centrifuged at 3000 rpm for 10 min at 4 °C. The supernatant was used to perform the antioxidant enzyme assays using commercially available ELISA kits (Cayman Chemical Co., Ann Arbor, MI, USA). The plates were read at 450 nm with a correction wavelength of 550 nm on the BioTek Synergy 4 Multi-Mode Microplate Reader (BioTek Instruments, Inc., Winooski, VT, USA).
4.5. Sample Collection and Histopathologic Examination
Seventy-two hours following injection with SM, the rats were anesthetized and sacrificed by cervical dislocation. For testis, small intestine and liver histopathology analysis, the sections of tissue were processed for light microscopy. This processing consisted of specimen fixation in 4% buffered neutral formalin solution for 1–2 days, embedding in paraffin wax, slicing sections (5 μm) and staining them with hematoxylin and eosin (H&E). Histopathologic examination was blindly evaluated by a pathologist. In addition, 24 h following injection with SM, the rats were sacrificed. Liver tissues of all groups were processed for the microarray experiment.
4.6. Gene Expression Microarray and Data Analysis
The microarray experiments were performed as described previously [
42]. The 4 × 44 K Whole Rat Genome Oligo Microarray (Agilent Technologies, Santa Clara, CA, USA) was hybridized with Cy3-labeled cRNA using the Gene Expression Hybridization Kit (Agilent Technologies) in a hybridization oven (Agilent Technologies), according to the manufacturer’s instructions. Raw data were obtained by Feature Extraction software 10.7 (Agilent Technologies) and normalized by the Quantile algorithm, Gene Spring Software 11.0 (Agilent Technologies). The microarray experiments were conducted at the National Engineering Center for Biochip in Shanghai, China, according to the procedures in the Agilent technical manual. After normalization, genes in the treatment groups with at least a 1.5-fold change in expression were considered as upregulated or downregulated in comparison to non-treated groups (control). To determine significant proportions of differentially-expressed genes within treated groups, the hypergeometric probability
p was calculated.
p < 0.05 was considered significant.
Microarray data analysis was performed using the SBC Analysis system, which is available on the website:
http://sas.ebioservice.com/protal/root/molnet_shbh/index.jsp. The username and password to access this website are available upon request. A general description of the SBC analysis system can be found on the website:
http://www.ebioservice.com/eng/index.asp. The microarray data generated in this study have been deposited in the Gene Expression Omnibus (GEO) database under Accession Number GSE62994.
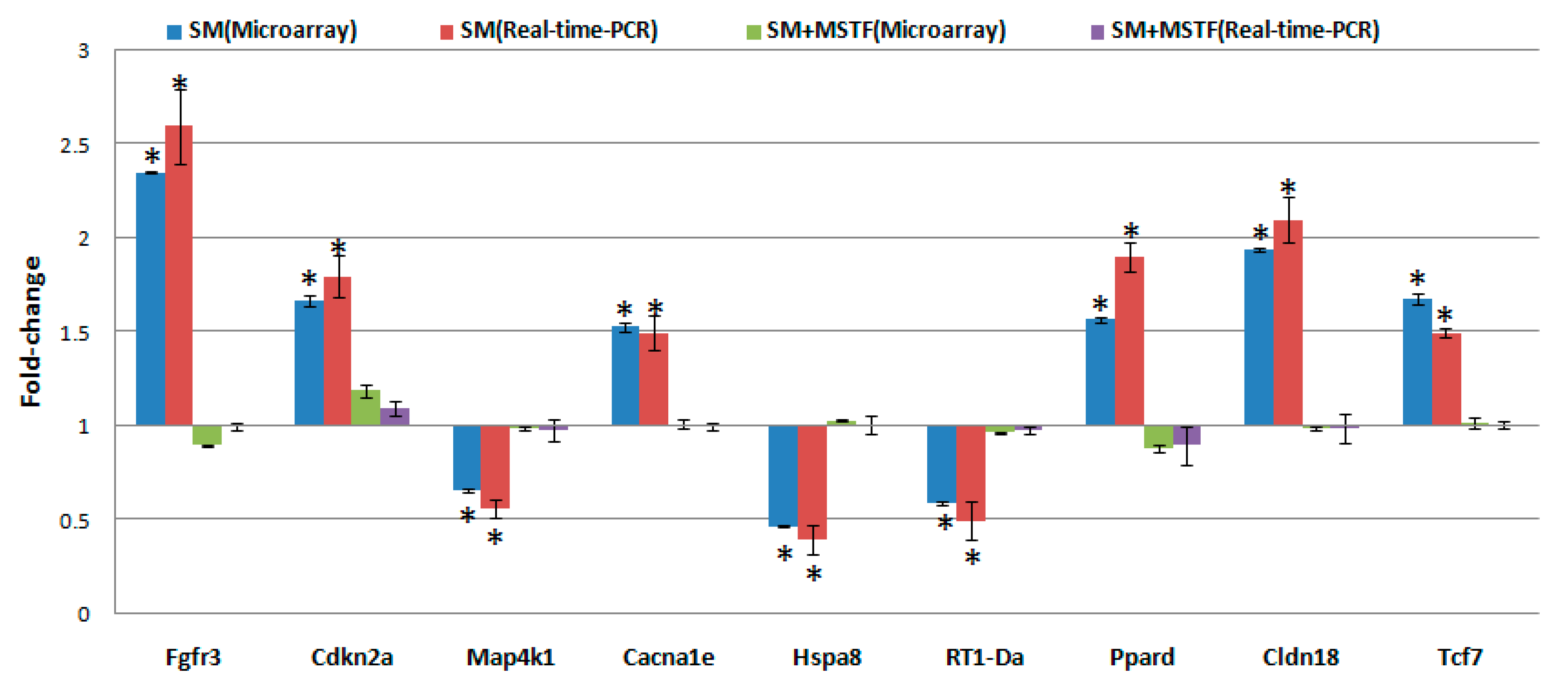
4.7. Quantitative Real-Time-PCR Array Validation
qRT-PCR was performed essentially as described previously [
42]. In total, nine genes were chosen for RT-PCR validation. PCR primers (
Table 7) were designed to either span or flank introns by using the ProbeFinder Version 2.50 (
http://www.roche-applied-science.com). Data are presented as the mean ± SD.
Table 7.
Oligonucleotides primers used in this study for quantitative real-time-PCR analysis. The primer sequences were designed by using the ProbeFinder Version 2.50 (
http://www.roche-applied-science.com).
Table 7.
Oligonucleotides primers used in this study for quantitative real-time-PCR analysis. The primer sequences were designed by using the ProbeFinder Version 2.50 (http://www.roche-applied-science.com).
| Accession No. | Symbol | Primer | Sequence |
|---|
| NM_053429 | Fgfr3 | Forward | acgccctacgtcactgtactc |
| Reverse | gaacctctagctccctgtcg |
| NM_031550 | Cdkn2a | Forward | cagattcgaactgcgagga |
| Reverse | tgcagtactaccagagtgtctagga |
| NM_001106243 | Map4k1 | Forward | cgcctgtctttcattggaac |
| Reverse | cacctttcagggccacag |
| NM_019294 | Cacna1e | Forward | taccgcgcctggatagac |
| Reverse | gctgatgttcccgagttttt |
| NM_024351 | Hspa8 | Forward | gcacaggaaaggagaacaagat |
| Reverse | catgcgctcaatatcctcct |
| NM_001008847 | RT1-Da | Forward | aacgcaacgcagtggaac |
| Reverse | tcaatgagctctcacggaag |
| NM_013141 | Ppard | Forward | ggaccagagcacacccttc |
| Reverse | gaggaaggggaggaattctg |
| NM_001014096 | Cldn18 | Forward | gtgccggccatacttcac |
| Reverse | atgcccacgatcatcagg |
| XM_001073458 | Tcf7 | Forward | tcccccatactgtgagctg |
| Reverse | ggcagggaagtgctgtctat |
| EF156276 | β-actin | Forward | cccgcgagtacaaccttct |
| Reverse | cgtcatccatggcgaact |
4.8. Statistical Analysis
Data were presented as the mean ± SD. One-way analysis of variance (ANOVA) followed by Dunnett’s test was used to examine differences among untreated (control) and treated (SM, SM + MSTF) groups. Differences were considered significant when p < 0.05.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}